
The Variation of Soil Phosphorus Fractions and Microbial Community Composition under Consecutive Cucumber Cropping in a Greenhouse

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Design
2.2. Soil Sampling
2.3. Chemical Analyses in Soil and Plants
2.4. Soil Phosphorus Fractionation and Phosphatase Activity
2.5. Soil of Phospholipid-Derived Fatty Acids (PLFAs)
2.6. Statistics Analysis
3. Results
3.1. Uptake and Distribution of P in Plants
3.2. Soil Chemical Properties
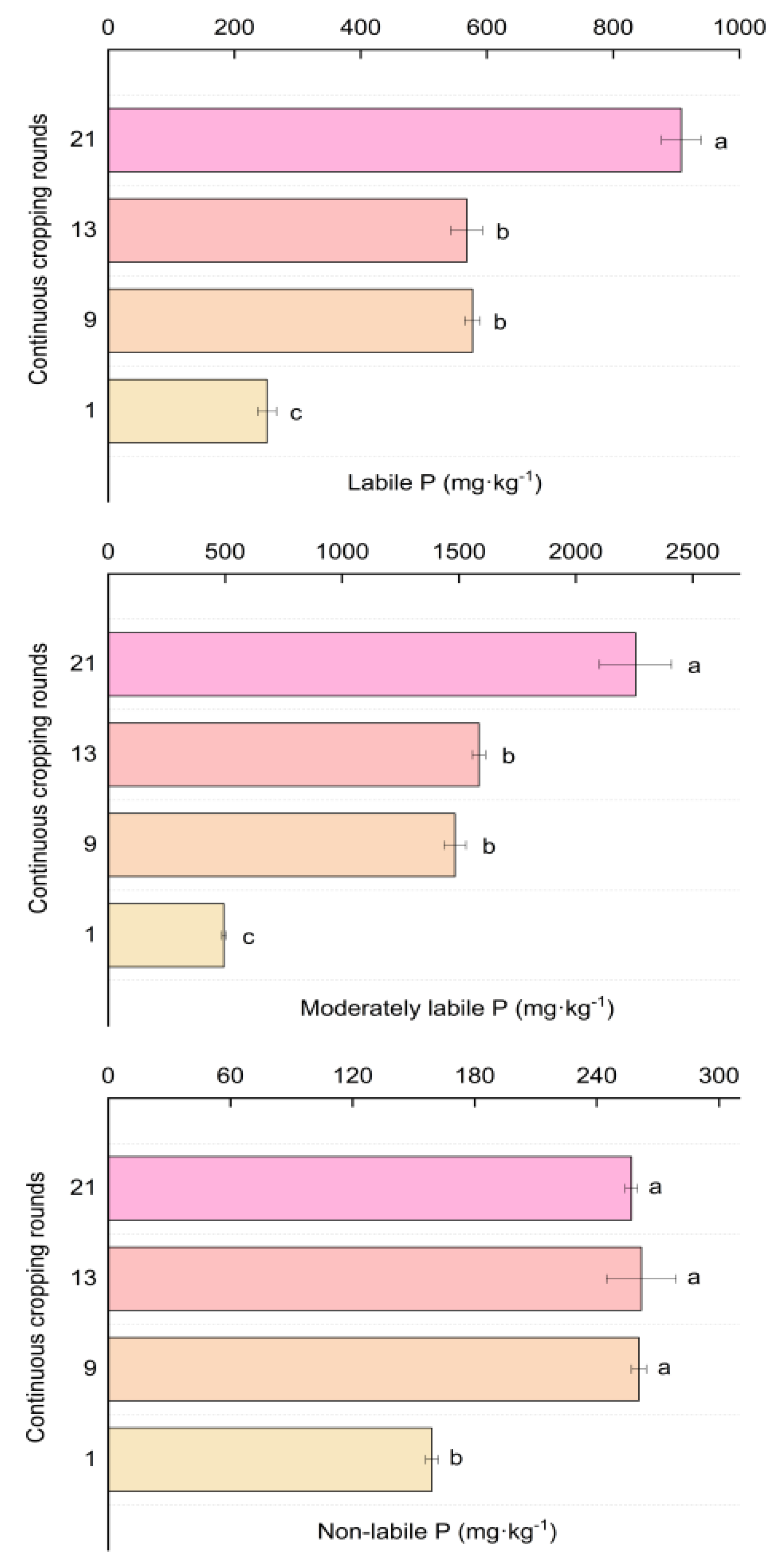
3.3. Soil Phosphorus Fractions
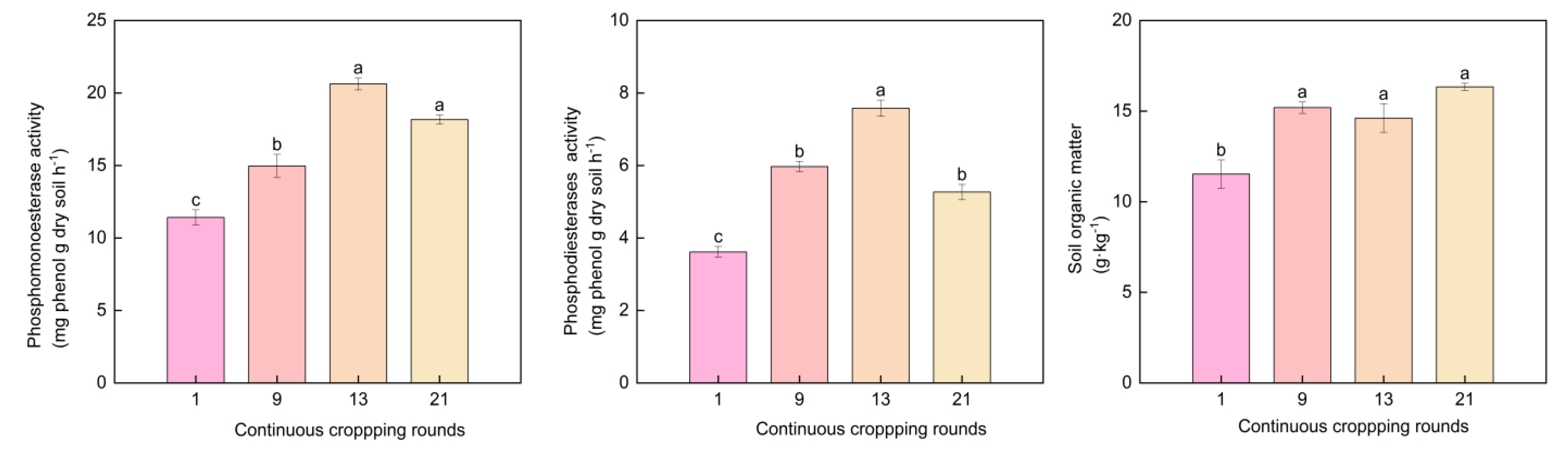
3.4. Soil Phosphatase Activity and Soil Organic Matter
3.5. Soil Microbial Community Composition
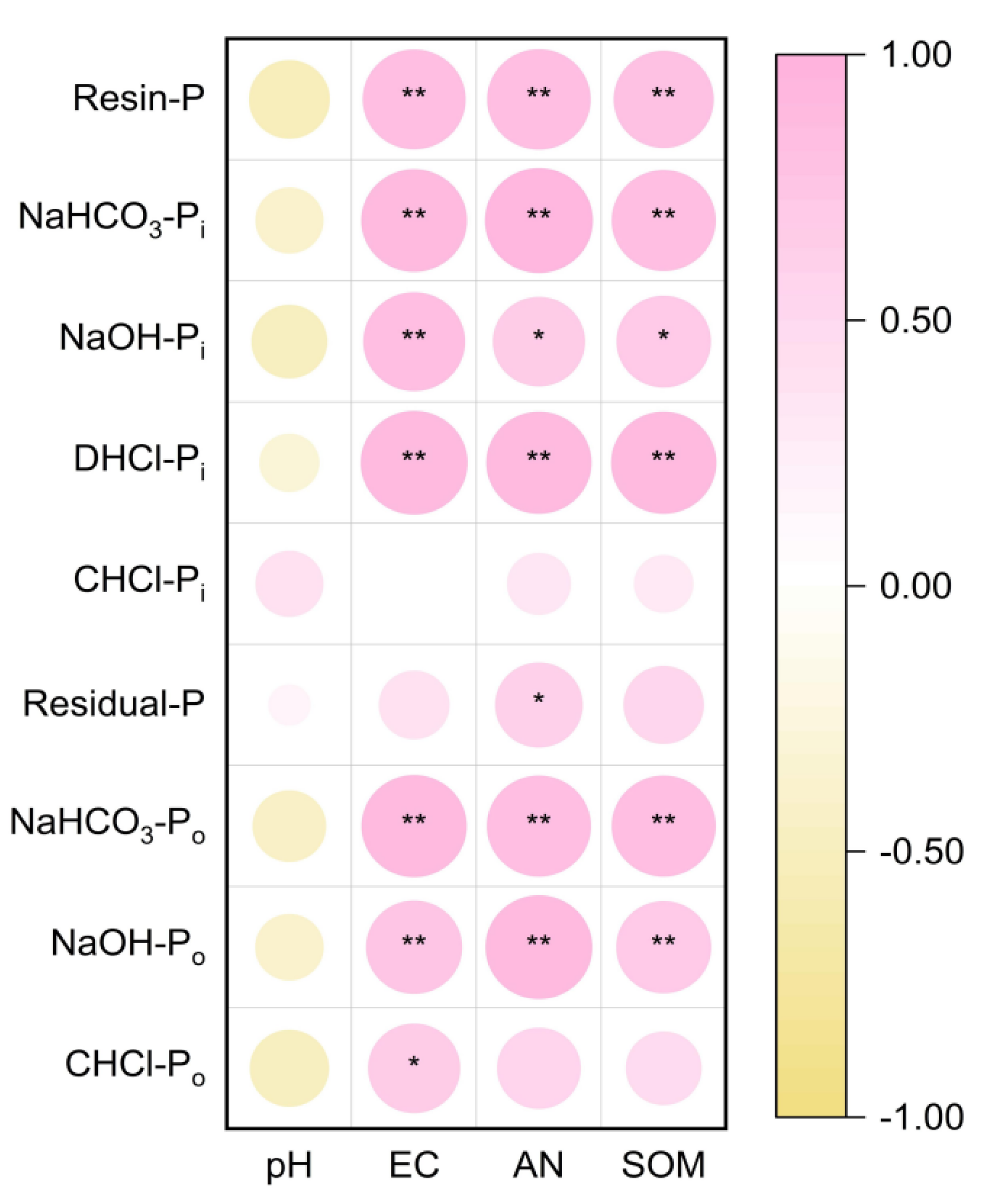
3.6. The Relationships between PLFA, Soil Parameters, and P Fractions
4. Discussions
4.1. Phosphorus Accumulation in the Soil of Solar Greenhouses
4.2. Effects of Continuous Cropping on Soil Microbial Community Structure
4.3. Correlation Analysis of the Soil P Fractions, Soil Microbial Community Structure, and Soil Chemical Properties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, K.; Xue, Y.; Zheng, X.Q.; Lv, W.G.; Yang, J.J. Effects of the continuous use of organic manure and chemical fertilizer on soil inorganic phosphorus fractions in calcareous soil. Sci. Rep. 2017, 7, 1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, S.B.; Dzidzienyo, D.K.; Yahaya, A.; Umar, M.L.; Ishiyaku, M.F.; Tongoona, P.B.; Gracen, V. High