
Effect of Salt Treatment on the Growth, Water Status, and Gas Exchange of Pyrus pyraster L. (Burgsd.) and Tilia cordata Mill. Seedlings


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
- Mn = moisture content (%) of the material n;
- Ww = wet weight of the sample;
- Wd = weight of the sample after drying.
2.3. Measurement and Analysis of Plant Parameters
2.4. Leaf Gas Exchange
2.5. Leaf Water Potential and Relative Water Content
2.6. Determination of Chlorophyll and Carotenoid Pigments
2.7. Ion Contents
2.8. Statistical Analysis
3. Results
3.1. Effect of Salinity on Growth and Mass Accumulation of Plant Organs
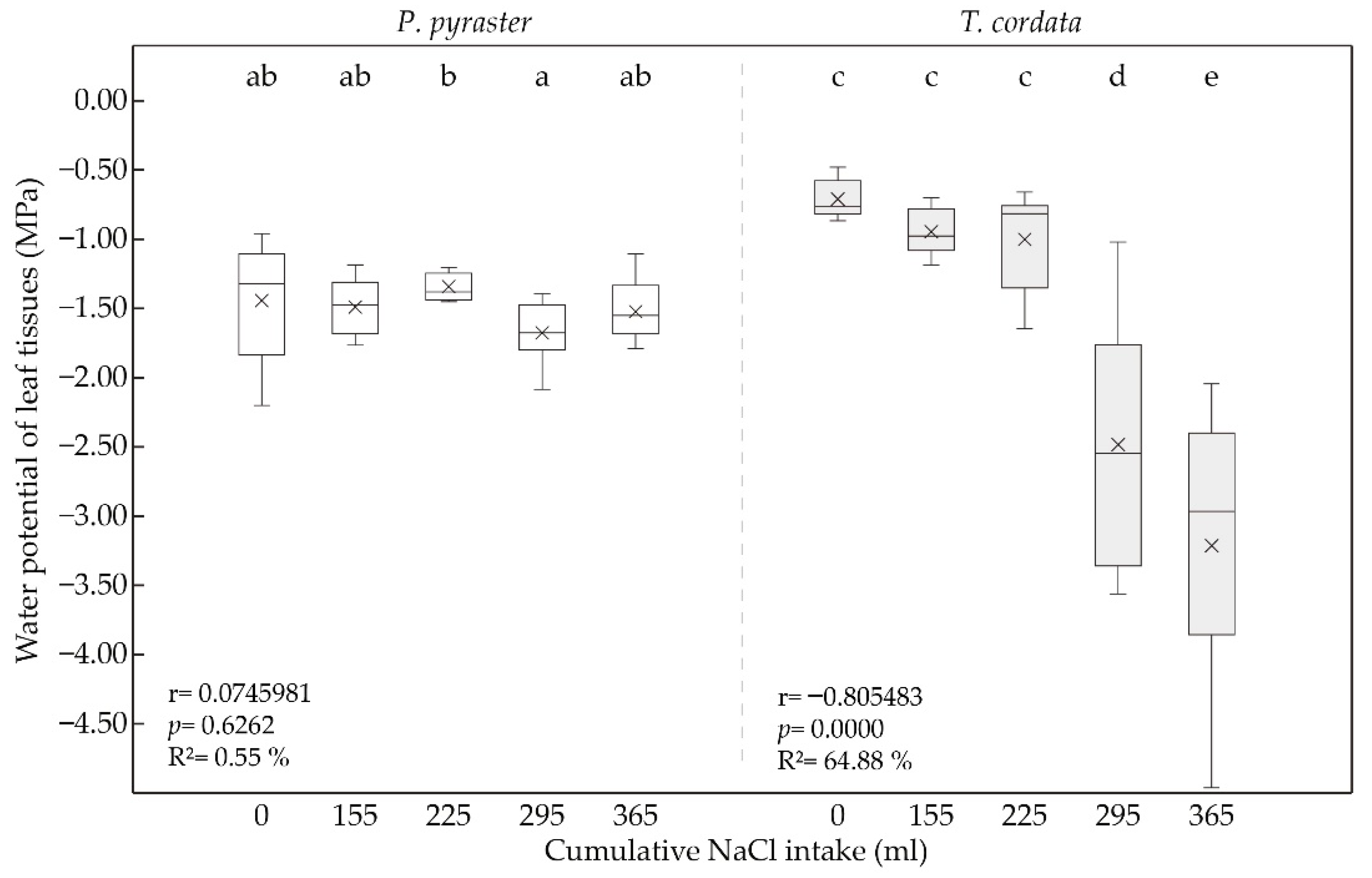
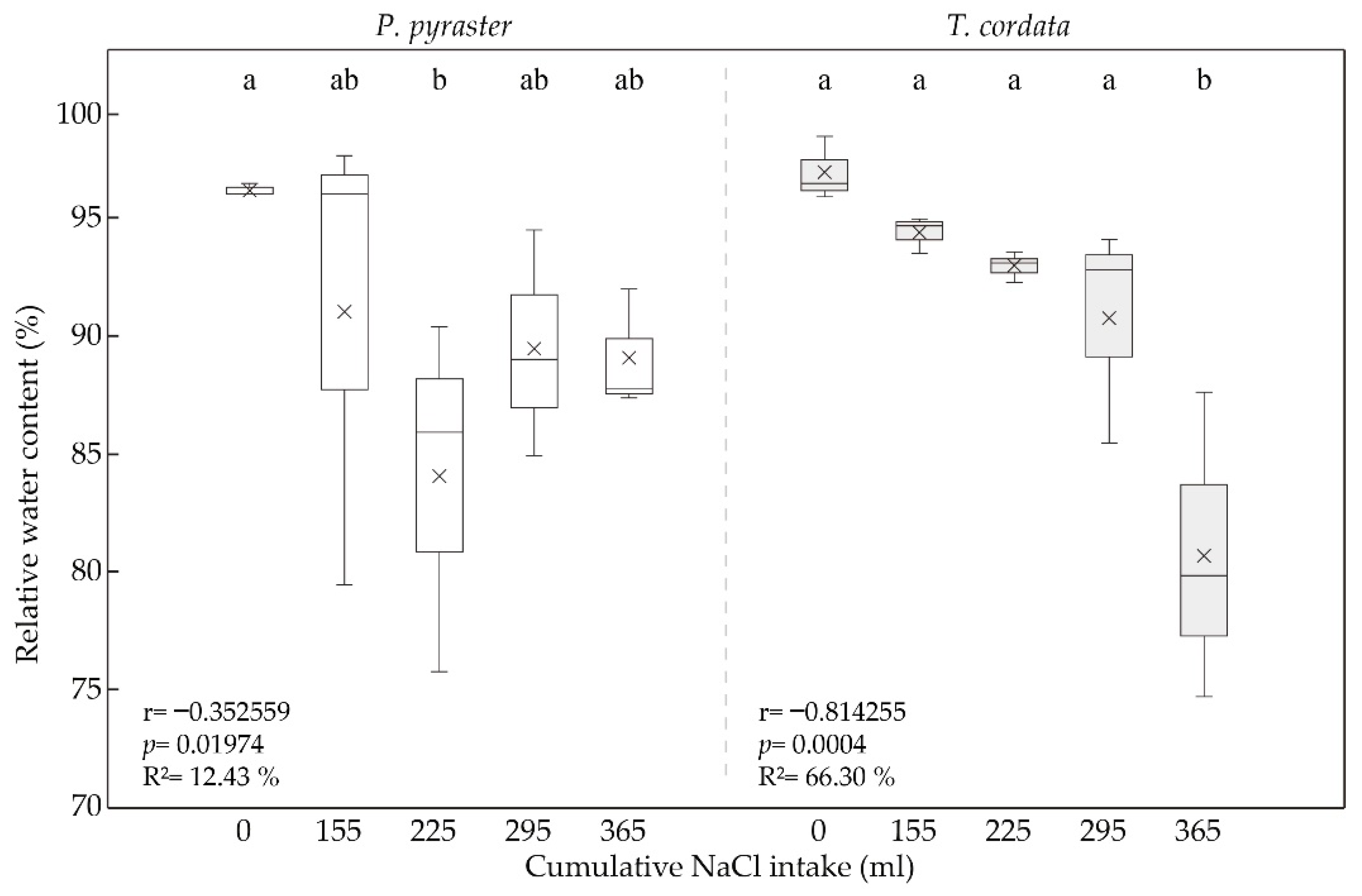
3.2. Leaf Water Status under Salt Treatment
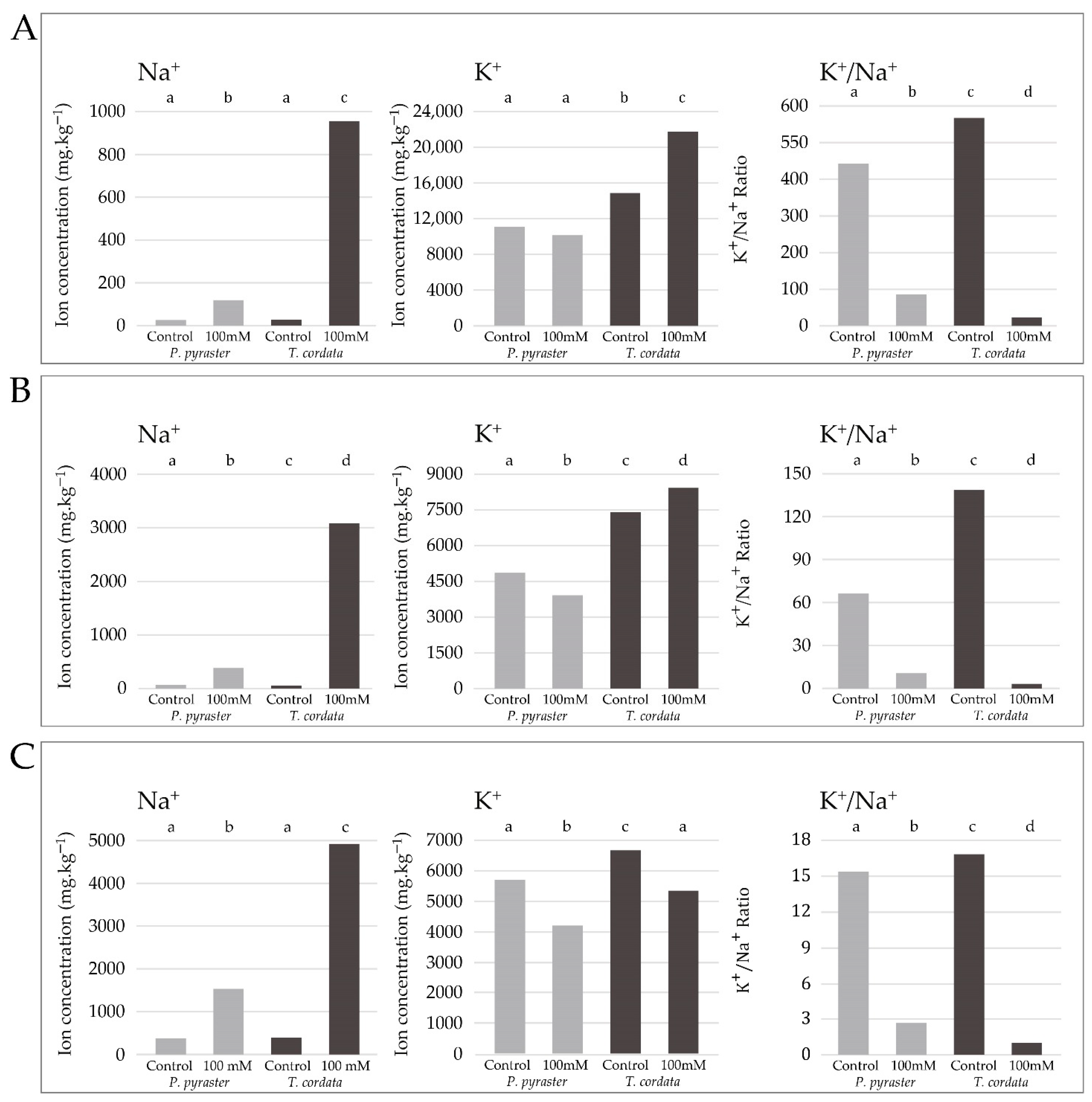
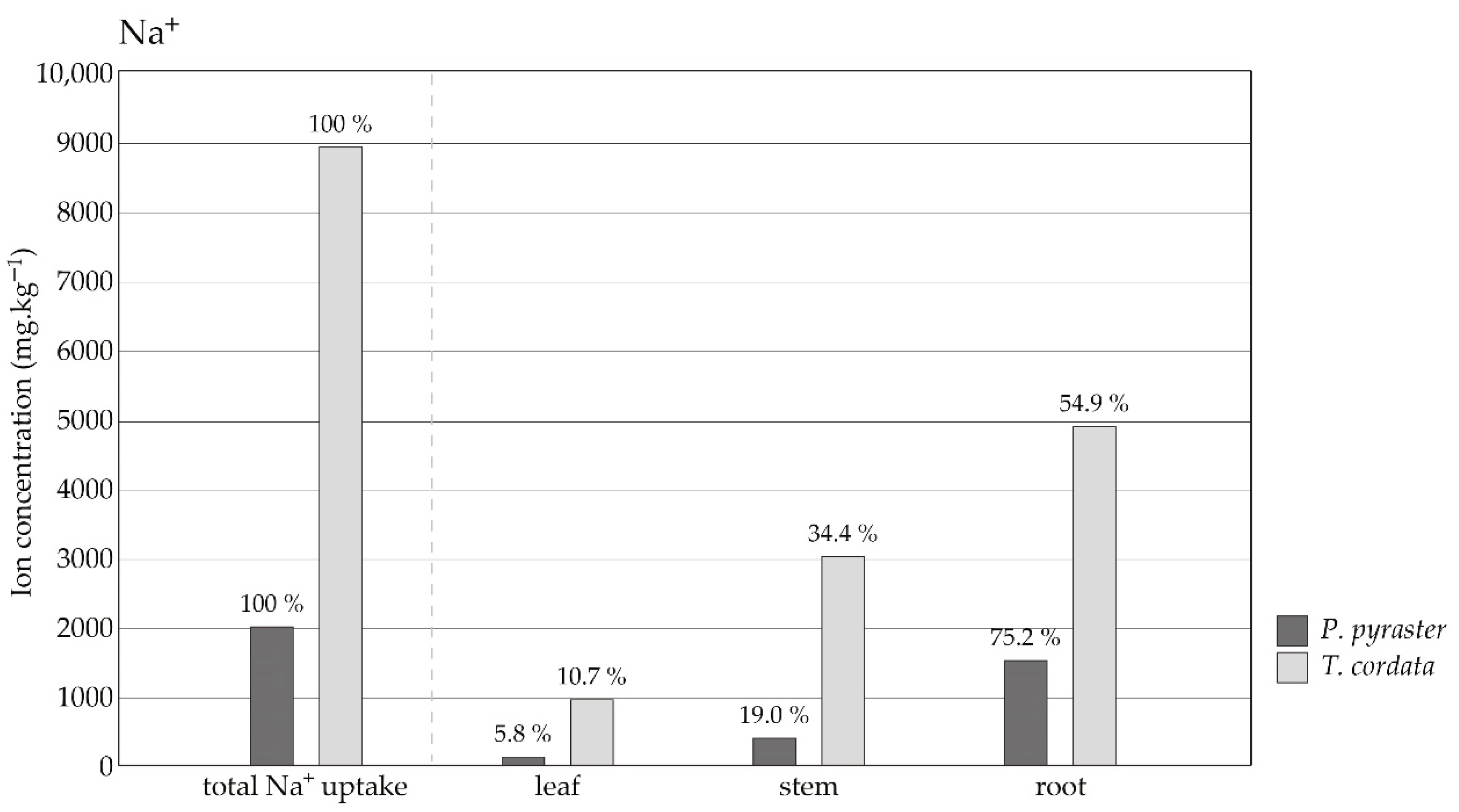
3.3. Salt Ion Uptake and Distribution
3.4. Effect of Salinity on Photosynthetic Pigments
3.5. Effect of Salinity on Leaf Gas Exchange
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Kordrostami, M.; Rabiei, B. Salinity Stress Tolerance in Plants: Physiological, Molecular, and Biotechnological Approaches. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Azza, M.A.M.; Fatma, A.M.; Farahat, M.M. Responses of ornamental plants and woody trees to salinity. World J. Agric. Sci. 2007, 3, 386–395. [Google Scholar]
