
Involvement of Organic Acid in the Control Mechanism of ε-Poly-L-lysine (ε-PL) on Blue Mold Caused by Penicillium expansum in Apple Fruits

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Culture and Preparation of Spore Suspension
2.2. Fruit
2.3. Effect of ε-PL Treatment on the Growth of P. expansum
2.4. Effect of ε-PL Treatment on Environmental pH Regulation by P. expansum In Vitro
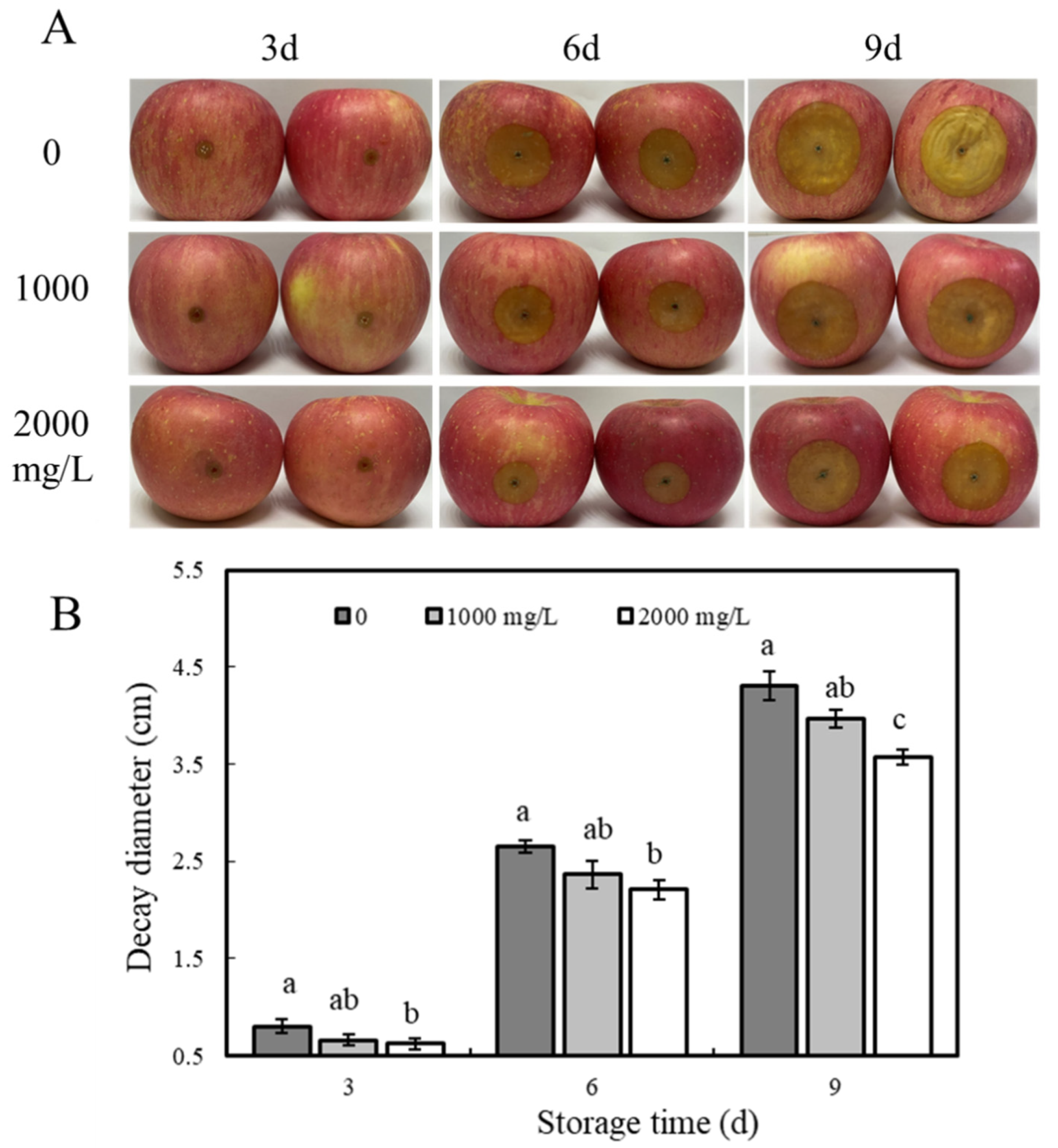
2.5. Effect of ε-PL Treatment on Blue Mold Disease in Apple Fruits
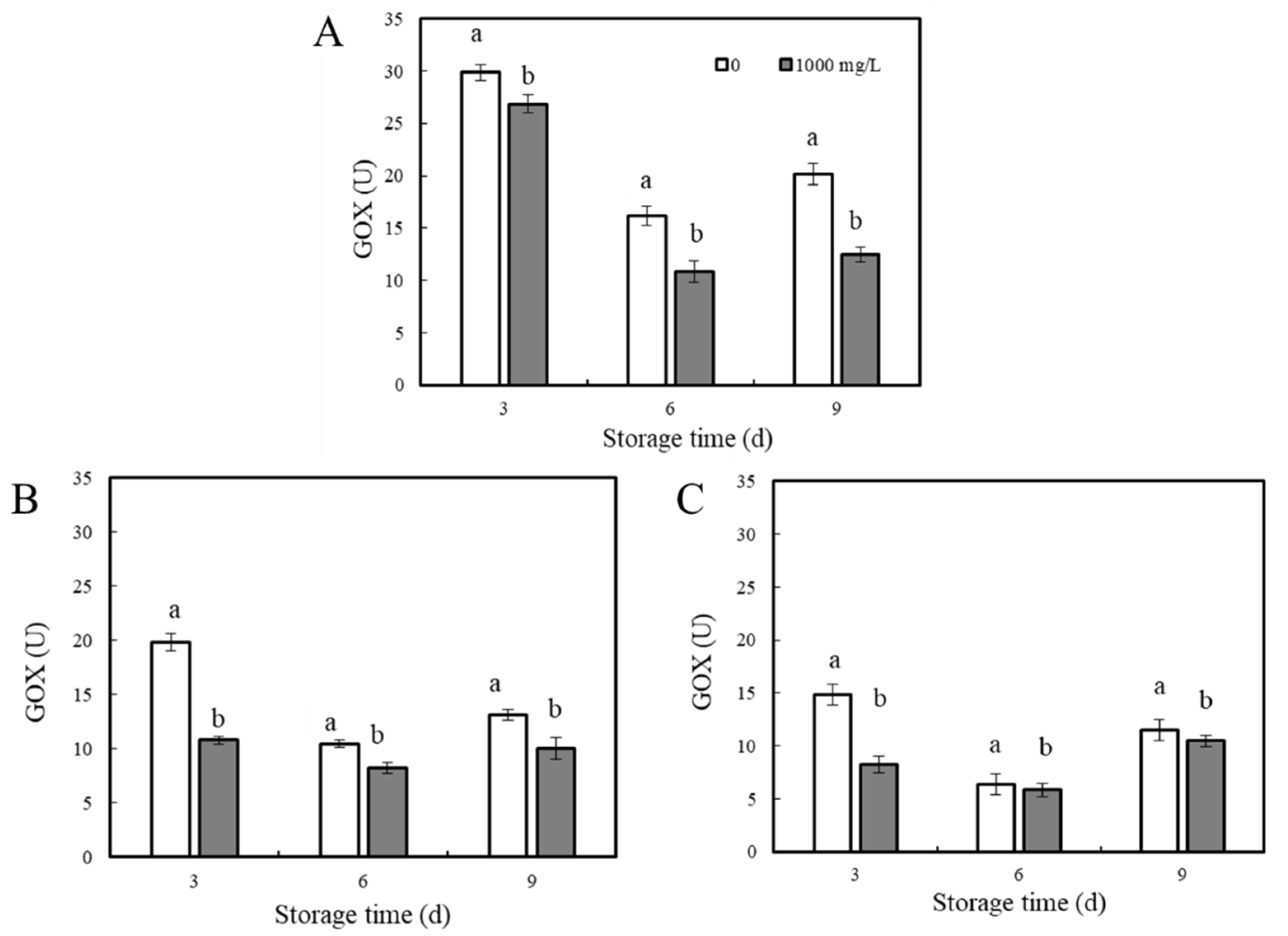
2.6. Determination of GOX Activity
2.7. Determination of Organic Acids
2.8. Determination of Cell-Wall-Degrading Enzyme Activity
2.9. Data Analysis
3. Result
3.1. Effect of ε-PL Treatment on the Growth of P. expansum
3.1.1. In Vitro Test
3.1.2. In Vivo Test
3.2. Effect of ε-PL Treatment on Environmental pH of P. expansum
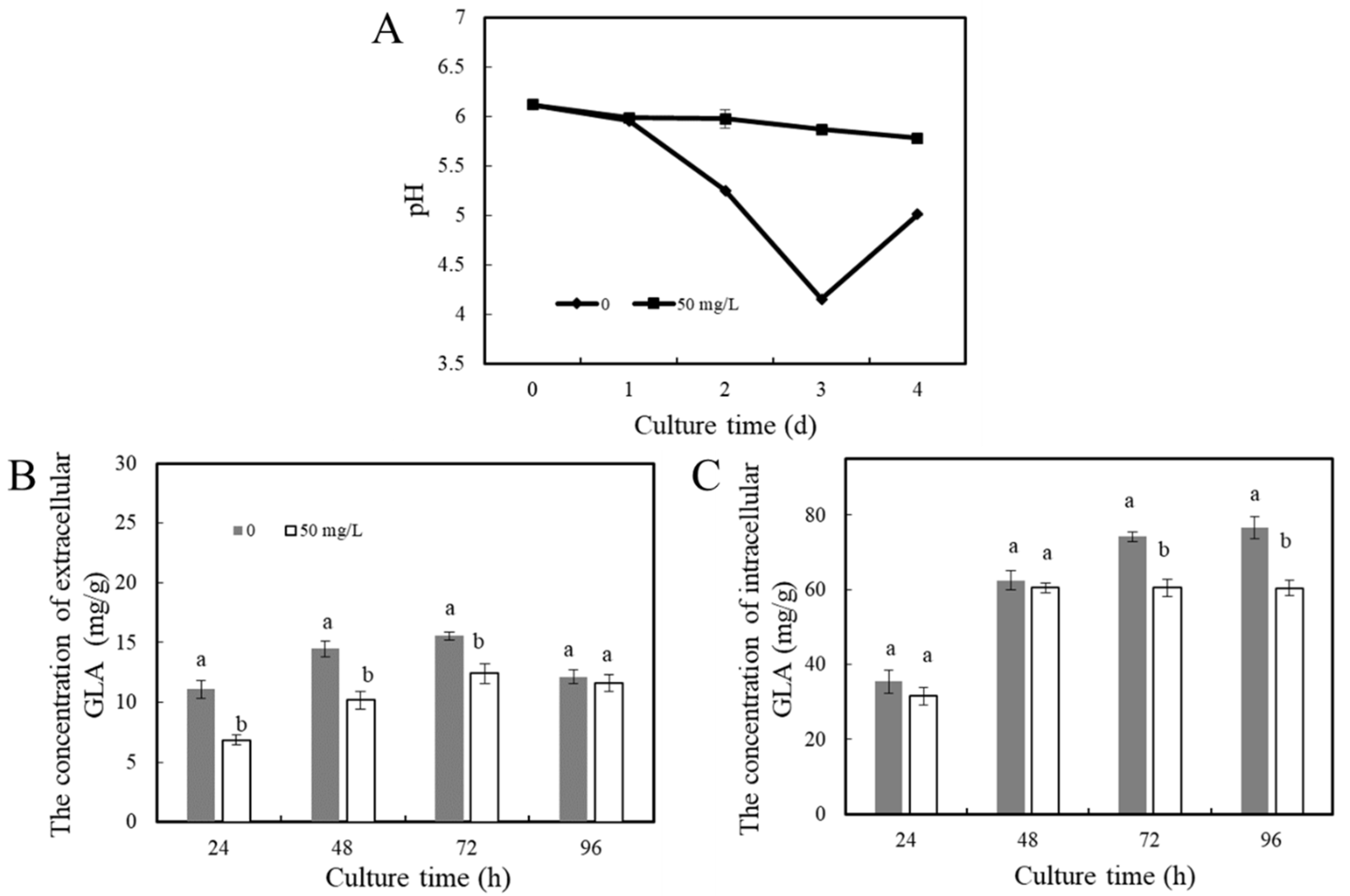
3.2.1. Environmental pH Change In Vitro
Acid Production during the Growth of P. expansum
Environmental pH Change by Aniline Blue Staining Method
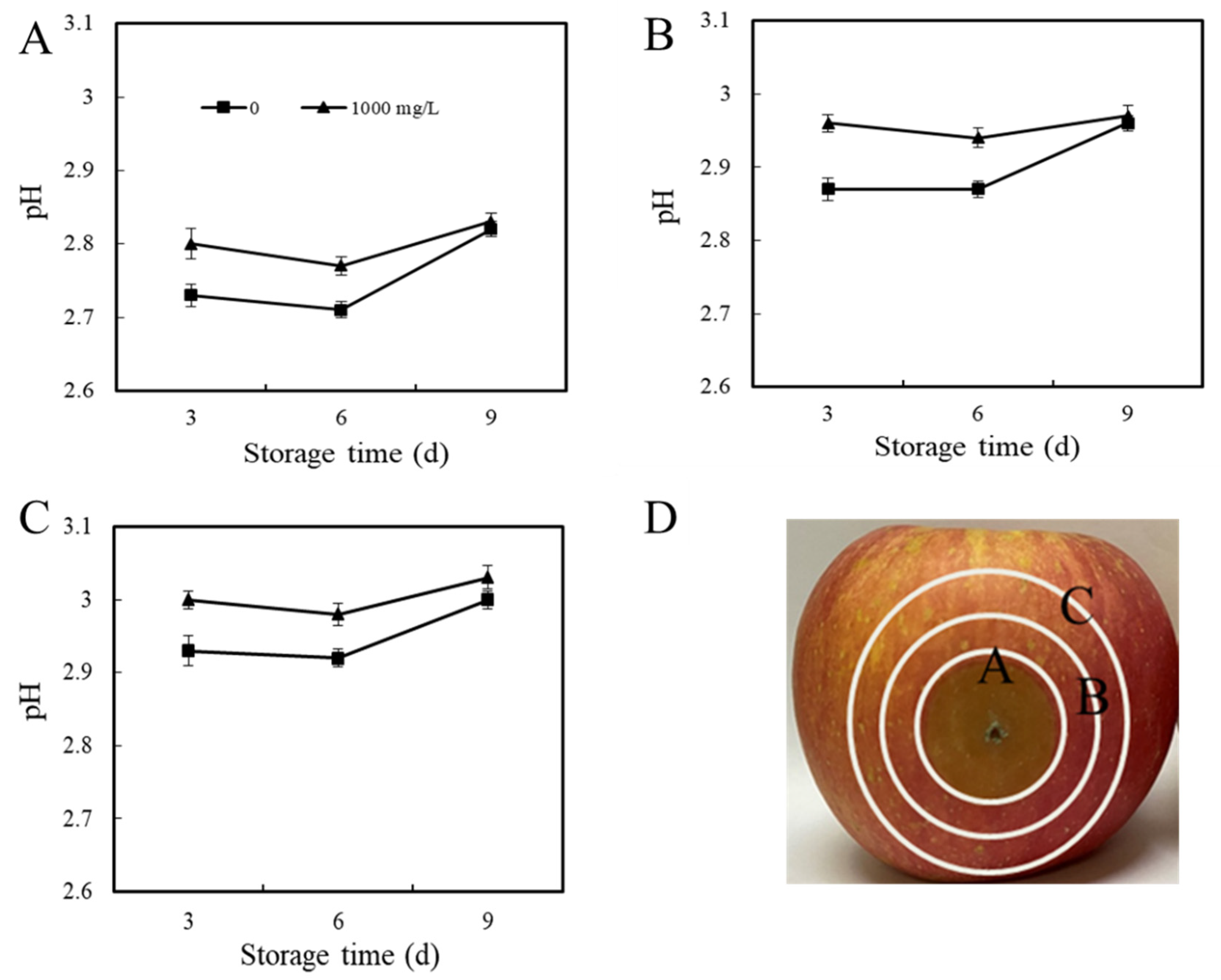
3.2.2. Environmental pH Change In Vivo
Environmental pH Change in P. expansum–Apple Interaction Areas
Organic Acids in P. expansum–Apple Interaction Areas
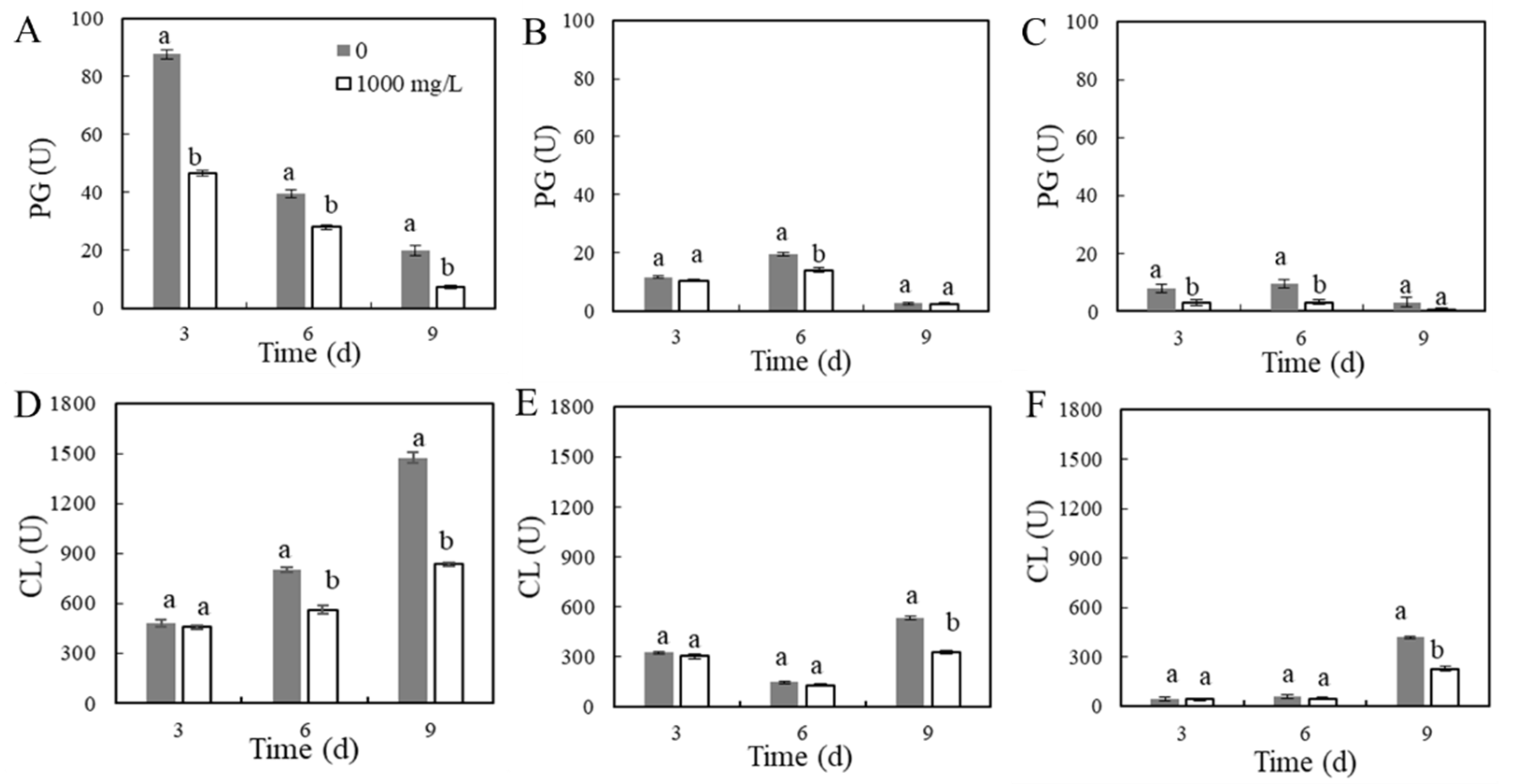
GOX Activity in P. expansum–Apple Interaction Areas
3.3. Cell-Wall-Degrading Enzyme Activities in P. expansum–Apple Interaction Areas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.Y.; Zhang, G.C.; Li, P.X.; Yang, Q.Y.; Chen, K.P.; Zhao, L.N.; Apaliya, M.T.; Gu, X.; Zhang, H.Y. Mechanisms of glycine betaine enhancing oxidative stress tolerance and biocontrol efficacy of Pichia caribbica against blue mold on apples. Biol. Control 2017, 108, 55–63. [Google Scholar] [CrossRef]