soil phosphorus application significantly increased grain yield, phosphorus content but not zinc content of cowpea grains. Agronomy 2021, 11, 802. [Google Scholar] [CrossRef]
- Mai, W.X.; Xue, X.G.; Feng, G.; Yang, R.; Tian, C.Y. Can optimization of phosphorus input lead to high productivity and high phosphorus use efficiency of cotton through maximization of root/mycorrhizal efficiency in phosphorus acquisition? Field Crops Res. 2018, 216, 100–108. [Google Scholar] [CrossRef]
- Carneiro, J.S.S.; Ribeiro, I.C.A.; Nardis, B.O.; Barbosa, C.F.; Lustosa Filho, J.F.; Melo, L.C.A. Long-Term Effect of Biochar-Based Fertilizers Application in Tropical Soil: Agronomic Efficiency and Phosphorus Availability. Sci. Total Environ. 2021, 760, 143955. [Google Scholar] [CrossRef] [PubMed]
- Hallama, M.; Pekrun, C.; Lambers, H.; Kandeler, E. Hidden miners-the roles of cover crops and soil microorganisms in phosphorus cycling through agroecosystems. Plant Soil 2019, 434, 7–45. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.Y.; Zhang, Y.X.; Huang, B.; Teng, Y. Soil environmental quality in greenhouse vegetable production systems in eastern china: Current status and management strategies. Chemosphere 2017, 170, 183–195. [Google Scholar] [CrossRef]