- Qados, A.M.A. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba (L.). J. Saudi Soc. Agric. Sci. 2011, 10, 7–15. [Google Scholar]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Geissler, N.; Hussin, S.; Koyro, H.W. Interactive effects of NaCl salinity and elevated atmospheric CO2 concentration on growth, photosynthesis, water relations and chemical composition of the potential cash crop halophyte Aster tripolium L. Environ. Exp. Bot. 2009, 65, 220–231. [Google Scholar] [CrossRef]
- Zhang, Z.-H.; Liu, Q.; Song, H.-X.; Rong, X.-M.; Ismail, A.M. Responses of different rice (Oryza sativa L.) genotypes to salt stress and relation to carbohydrate metabolism and chlorophyll content. Afr. J. Agric. Res. 2012, 7, 19–27. [Google Scholar]
- Pauleit, S.; Jones, N.; Garcia-Martin, G.; Garcia-Valdecantos, J.L.; Rivière, L.M.; Vidal-Beaudet, L.; Bodson, M.; Randrup, T.B. Tree establishment practice in towns and cities–Results from a European survey. Urban For. Urban Green. 2002, 1, 83–96. [Google Scholar] [CrossRef]
- Pigott, C.D. Tilia cordata Miller. (T. europaea L. pro parte, T. parvifolia Ehrh. ex Hoffm., T. sylvestris Desf., T. foemina folio minore Bauhin). J. Ecol. 1991, 79, 1147–1207. [Google Scholar] [CrossRef]
- Rameau, J.C.; Mansion, D.; Dumé, G.; Timbal, J.; Lecointe, A.; Dupont, P.; Keller, R. Flore Forestière Française. Plaines et Collines; IDF: Paris, France; ENGREF: Nancy, France, 1989. [Google Scholar]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate Northern Hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Pigott, D. Lime-Trees and Basswoods: A Biological Monograph of the Genus Tilia, 1st ed.; Cambridge University Press: New York, NY, USA, 2012; p. 405. [Google Scholar]