- Li, J.K.; Lei, H.H.; Song, H.M.; Lai, T.F.; Xu, X.B.; Shi, X.Q. 1-methylcyclopropene (1-MCP) suppressed postharvest blue mold of apple fruit by inhibiting the growth of Penicillium expansum. Posthavest. Biol. Technol. 2017, 125, 59–64. [Google Scholar] [CrossRef]
- Simonato, B.; Lorenzini, M.; Zapparoli, G. Effects of post-harvest fungal infection of apples on chemical characteristics of cider. LWT 2021, 138, 110620. [Google Scholar] [CrossRef]
- Žebeljan, A.; Vico, I.; Duduk, N.; Žiberna, B.; Krajnc, A.U. Dynamic changes in common metabolites and antioxidants during Penicillium expansum-apple fruit interactions. Physiol. Mol. Plant Pathol. 2019, 106, 166–174. [Google Scholar] [CrossRef]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Xu, W.; Zheng, X.; Zhang, X.; Abdelhai, M.H.; Zhao, L.; Li, H.; Diao, J.; Zhang, H. Exploring the effect of β-glucan on the biocontrol activity of Cryptococcus podzolicus against postharvest decay of apples and the possible mechanisms involved. Biol. Control 2018, 121, 14–22. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Bhayani, K.; Ghosh, T.; Bajaj, S.; Trivedi, N.; Mishra, S. Stability of phycobiliproteins using natural preservative ε-Polylysine (ε-PL). Ferment. Technol. 2018, 7, 149. [Google Scholar] [CrossRef]
- Ge, Y.H.; Wei, M.L.; Li, C.Y.; Chen, Y.R.; Lv, J.Y.; Meng, K.; Wang, W.H.; Li, J.R. Reactive oxygen species metabolism and phenylpropanoid pathway involved in disease resistance against Penicillium expansum in apple fruit induced by ε-poly-L-lysine. J. Sci. Food Agric. 2018, 98, 5082–5088. [Google Scholar] [CrossRef]
- Shu, C.; Cui, K.B.; Li, Q.Q.; Cao, J.K.; Jiang, W.B. Epsilon-poly-l-lysine (ε-PL) exhibits multifaceted antifungal mechanisms of action that control postharvest Alternaria rot. Int. J. Food Microbiol. 2021, 348, 109224. [Google Scholar] [CrossRef]