- Qin, H.L.; Quan, Z.; Liu, X.L.; Ming-De, L.I.; Zong, Y.; Jin-Shui, W.U. Phosphorus status and risk of phosphate leaching loss from vegetable soils of different planting years in suburbs of Changsha, China. Agric. Sci. China 2010, 9, 1641–1649. [Google Scholar] [CrossRef]
- Preston, C.L.; Ruiz Diaz, D.A.; Mengel, D.B. Corn response to longterm phosphorus fertilizer application rate and placement with strip-tillage. Agron. J. 2019, 111, 841–850. [Google Scholar] [CrossRef]
- Ahmed, W.; Liu, K.L.; Qaswar, M.; Huang, J.; Zhang, H. Long-term mineral fertilization improved the grain yield and phosphorus use efficiency by changing soil p fractions in ferralic cambisol. Agron. J. 2019, 9, 784. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.H.; Zhang, S.X.; Zhu, P.; Huang, S.M.; Wang, B.R.; Zhao, L.P.; Xu, M.G. Characterizing differences in the phosphorus activation coefficient of three typical cropland soils and the influencing factors under long-term fertilization. PLoS ONE 2017, 12, e0176437. [Google Scholar] [CrossRef]
- Yan, X.; Wei, Z.; Hong, Q.; Lu, Z.; Wu, J. Phosphorus fractions and sorption characteristics in a subtropical paddy soil as influenced by fertilizer sources. Geoderma 2017, 295, 80–85. [Google Scholar] [CrossRef]
- Chang, H.L.; Chang, Y.P.; Park, K.D.; Jeon, W.T.; Kim, P.J. Long-term effects of fertilization on the forms and availability of soil phosphorus in rice paddy. Chemosphere 2004, 56, 299–304. [Google Scholar]