- De Jaegere, T.D.; Hein, S.; Claessens, H. A Review of the Characteristics of Small-Leaved Lime (Tilia cordata Mill.) and Their Implications for Silviculture in a Changing Climate. Forests 2016, 7, 56. [Google Scholar] [CrossRef]
- Moser, A.; Rötzer, T.; Pauleit, S.; Pretzsch, H. Structure and ecosystem services of small-leaved lime (Tilia cordata Mill.) and black locust (Robinia pseudoacacia L.) in urban environments. Urban For. Urban Green. 2015, 14, 1110–1121. [Google Scholar] [CrossRef]
- Jensen, J.S. Lime (Tilia cordata and Tilia platyphyllos). In EUFORGEN Technical Guidelines for Genetic Conservation and Use for Lime (Tilia spp.); International Plant Genetic Resources Institute: Rome, Italy, 2003. [Google Scholar]
- Martynova, M.; Sultanova, R.; Odintsov, G.; Sazgutdinova, R.; Khanova, E. Growth of Tilia cordata Mill. in urban forests. South East Eur. For. SEEFOR 2020, 11, 51–59. [Google Scholar] [CrossRef]
- Stephan, B.R.; Wagner, I.; Kleinschmit, J. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Wild Apple and Pear (Malus sylvestris and Pyrus pyraster); International Plant Genetic Resources Institute: Rome, Italy, 2003. [Google Scholar]