- Jiao, W.X.; Liu, X.; Chen, Q.M.; Du, Y.M.; Li, Y.Y.; Yue, F.L.; Dong, X.Q.; Fu, M.R. Epsilon-poly-l-lysine (ε-PL) exhibits antifungal activity in vivo and in vitro against Botrytis cinerea and mechanism involved. Postharvest Biol. Technol. 2020, 168, 111270. [Google Scholar] [CrossRef]
- Liu, K.W.; Zhou, X.J.; Fu, M.R. Inhibiting effects of epsilon-poly-lysine (ε-PL) on Pencillium digitatum and its involved mechanism. Postharvest Biol. Technol. 2017, 123, 94–101. [Google Scholar] [CrossRef]
- Lin, L.; Gu, Y.L.; Li, C.Z.; Vittayapadung, S.; Cui, H.Y. Antibacterial mechanism of ε-Poly-lysine against Listeria monocytogenes and its application on cheese. Food Control 2018, 91, 76–84. [Google Scholar] [CrossRef]
- Li, H.; He, C.; Li, G.; Zhang, Z.; Li, B.; Tian, S.P. The modes of action of epsilon-polylysine (ε-PL) against Botrytis cinerea in jujube fruit. Postharvest Biol. Technol. 2019, 147, 1–9. [Google Scholar] [CrossRef]
- Qin, G.Z.; Zong, Y.Y.; Chen, Q.L.; Hua, D.L.; Tian, S.P. Inhibitory effect of boron against Botrytis cinerea on table grapes and its possible mechanisms of action. Int. J. Food Microbiol. 2010, 138, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Brito, N.; Espino, J.J.; Gonzalez, C. Endo-β-1,4-xylanase xyn11A required for virulence in Botrytis cinereal. Mol. Plant Microbe Interact. 2006, 19, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Prusky, D.; McEvoy, J.L.; Leverentz, B.; Conway, W.S. Local modulation of host pH by Colletotrichum species as a mechanism to increase virulence. Mol. Plant Microbe Interact. 2001, 14, 1105–1113. [Google Scholar] [CrossRef] [Green Version]
- Bateman, D.F.; Beer, S.V. Simultaneous production and synergistic action of oxalic acid and polygalacturonase during pathogenesis by Sclerotium rolfsii. Phytopathology 1965, 55, 204–211. [Google Scholar]