- Van, A.H.C.; Goss, E.M.; Havelaar, A.; Van, A.D.; Finckh, M.R.; Morris, J.G. One health-cycling of diverse microbial communities as a connecting force for soil, plant, animal, human and ecosystem health. Sci. Total Environ. 2019, 664, 927–937. [Google Scholar]
- Gao, Y.H.; Lu, X.H.; Guo, R.J.; Hao, J.J.; Miao, Z.Q.; Li, Y.; Shi, D.L. Responses of soil abiotic properties and microbial community structure to 25-year cucumber monoculture in commercial greenhouses. Agriculture 2021, 11, 341. [Google Scholar] [CrossRef]
- Ding, G.C.; Bai, M.; Han, H.; Li, H.; Ding, X.; Yang, H.; Xu, T.; Li, J. Microbial taxonomic, nitrogen cycling and phosphorus recycling community composition during long-term organic greenhouse farming. FEMS Microbiol. Ecol. 2019, 95, fiz042. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.L.; Jing, F.; Sun, W.Q.; Liu, J.G.; Jiang, G.Y. Soil microbial communities changed with a continuously monocropped processing tomato system. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2018, 68, 149–160. [Google Scholar] [CrossRef]
- Shang, Q.Y.; Yang, X.X.; Yan, H.; Wang, X.H. Integrated soil fertility and yield trend in response to long-term fertilisation under the Chinese double rice-cropping systems. Plant Soil Environ. 2020, 66, 22–32. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; United States Department of Agriculture (USDA) Circular No.939; US Department of Agriculture: Washington, DC, USA, 1954.