- Wagner, I.; Büttner, R. Hybridization in wild pear (Pyrus pyraster) from various regions in Germany and from Luxembourg with respect to Pyrus × communis. In Proceedings of the III International Symposium on Horticulture in Europe-SHE2016, Chania, Greece, 17–21 October 2016; pp. 427–434. [Google Scholar]
- Ellenberg, H. Vegetation Mitteleuropas mit den Alpen in Ökologischer Sicht, 3rd ed.; Ulmer Verlag: Stuttgart, Germany, 1983. [Google Scholar]
- Paganová, V. Wild pear Pyrus pyraster (L.) Burgsd. requirements on environmental conditions. Ekológia 2003, 22, 225–241. [Google Scholar]
- Milner, E. Trees of Britain and Ireland; Natural History Museum: London, UK, 2011. [Google Scholar]
- Lawesson, J.E.; Oksanen, J. Niche characteristics of Danish woody species as derived from coenoclines. J. Veg. Sci. 2002, 13, 279–290. [Google Scholar] [CrossRef]
- Shannon, M.C.; Grieve, C.M.; Francois, L.E. Whole-plant response to salinity. In Plant—Environment Interactions; Wilkinson, R.E., Ed.; Marcel Dekker: New York, NY, USA, 1994; pp. 199–244. [Google Scholar]
- Kozlowski, T.T. Responses of woody plants to flooding and salinity. Tree Physiol. 1997, 17, 490. [Google Scholar] [CrossRef]
- Lapin, M.; Faško, P.; Melo, M.; Štastný, P.; Tomlain, J. Klimatické oblasti. In Atlas Krajiny Slovenskej Republiky; Ministerstvo životného prostredia: Banská Bystrica, Slovakia, 2002; p. 344. [Google Scholar]
- Trautmann, N.; Richard, T. Moisture Content. Cornell Waste Management Institute. 1996. Available online: http://compost.css.cornell.edu/calc/moisture_content.html (accessed on 15 August 2021).