- Eshel, D.; Miyara, I.; Ailing, T.; Dinoor, A.; Prusky, D. pH regulates endoglucanase expression and virulence of Alternaria alternata persimmon fruit. Mol. Plant Microbe Interact. 2002, 15, 774–779. [Google Scholar] [CrossRef] [Green Version]
- Vilanova, L.; Wisniewski, M.; Norelli, J.; Viñas, I.; Torres, R.; Usall, J.; Phillips, J.; Droby, S.; Teixidó, N. Transcriptomic profiling of apple in response to inoculation with a pathogen (Penicillium expansum) and a non-pathogen (Penicillium digitatum). Plant Mol. Biol. 2014, 32, 566–583. [Google Scholar] [CrossRef]
- Prusky, D.; Yakoby, N. Pathogenic fungi: Leading or led by ambient pH? Mol. Plant Pathol. 2003, 4, 509–516. [Google Scholar] [CrossRef]
- Manteau, S.; Abouna, S.; Lambert, B.; Legendre, L. Differential regulation by ambient pH of putative virulence factor secretion by the phytopathogenic fungus Botrytis cinerea. FEMS Microbiol. Ecol. 2003, 43, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Prusky, D.; Mcevoy, J.L.; Saftner, R.; Conway, W.S.; Jones, R. Relationship between host acidification and virulence of Penicillium spp. on apple and citrus fruit. Phytopathology 2004, 94, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollins, J.A.; Dickman, M.B. pH signaling in Sclerotinia sclerotiorum: Identification of a pacC/RIM1 homolog. Appl. Environ. Microbiol. 2001, 67, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidzon, M.; Alkan, N.; Kobiler, I.; Kobiler, I.; Prusky, D. Acidification by gluconic acid of mango fruit tissue during colonization via stem end infection by Phomopsis mangiferae. Postharvest Biol. Technol. 2010, 55, 71–77. [Google Scholar] [CrossRef]
- Deng, Y.; Wu, Y.; Li, M.; Yang, C.; Shi, C.; Zheng, C.J. Studies of postharvest berry abscission of “Kyoho” table grapes during cold storage and high oxygen atmospheres. Postharvest Biol. Technol. 2007, 43, 95–101. [Google Scholar] [CrossRef]