- Hedley, M.J.; Stewart, J.W.B.; Chauhan, B. Changes in inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubations. Soil Sci. Soc. Am. J. 1982, 46, 970–976. [Google Scholar] [CrossRef]
- Tiessen, H.; Moir, J.O. Characterization of available P by sequential extraction. Soil Sampl. Methods Anal. 1993, 7, 5–229. [Google Scholar]
- Guan, S.Y. Soil Enzymes and Research Methods; Agriculture Press: Beijing, China, 1986. [Google Scholar]
- Cavigelli, M.A.; Robertson, G.P.; Klug, M.J. Fatty acid methyl ester (fame) profiles as measures of soil microbial community structure. Plant Soil 1995, 170, 99–113. [Google Scholar] [CrossRef]
- Fu, H.D.; Zhang, G.X.; Zhang, F.; Sun, Z.P.; Geng, G.M.; Li, T.L. Effects of continuous tomato monoculture on soil microbial properties and enzyme activities in a solar greenhouse. Sustainability 2017, 9, 317. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.C.; Guo, S.H.; Zhai, L.M.; Pan, J.T.; Khoshnevisan, B.; Wu, S.X.; Wang, H.Y.; Yang, B.; Ji, J.H.; Liu, H.B. How long-term excessive manure application affects soil phosphorous species and risk of phosphorous loss in fluvo-aquic soil. Environ. Pollut. 2020, 266 Pt 2, 115304. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.H.; Zhang, S.X.; Ren, Y.; Zhan, X.Y.; Xu, M.G.; Feng, G. Soil phosphorus management based on the agronomic critical value of Olsen P. Commun. Soil Sci. Plant Anal. 2018, 49, 934–944. [Google Scholar] [CrossRef]
- Zhang, W.W.; Wang, Q.; Wu, Q.; Zhang, S.X.; Zhu, P.; Peng, C.; Huang, S.M.; Wang, B.R.; Zhang, H. The response of soil Olsen-P to the P budgets of three typical cropland soil types under long-term fertilization. PLoS ONE 2020, 15, e0230178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Z.H.; Li, H.G.; Yang, X.; Zhou, B.K.; Shi, X.J.; Wang, B.R.; Li, D.C.; Shen, J.B.; Chen, Q.; Qin, W.; et al. The critical soil P levels for crop yield, soil fertility and environmental safety in different soil types. Plant Soil 2013, 372, 27–37. [Google Scholar] [CrossRef]
- Liu, C.; Dang, X.L.; Mayes, M.A.; Chen, L.L.; Zhang, Y.L.; Anup, D. Effect of long-term irrigation patterns on phosphorus forms and distribution in the brown soil zone. PLoS ONE 2017, 12, e0188361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.Q.; Dalal, R.C.; Bhattacharyya, R.; Meyer, G.; Wang, P.; Menzies, N.W.; Kopittke, P.M. Effect of long-term no-tillage and nitrogen fertilization on phosphorus distribution in bulk soil and aggregates of a Vertisol. Soil Till. Res. 2021, 205, 104760. [Google Scholar] [CrossRef]
- Bai, X.L.; Gao, J.J.; Wang, S.C.; Cai, H.M.; Chen, Z.J.; Zhou, B. Excessive nutrient balance surpluses in newly built solar greenhouses over five years leads to high nutrient accumulations in soil. Agr. Ecosyst. Environ. 2019, 288, 106717. [Google Scholar] [CrossRef]
- Yang, Y.J.; Zhang, H.P.; Qian, X.Q.; Duan, J.N.; Wang, G.L. Excessive application of pig manure increases the risk of P loss in calcic cinnamon soil in China. Sci. Total Environ. 2017, 609, 102–108. [Google Scholar] [CrossRef]
- Liao, D.; Zhang, C.C.; Li, H.; Lambers, H.; Zhang, F.S. Changes in soil phosphorus fractions following sole cropped and intercropped maize and faba bean grown on calcareous soil. Plant Soil 2020, 463, 589–600. [Google Scholar] [CrossRef]