- Ostonen, I.; Püttstep, U.; Biel, C.; Alberton, O.; Bakker, M.R.; Löhmus, H.; Majdi, D.; Metcalfe, D.; Olsthoorn, A.F.M.; Pronk, A.; et al. Specific root length as an indicator of environmental changes. Plant Biosyst. 2007, 141, 3426–3442. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Parsons, R.; Weyers, J.D.B.; Lawson, T.; Godber, I.M. Rapid and straightforward estimates of photosynthetic characteristics using a portable gas exchange system. Photosynthetica 1998, 34, 265–279. [Google Scholar] [CrossRef]
- Hunt, S. Measurements of photosynthesis and respiration in plants. Physiol. Plant. 2003, 117, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimating water deficit in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Extraction of photosynthetic tissues: Chlorophylls and carotenoids. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.2.1–F4.2.6. [Google Scholar] [CrossRef]
- Kovacik, A.; Tvrda, E.; Miskeje, M.; Arvay, J.; Tomka, M.; Zbynovska, K. Trace metals in the freshwaterfish Cyprinus carpio: Effect to serum biochemistry and oxidative status markers. Biol. Trace Elem. Res. 2018, 188, 494–507. [Google Scholar] [CrossRef]
- Ziska, L.H.; Seeman, L.H.; DeJong, T.M. Salinity induced limitations on photosynthesis in Prunus salicina, a deciduous tree species. Plant Physiol. 1990, 93, 864–870. [Google Scholar] [CrossRef]
- Zekri, M.; Parsons, L.R. Response of split-root sour orange seedlings to NaCl and polyethylene glycol stresses. J. Exp. Bot. 1990, 41, 35–40. [Google Scholar] [CrossRef]
- Abassi, M.; Mguis, K.; Béjaoui, Z.; Albouchi, A. Morphogenetic responses of Populus alba L. under salt stress. J. For. Res. 2014, 25, 155–161. [Google Scholar] [CrossRef]
- Chen, P.F.; Zuo, L.H.; Yu, X.Y.; Dong, Y.; Zhang, S.; Yang, M. Response mechanism in Populus× euramericana cv. ‘74/76′ revealed by RNA-seq under salt stress. Acta Physiol. Plant 2018, 40, 96. [Google Scholar] [CrossRef]
- Paganová, V.; Jureková, Z.; Lichtnerová, H. The nature and way of root adaptation of juvenile woody plants Sorbus and Pyrus to drought. Environ. Monit. Assess. 2019, 191, 714. [Google Scholar] [CrossRef]
- Paganová, V.; Hus, M.; Jureková, Z. Physiological performance of Pyrus pyraster L. (Burgsd.) and Sorbus torminalis (L.) crantz seedlings under drought treatment. Plants 2020, 9, 1496. [Google Scholar] [CrossRef]
- Hsiao, T.C.; Xu, L.K. Sensitivity of growth of roots versus leaves to water stress: Biophysical analysis and relation to water transport. J. Exp. Bot. 2000, 51, 1595–1616. [Google Scholar] [CrossRef]
- Cassaniti, C.; Leonardi, C.; Flowers, T.J. The effects of sodium chloride ornamental shrubs. Sci. Hortic. 2009, 122, 586–593. [Google Scholar] [CrossRef]
- Cassaniti, C.; Romano, D.; Flowers, T.J. The response of ornamental plants to saline irrigation water. In Irrigation Water Management. Pollution and Alternative Strategies; Garcia-Garizabal, I., Ed.; InTech Europe: Rijeka, Croatia, 2012; pp. 132–158. [Google Scholar]
- Boursier, P.; Läuchli, A. Growth responses and mineral nutrient relations of salt stressed sorghum. Crop Sci. 1990, 30, 1226–1233. [Google Scholar] [CrossRef]
- Maathuis, F.J.M.; Amtmann, A. K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ ratios. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef]
- Pérez-Alfocea, F.; Balibrea, M.E.; Alarçon, J.J.; Bolarín, M.C. Composition of xylemand phloem exudates in relation to the salt tolerance of domestic and wild tomato species. J. Plant Physiol. 2000, 156, 367–374. [Google Scholar] [CrossRef]
- Colmer, T.D.; Munns, R.; Flowers, T.J. Improving salt tolerance of wheat and barley: Future prospects. Aust. J. Exp. Agric. 2005, 45, 1425–1443. [Google Scholar] [CrossRef]