- Nakano, T.; Ito, Y. Molecular mechanisms controlling plant organ abscission. Plant Biotechnol. 2013, 30, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, R.; Ma, W.; Huang, X.; Li, M.; Chen, Q. Yong Enhanced saccharification of SO2 catalyzed steam-exploded corn stover by polyethylene glycol addition. Biomass Bioenergy 2011, 35, 2053–2058. [Google Scholar] [CrossRef]
- McCleary, D.; Mangan, R.; Daly, S.; Fort, R.; Ivory, N. Novel substrates for the measurement of endo-1,4-β-glucanase (endo-cellulase). Carbohyd. Res. 2014, 385, 9–17. [Google Scholar] [CrossRef]
- Zheng, B.B.; Zhao, L.; Jiang, X.H.; Cheronoac, S.; Liu, J.J.; Ogutuad, C.; Ntini, C.; Zhang, X.J.; Han, Y.P. Assessment of organic acid accumulation and its related genes in peach. Food Chem. 2020, 334, 127567. [Google Scholar] [CrossRef]
- Askarne, L.; Talibi, I.; Boubaker, H.; Boudyach, E.H.; Msanda, F.; Saadi, B.; Serghini, M.A.; Ait Ben Aoumar, A. In vitro and in vivo antifungal activity of several Moroccan plants against Penicillium italicum, the causal agent of citrus blue mold. Crop Prot. 2012, 40, 53–58. [Google Scholar] [CrossRef]
- Liu, X.; Jiao, W.X.; Du, Y.M.; Chen, Q.M.; Su, Z.B.; Fu, M.R. Chlorine dioxide controls green mold caused by Penicillium digitatum in citrus fruits and the mechanism involved. J. Agric. Food Chem. 2020, 68, 13897–13905. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Nie, J.Y.; Jing, L.I.; Zhang, H.; Li, Y.; Saqib, F.; Syed, A.S.B.; Wang, J. Evaluation of sugar and organic acid composition and their levels in highbush blueberries from two regions of china. J. Integr. Agric. 2020, 19, 2352–2361. [Google Scholar] [CrossRef]
- Wu, J.Y.; Fan, J.B.; Li, Q.H.; Jia, L.T.; Xu, L.L.; Wu, X.; Wang, Z.W.; Li, H.Y.; Qi, K.J.; Qiao, X.; et al. Variation of organic acids in mature fruits of 193 pear (Pyrus spp.) cultivars. J. Food Compos. Anal. 2022, 109, 104483. [Google Scholar] [CrossRef]