- Wierzbowska, J.; Sienkiewicz, S.; Zalewska, M.; Żarczyński, P.; Krzebietke, S. Phosphorus fractions in soil fertilised with organic waste. Environ. Monit Assess. 2020, 92, 315. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, P.S.; Condron, L.M.; Di, H.J.; Sinaj, S.; McLenaghen, R.D. Effect of green manure addition on soil organic phosphorus mineralisation. Nutr. Cycl. Agroecosyst. 2005, 73, 181–189. [Google Scholar] [CrossRef]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Cao, N.; Chen, X.; Cui, Z. Change in soil available phosphorus in relation to the phosphorus budget in China. Nutr. Cycl. Agroecosyst. 2012, 94, 161–170. [Google Scholar] [CrossRef]
- Coelho, M.; Diaz, D.R.; Hettiarachchi, G.M.; Hansel, F.D.; Pavinato, P.S. Soil phosphorus fractions and legacy in a corn-soybean rotation on mollisols in kansas, USA. Geoderma Regional. 2019, 18, e00228. [Google Scholar] [CrossRef]
- Zicker, T.; Von Tucher, S.; Kavka, M.; Eichler, L.; Bermann, B. Soil test phosphorus as affected by phosphorus budgets in two long-term field experiments in Germany. Field Crop Res. 2018, 218, 158–170. [Google Scholar] [CrossRef]
- Ahmed, W.; Huang, J.; Liu, K.; Qaswar, M.; Zhang, H. Changes in phosphorus fractions associated with soil chemical properties under long-term organic and inorganic fertilization in paddy soils of southern China. PLoS ONE 2019, 14, e0216881. [Google Scholar] [CrossRef]
- Singh, M.; Reddy, K.S.; Singh, V.; Rupa, T. Phosphorus availability to rice (Oriza sativa L.)-wheat (Triticum estivum L.) in a Vertisol after eight years of inorganic and organic fertilizer additions. Bioresour. Technol. 2007, 98, 1474–1481. [Google Scholar] [CrossRef]
- Rong, Q.L.; Li, R.N.; Huang, S.W. Soil microbial characteristics and yield response to partial substitution of chemical fertilizer with organic amendments in greenhouse vegetable production. J. Integr. Agric. 2018, 17, 1432–1444. [Google Scholar] [CrossRef]
- Liu, X.; Li, Y.J.; Ren, X.J.; Chen, B.H.; Zhang, Y.; Shen, C.W.; Wang, F.; Wu, D.F. Long-Term Greenhouse Cucumber Production Alters Soil Bacterial Community Structure. J. Soil Sci. Plant Nutr. 2020, 20, 306–321. [Google Scholar] [CrossRef]
- Bartelt, R.J.; Joshi, J.; Schmid, B.; Brandl, H.; Balser, T. Soil feedbacks of plant diversity on soil microbial communities and subsequent plant growth. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 27–49. [Google Scholar] [CrossRef]
- Wang, J.C.; Ren, C.; Cheng, H.; Zou, Y.; Bughio, M.A.; Li, Q. Conversion of rainforest into agroforestry and monoculture plantation in China: Consequences for soil phosphorus forms and microbial community. Sci. Total Environ. 2017, 595, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Plette, A.C.; Benedetti, M.F.; Van Riemsdijk, W.H. Competitive binding of protons, calcium, cadmium, and zinc to isolated cell walls of a gram-positive soil bacterium. Environ. Sci. Technol. 1996, 30, 1902–1910. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Firestone, M.K. Microbial community utilization of recalcitrant and simple carbon compounds: Impact of oak-woodland plant communities. Oecologia 2004, 138, 275–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.G.; Gao, D.M.; Liu, J. Changes in rhizosphere soil microbial communities in a continuously monocropped cucumber (Cucumis sativus L.) system. Eur. J. Soil Biol. 2014, 60, 1–8. [Google Scholar] [CrossRef]