- Murillo-Amador, B.; Troyo-Diéguez, E.; García-Hernández, J.L.; López-Aguilar, R.; Ávila-Serrano, N.Y.; Zamora-Salgado, S.; Rueda-Puente, E.O.; Kaya, C. Effect of NaCl salinity in the genotypic variation of cowpea (Vigna unguiculata) during early vegetative growth. Sci. Hortic. 2006, 108, 423–441. [Google Scholar] [CrossRef]
- Matsumoto, K.; Tamura, F.; Chun, J.; Tanabe, K. Native Mediterranean Pyrus rootstock, P. amygdaliformis and P. elaeagrifolia present higher tolerance to salinity stress compared with Asian natives. J. Jpn. Soc. Hort. Sci. 2006, 75, 450–457. [Google Scholar] [CrossRef][Green Version]
- Boland, A.M.; Jerie, P.; Maas, E. Long-term effects of salinity on fruit trees. Acta Hortic. 1997, 449, 599–606. [Google Scholar] [CrossRef]
- Munns, R.; Termaat, A. Whole-plant responses to salinity. Funct. Plant Biol. 1986, 13, 143–160. [Google Scholar] [CrossRef]
- Laffray, X.; Alaoui-Sehmer, L.; Bourioug, M.; Bourgeade, P.; Alaoui-Sossé, B.; Aleya, L. Effects of sodium chloride salinity on ecophysiological and biochemical parameters of oak seedlings (Quercus robur L.) from use of de-icing salts for winter road maintenance. Environ. Monit Assess. 2018, 190, 266. [Google Scholar] [CrossRef] [PubMed]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Tester, M.; Davenport, R. Na+ Tolerance and Na+ Transport in Higher Plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, L.; Chen, B.; Qin, Z.; Xiao, Y.; Zhang, Y.; Yao, R.; Liu, H.; Yang, H. Progress in Understanding the Physiological and Molecular Responses of Populus to Salt Stress. Int. J. Mol. Sci. 2019, 20, 1312. [Google Scholar] [CrossRef]
- Navarro, A.; Bañón, S.; Conejero, W.; Sánchez-Blanco, M.J. Ornamental characters. ion accumulation and water status in Arbutus unedo seedlings irrigated with saline water and subsequent relief and transplanting. Environ. Exp. Bot. 2008, 62, 364–370. [Google Scholar] [CrossRef]
- Franco, J.A.; Bañón, S.; Vicente, M.J.; Miralles, J.; Martínez-Sánchez, J.J. Root development in horticultural plants grown under abiotic stress conditions—A review. J. Hortic. Sci. Biotechnol. 2011, 86, 543–556. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.J.; Rodríguez, P.; Olmos, E.; Morales, M.A.; Torrecillas, A. Differences in the effects of Simulated Sea Aerosol on Water Relations, Salt Content, and Leaf Ultrastructure of Rock-Rose Plants. J. Environ. Qual. 2004, 33, 1369–1375. [Google Scholar] [CrossRef]
- Slama, I.; Ghnaya, T.; Savouré, A.; Abdelly, C. Combined effects of long-term salinity and soil drying on growth, water relations, nutrient status and proline accumulation of Sesuvium portulacastrum. C. R. Biologies 2008, 331, 442–451. [Google Scholar] [CrossRef]
- Álvarez, S.; Gómez-Bellot, M.J.; Castillo, M.; Bañón, S.; Sánchez-Blanco, M.J. Osmotic and saline effect on growth water relations and ion uptake and translocation in Phlomis purpurea plants. Environ. Exp. Bot. 2012, 78, 138–145. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.J.; Rodríguez, P.; Morales, M.A.; Ortuño, M.F.; Torrecillas, A. Comparative growth and water relations of Cistus albidus and Cistus monspeliensis plants during water deficit conditions and recovery. Plant Sci. 2002, 162, 107–113. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of Salt Tolerance in Non-Halophytes. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Stepien, P.; Johnson, G.N. Contrasting responses of photosynthesis to salt stress in the glycophyte Arabidopsis and the halophyte thellungiella: Role of the plastid terminal oxidase as an alternative electron sink. Plant Physiol. 2009, 149, 1154–1165. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W. The role of xanthophylls cycle carotenoids in the protection of photosynthesis. Trends. Plant Sci. 1996, 1, 21–26. [Google Scholar] [CrossRef]