- Wei, M.L.; Ge, Y.H.; Li, C.Y.; Chen, Y.R.; Wang, W.H.; Duan, B.; Li, X. Antifungal activity of ε-poly-L-lysine on Trichothecium roseum in vitro and its mechanisms. Physiol. Mol. Plant Pathol. 2018, 103, 23–27. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; Jiao, X.; Li, K.H.; Sun, Y.J.; Zhou, W.L.; Shen, Y.L.; Qian, J.; Chang, A.P.; Wang, J.Q.; et al. Characterization of the biosynthetic pathway of nucleotide sugar precursor UDP-glucose during sphingan WL gum production in Sphingomonas sp. WG. J. Biotechnol. 2019, 302, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Li, W.S.; Yuan, S.Z.; Li, Q.Q.; Sang, W.N.; Cao, J.K.; Jiang, W.B. Methyl p-coumarate inhibits black spot rot on jujube fruit through membrane damage and oxidative stress against Alternaria alternata. Postharvest Biol. Technol. 2018, 145, 230–238. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.Y.; Chen, T.; Xu, Y.; Tian, S.P. Antifungal effects of hinokitiol on development of Botrytis cinerea in vitro and in vivo. Postharvest Biol. Technol. 2020, 159, 111038. [Google Scholar] [CrossRef]
- Dou, Y.; Routledge, M.N.; Gong, Y.Y.; Godana, E.A.; Dhanasekaran, S.; Yang, Q.Y.; Zhang, X.Y.; Zhang, H.Y. Efficacy of epsilon-poly-L-lysine inhibition of postharvest blue mold in apples and potential mechanisms. Postharvest Biol. Technol. 2021, 171, 111346. [Google Scholar] [CrossRef]
- Bo, T.; Liu, M.; Zhong, C.; Zhang, Q.; Su, Q.Z.; Tan, Z.L.; Jia, S.R. Metabolomic analysis of antimicrobial mechanisms of ε-poly-L-lysine on Saccharomyces cerevisiae. J. Agric. Food Chem. 2014, 62, 4454–4465. [Google Scholar] [CrossRef]
- Vilanova, L.; Viñas, I.; Torres, R.; Usall, J.; Buron-Moles, G.; Teixidó, N. Acidification of apple and orange hosts by Penicillium digitatum and Penicillium expansum. Int. J. Food Microbiol. 2014, 178, 39–49. [Google Scholar] [CrossRef]
- Hadas, Y.; Goldberg, I.; Pines, O.; Prusky, D. Involvement of gluconic acid and glucose oxidase in the pathogenicity of Penicillium expansum in apples. Phytopathology 2008, 97, 384–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prusky, D.; Lichter, A. Mechanisms modulating fungal attack in post-harvest pathogen interactions and their control. Eur. J. Plant Pathol. 2008, 121, 281–289. [Google Scholar] [CrossRef]
- Hématy, K.; Cherk, C.; Somerville, S. Host-pathogen warfare at the plant cell wall. Curr. Opin. Plant Biol. 2009, 12, 406–413. [Google Scholar] [CrossRef] [PubMed]
- D’Ovidio, R.; Mattei, B.; Roberti, S.; Bellincampi, D. Polygalacturonases, polygalacturonase-inhibiting proteins and pectic oligomers in plant–pathogen interactions. BBA-Proteins Proteom. 2004, 1696, 237–244. [Google Scholar] [CrossRef]
- Xu, M.Q.; Yang, Q.Y.; Boateng, N.A.S.; Ahima, J.; Dou, Y.; Zhang, H.Y. Ultrastructure observation and transcriptome analysis of Penicillium expansum invasion in postharvest pears. Postharvest Biol. Technol. 2020, 165, 111198. [Google Scholar] [CrossRef]
- Luca, S.; Carla, C.; Serena, R.; Renato, D.; Francesco, F. An endo-polygalacturonase (pg) of fusarium moniliforme escaping inhibition by plant polygalacturonase-inhibiting proteins (pgips) provides new insights into the pg-pgip interaction. FEMS Microbiol. Lett. 2004, 1, 117–124. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | ε-PL (mg/L) | 3 d | 6 d | 9 d | (mg/g) |
|---|---|---|---|---|---|
| A | 0 | 17.72 ± 0.15 a | 13.47 ± 0.28 a | 11.17 ± 0.13 a | Gluconic acid |
| 1000 | 13.07 ± 0.23 b | 11.48 ± 0.15 b | 9.83 ± 0.24 b | ||
| B | 0 | 11.43 ± 0.34 c | 9.9 ± 0.24 c | 9.37 ± 0.19 bc | |
| 1000 | 10.39 ± 0.41 d | 9.06 ± 0.25 d | 9.05 ± 0.21 cd | ||
| C | 0 | 9.47 ± 0.28 e | 8.83 ± 0.17 de | 8.96 ± 0.28 cd | |
| 1000 | 8.24 ± 0.19 f | 8.38 ± 0.14 e | 8.52 ± 0.20 d | ||