- Davies, W.J.; Kudoyarova, G.; Hartung, W. Long-distance ABA signaling and its relation to other signaling pathways in the detection of soil drying and the mediation of the plant’s response to drought. J. Plant Growth Regul. 2005, 24, 285–295. [Google Scholar] [CrossRef]
- James, R.A.; Rivelli, A.R.; Munns, R.; Caemmerer, S.V. Factors affecting CO2 assimilation.leaf injury and growth in salt-stressed durum wheat. Funct. Plant Biol. 2002, 29, 1393–1403. [Google Scholar] [CrossRef]
- Chaves, M.M.; Osorio, J.; Pereira, J.S. Water use efficiency and photosynthesis. In Water Use Efficiency in Plant Biology; Bacon, M., Ed.; Blackwell Publishing: Oxford, UK, 2004; pp. 42–74. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Location | Exposure | Altitude (m) | TI.(°C) | TVII. (°C) | Precipitation (mm) | Type |
|---|---|---|---|---|---|---|---|
| P. pyraster | Kremnica hills (Tŕnie) | S | 540 | −3 | 18 | 750 | MW |
| T. cordata | Dubeň (Žilina) | S | 410 | −3.5 | 16 | 800 | MW |
| p-Value | Control | 100 mmol NaCl | |||||
|---|---|---|---|---|---|---|---|
| Parameter | T | S | T × S | P. pyraster | T. cordata | P. pyraster | T. cordata |
| Stem length (mm) | 0.00 | 0.02 | 0.84 | 281.94 (±64.82) b | 399.90 (±91.42) a | 242.79 (±49.69) b | 353.50 (±74.51) a |
| Stem increment (mm) | 0.00 | 0.02 | 0.42 | 117.44 (±62.65) b | 235.70 (±82.90) a | 65.86 (±37.06) c | 210.40 (±71.10) a |
| LA (mm2) | 0.00 | 0.00 | 0.02 | 12,843.20 (±4810.70) b | 51,364.80 (±10,898.80) a | 8340.84 (±2732.16) c | 37,860.00 (±9278.79) d |
| SLA (mm2·mg−1) | 0.00 | 0.00 | 0.18 | 17.76 (±2.23) b | 33.01 (±5.06) a | 15.53 (±2.22) c | 28.19 (±5.04) d |
| DWL (mg) | 0.00 | 0.01 | 0.74 | 723.78 (±256.67) b | 1594.65 (±423.24) a | 544.57 (±183.86) c | 1364.20 (±332.04) a |
| SDW (mg) | 0.00 | 0.00 | 0.01 | 1134.83 (±469.02) b | 2357.10 (±779.90) a | 979.21 (±369.60) b | 1542.45 (±397.21) c |
| DWS (mg) | 0.00 | 0.00 | 0.08 | 1858.61 (±710.76) b | 3951.75 (±1185.07) a | 1523.79 (±505.60) b | 2906.65 (±709.21) c |
| RBI (%) | 0.01 | 0.00 | 0.36 | 123.80 (±67.41) b | 175.13 (±70.47) a | 54.87 (±37.68) c | 80.49 (±45.98) c |
| FWS (mg) | 0.00 | 0.00 | 0.00 | 4218.50 (±1628.89) a | 11,942.20 (±3353.35) b | 3201.21 (±1059.03) a | 7386.70 (±1866.87) c |
| FWR (mg) | 0.00 | 0.00 | 0.00 | 3588.11 (±1241.37) a | 8170.40 (±2945.84) c | 3083.79 (±1127.77) a | 4164.90 (±885.26) a |
| RL (mm) | 0.00 | 0.00 | 0.00 | 7826.26 (±1925.41) a | 19,607.50 (±4212.76) b | 8439.39 (±3935.52) ac | 10,326.20 (±1812.39) c |
| SRL (mm·mg−1) | 0.00 | 0.91 | 0.30 | 5.81 (±1.58) b | 8.10 (±2.54) a | 6.45 (±3.10) ab | 7.59 (±2.05) a |
| DWR (mg) | 0.00 | 0.00 | 0.00 | 1405.33 (±420.96) b | 2650.30 (±928.65) a | 1377.79 (±511.18) b | 1429.55 (±331.48) b |
| RSA (mm2) | 0.00 | 0.00 | 0.00 | 11,224.10 (±2955.99) b | 25,545.90 (±6564.05) a | 8812.52 (±3580.33) c | 13,677.80 (±2451.54) d |
| ARD (mm) | 0.13 | 0.00 | 0.00 | 0.54 (±0.06) a | 0.49 (±0.06) b | 0.40 (±0.06) c | 0.49 (±0.04) b |
| NRT | 0.00 | 0.00 | 0.00 | 2044.78 (±710.42) b | 7375.80 (±2502.22) a | 2742.93 (±1178.07) c | 3923.70 (±976.10) d |
| RV (mm3) | 0.00 | 0.00 | 0.00 | 4981.76 (±1804.35) b | 12,760.70 (±4552.20) a | 3386.21 (±1260.69) c | 5919.14 (±1548.73) b |
| R:S | 0.00 | 0.93 | 0.00 | 0.83 (±0.22) a | 0.68 (±0.16) b | 1.00 (±0.42) a | 0.50 (±0.09) c |
| p-Value | Control | 100 mM NaCl | |||||
|---|---|---|---|---|---|---|---|
| Parameter | T | S | T × S | P. pyraster | T. cordata | P. pyraster | T. cordata |
| Chl a (mg·mm−2) | 0.00 | 0.04 | 0.02 | 378.15 (±78.82) a | 289.94 (±41.93) b | 380.93 (±87.60) a | 221.22 (±45.92) c |