| A | 0 | 9.11 ± 0.21 c | 8.61 ± 0.15 b | 7.61 ± 0.12 d | Malic acid |
| 1000 | 10.60 ± 0.13 b | 9.99 ± 0.22 a | 8.62 ± 0.17 a | ||
| B | 0 | 9.96 ± 0.17 b | 8.84 ± 0.13 b | 7.70 ± 0.20 cd | |
| 1000 | 10.91 ± 0.23 a | 9.75 ± 0.24 a | 8.10 ± 0.15 bc | ||
| C | 0 | 10.00 ± 0.20 b | 8.65 ± 0.17 b | 7.63 ± 0.16 cd | |
| 1000 | 11.27 ± 0.21 a | 9.59 ± 0.19 a | 8.50 ± 0.23 ab | ||
| A | 0 | 0.64 ± 0.05 a | 0.20 ± 0.02 b | 0.28 ± 0.01 a | Citric acid |
| 1000 | 0.59 ± 0.03 a | 0.15 ± 0.03 bc | 0.15 ± 0.02 b | ||
| B | 0 | 0.33 ± 0.04 b | 0.15 ± 0.02 bc | 0.07 ± 0.04 c | |
| 1000 | 0.26 ± 0.04 b | 0.35 ± 0.02 a | 0.10 ± 0.03 bc | ||
| C | 0 | 0.09 ± 0.03 c | 0.11 ± 0.02 c | 0.07 ± 0.03 c | |
| 1000 | 0.11 ± 0.03 c | 0.12 ± 0.02 c | 0.09 ± 0.02 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Jiao, W.; Du, Y.; Wang, M.; Fu, M. Involvement of Organic Acid in the Control Mechanism of ε-Poly-L-lysine (ε-PL) on Blue Mold Caused by Penicillium expansum in Apple Fruits. Horticulturae 2022, 8, 468. https://doi.org/10.3390/horticulturae8060468
Li Y, Jiao W, Du Y, Wang M, Fu M. Involvement of Organic Acid in the Control Mechanism of ε-Poly-L-lysine (ε-PL) on Blue Mold Caused by Penicillium expansum in Apple Fruits. Horticulturae. 2022; 8(6):468. https://doi.org/10.3390/horticulturae8060468
Chicago/Turabian StyleLi, Youyuan, Wenxiao Jiao, Yamin Du, Min Wang, and Maorun Fu. 2022. "Involvement of Organic Acid in the Control Mechanism of ε-Poly-L-lysine (ε-PL) on Blue Mold Caused by Penicillium expansum in Apple Fruits" Horticulturae 8, no. 6: 468. https://doi.org/10.3390/horticulturae8060468
APA StyleLi, Y., Jiao, W., Du, Y., Wang, M., & Fu, M. (2022). Involvement of Organic Acid in the Control Mechanism of ε-Poly-L-lysine (ε-PL) on Blue Mold Caused by Penicillium expansum in Apple Fruits. Horticulturae, 8(6), 468. https://doi.org/10.3390/horticulturae8060468