| Chl b (mg·mm−2) | 0.00 | 0.02 | 0.05 | 145.29 (±34.35) a | 122.54 (±16.31) b | 143.13 (±32.20) a | 95.52 (±19.79) c |
| Chl ab (mg·mm−2) | 0.00 | 0.03 | 0.03 | 523.31 (±112.26) a | 412.37 (±57.72) b | 523.92 (±117.65) a | 316.67 (±65.38) c |
| Chl a/b (mg·mm−2) | 0.00 | 0.96 | 0.25 | 2.62 (±0.14) a | 2.37 (±0.10) b | 2.66 (±0.29) a | 2.32 (±0.11) b |
| Carotenoids (mg·mm−2) | 0.00 | 0.62 | 0.22 | 109.79 (±19.55) a | 83.78 (±11.82) b | 112.87 (±23.30) a | 76.63 (±14.00) b |
| CSI (mL Plant−1) DOT | Parameter | p-Value | P. pyraster | T. cordata | ||||
|---|---|---|---|---|---|---|---|---|
| T | S | T × S | Control | 100 mM NaCl | Control | 100 mM NaCl | ||
| 155 | gs (mmol H2O m−2 s−1) | 0.06 | 0.00 | 0.53 | 83.00 (±19.65) a | 46.80 (±14.60) bc | 69.50 (±4.95) ac | 21.50 (±0.71) b |
| 20th | An (μmol CO2 m−2 s−1) | 0.85 | 0.34 | 0.13 | 4.66 (±1.17) a | 4.36 (±0.57) a | 4.00 (±0.44) a | 5.20 (±0.42) a |
| E (mmol H2O m−2 s−1) | 0.22 | 0.01 | 0.48 | 0.73 (±0.23) a | 0.44 (±0.13) b | 0.67 (±0.24) ab | 0.23 (±0.01) b | |
| WUE (mmol CO2 mol−1 H2O) | 0.00 | 0.00 | 0.00 | 7.11 (±3.15) a | 8.61 (±1.23) a | 6.61 (±2.55) a | 22.60 (±0.46) b | |
| 225 | gs (mmol H2O m−2 s−1) | 0.00 | 0.00 | 0.38 | 144.60 (±24.11) a | 96.40 (±22.50) b | 59.25 (±15.35) c | 28.00 (±11.63) d |
| 30th | An (μmol CO2 m−2 s−1) | 0.00 | 0.13 | 0.61 | 7.44 (±0.80) a | 6.98 (±0.39) a | 5.46 (±1.02) b | 4.56 (±1.31) b |
| E (mmol H2O m−2 s−1) | 0.00 | 0.02 | 0.51 | 1.16 (±0.15) a | 1.02 (±0.15) a | 0.65 (±0.18) b | 0.40 (±0.17) b | |
| WUE (mmol CO2 mol−1 H2O) | 0.00 | 0.21 | 0.47 | 6.48 (±1.00) a | 6.96 (±0.87) a | 10.35 (±2.60) b | 12.07 (±2.36) b | |
| 295 | gs (mmol H2O m−2 s−1) | 0.00 | 0.00 | 0.90 | 103.2 (±29.12) a | 69.25 (±27.11) ab | 61.89 (±23.18) b | 25.40 (±14.72) c |
| 40th | An (μmol CO2 m−2 s−1) | 0.00 | 0.64 | 0.36 | 7.62 (±0.70) a | 7.06 (±1.18) a | 5.71 (±1.03) b | 4.13 (±2.05) b |
| E (mmol H2O m−2 s−1) | 0.01 | 0.00 | 0.37 | 0.74 (±0.16) a | 0.56 (±0.20) a | 0.58 (±0.19) a | 0.27 (±0.12) b | |
| WUE (mmol CO2 mol−1 H2O) | 0.74 | 0.03 | 0.63 | 10.79 (±2.59) ab | 13.35 (±4.48) ab | 10.55 (±3.02) a | 14.51 (±3.83) b | |
| 365 | gs (mmol H2O m−2 s−1) | 0.00 | 0.21 | 0.80 | 115.00 (±30.22) a | 101.80 (±39.06) a | 56.57 (±25.05) b | 37.00 (±8.79) b |
| 50th | An (μmol CO2 m−2 s−1) | 0.00 | 0.06 | 0.42 | 7.36 (±0.82) a | 6.86 (±1.21) a | 5.11 (±0.74) b | 3.95 (±0.85) c |
| E (mmol H2O m−2 s−1) | 0.00 | 0.27 | 0.93 | 0.87 (±0.17) a | 0.79 (±0.22) ac | 0.53 (±0.19) bc | 0.43 (±0.13) b | |
| WUE (mmol CO2 mol−1 H2O) | 0.35 | 0.75 | 0.96 | 8.47 (±1.62) a | 8.91 (±0.91) a | 9.66 (±3.05) a | 9.98 (±3.47) a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paganová, V.; Hus, M.; Lichtnerová, H. Effect of Salt Treatment on the Growth, Water Status, and Gas Exchange of Pyrus pyraster L. (Burgsd.) and Tilia cordata Mill. Seedlings. Horticulturae 2022, 8, 519. https://doi.org/10.3390/horticulturae8060519
Paganová V, Hus M, Lichtnerová H. Effect of Salt Treatment on the Growth, Water Status, and Gas Exchange of Pyrus pyraster L. (Burgsd.) and Tilia cordata Mill. Seedlings. Horticulturae. 2022; 8(6):519. https://doi.org/10.3390/horticulturae8060519
Chicago/Turabian StylePaganová, Viera, Marek Hus, and Helena Lichtnerová. 2022. "Effect of Salt Treatment on the Growth, Water Status, and Gas Exchange of Pyrus pyraster L. (Burgsd.) and Tilia cordata Mill. Seedlings" Horticulturae 8, no. 6: 519. https://doi.org/10.3390/horticulturae8060519
APA StylePaganová, V., Hus, M., & Lichtnerová, H. (2022). Effect of Salt Treatment on the Growth, Water Status, and Gas Exchange of Pyrus pyraster L. (Burgsd.) and Tilia cordata Mill. Seedlings. Horticulturae, 8(6), 519. https://doi.org/10.3390/horticulturae8060519