- Stewart, C.E.; Plante, A.F.; Paustian, K.; Conant, R.T.; Six, J. Soil carbon saturation: Linking concept and measurable carbon pools. Soil Sci. Soc. Am. J. 2008, 72, 379–392. [Google Scholar] [CrossRef]
- Rahman, M.H.; Okubo, A.; Sugiyama, S.; Mayland, H.F. Physical, chemical and microbiological properties of an Andisol as related to land use and tillage practice. Soil Tillage Res. 2008, 101, 10–19. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | pH | EC | TC | TN | TP | TK | AN | AP | AK |
|---|---|---|---|---|---|---|---|---|---|
| ms·cm−1 | g·kg−1 | g·kg−1 | g·kg−1 | g·kg−1 | mg·kg−1 | mg·kg−1 | mg·kg−1 | ||
| Original soil | 7.5 | 0.2 | 12.0 | 1.3 | 0.9 | 10.6 | 102.0 | 41.3 | 222.8 |
| Resin-P | NaHCO3-Pi | NaOH-Pi | Dil. HCl-Pi | Conc. HCl-Pi | NaHCO3-Po | NaOH-Po | Conc. HCl-Po | Residual-P | |
| mg·kg−1 | mg·kg−1 | mg·kg−1 | mg·kg−1 | mg·kg−1 | mg·kg−1 | mg·kg−1 | mg·kg−1 | mg·kg−1 | |
| 92.0 | 16.3 | 43.9 | 233.8 | 42.0 | 14.8 | 18.9 | 3.1 | 14.1 |
| Elemental Sources | Elemental Application Amount (kg·ha–1 year–1) | ||
|---|---|---|---|
| N | P2O5 | K2O | |
| Chemical fertilizer | 450 | 142 | 593 |
| Chicken manure | 361 | 152 | 249 |
| Total fertilizer | 812 | 294 | 842 |
| Microbial Type | Phospholipid Fatty Acid Labelled |
|---|---|
| Gram-positive bacteria (G+) | 17:0 iso, 17:0 anteiso, 16:0 iso, 15:0 iso, 15:0 anteiso, 14:0 iso |
| Gram-negative bacteria (G−) | 16:1 w7c, 16:1 w9c, 17:0 cyclo w7c, 17:1 w8c, 18:1 w7c, 19:0 cyclo w7c |
| Actinomycetes | 18:0, 17:0 10-methyl, 16:0 10-methyl, 18:0 10-methyl |
| Fungi | 18:2 w6c, 18:1 w9c, 16:1 w5c |
| Treatments | P Accumulation (g·Plant−1) | P Distribution Proportion (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Fruit | Total | Root | Stem | Leaf | Fruit | |
| 1 | 0.0068 ± 0.0004 a | 0.05 ± 0.01 b | 0.21 ± 0.02 a | 0.19 ± 0.02 a | 0.45 ± 0.02 b | 1.50 ± 0.16 a | 10.49 ± 1.08 c | 47.15 ± 3.04 a | 40.86 ± 3.70 a |
| 9 | 0.0080 ± 0.0010 a | 0.11 ± 0.01 a | 0.22 ± 0.01 a | 0.19 ± 0.01 a | 0.52 ± 0.03 ab | 1.56 ± 0.25 a | 20.09 ± 1.52 a | 42.59 ± 2.08 a | 35.76 ± 0.74 a |
| 13 | 0.0084 ± 0.0010 a | 0.08 ± 0.01 ab | 0.21 ± 0.02 a | 0.29 ± 0.04 a | 0.59 ± 0.01 a | 1.42 ± 0.15 a | 12.96 ± 2.30 bc | 36.53 ± 3.51 a | 49.09 ± 5.64 a |
| 21 | 0.0068 ± 0.0008 a | 0.11 ± 0.01 a | 0.29 ± 0.06 a | 0.18 ± 0.05 a | 0.58 ± 0.04 a | 1.20 ± 0.21 a | 17.99 ± 1.22 ab | 48.96 ± 8.80 a | 31.85 ± 10.02 a |
| Treatments | Bacteria ug·g−1 | Fungi ug·g−1 | Actinomycetes ug·g−1 | G+ ug·g−1 | G− ug·g−1 | F/B | G+/G− | Total PLFA ug·g−1 |
|---|---|---|---|---|---|---|---|---|
| R1 | 33.00 ± 3.12 b | 5.12 ± 0.74 b | 5.91 ± 0.42 c | 13.46 ± 1.55 c | 18.64 ± 1.58 b | 0.15 ± 0.01 b | 0.72 ± 0.02 b | 48.62 ± 5.08 b |
| R9 | 60.11 ± 3.17 a | 10.48 ± 0.54 a | 11.28 ± 0.58 a | 25.51 ± 1.25 a | 32.87 ± 2.06 a | 0.17 ± 0.01 b | 0.78 ± 0.04 b | 88.06 ± 4.95 a |
| R13 | 57.79 ± 4.36 a | 8.89 ± 0.43 a | 11.43 ± 0.53 a | 24.95 ± 1.59 a | 31.35 ± 2.64 a | 0.16 ± 0.01 b | 0.80 ± 0.02 b | 86.33 ± 8.87 a |
| R21 | 41.18 ± 2.59 b | 8.33 ± 0.92 a | 9.14 ± 0.57 b | 18.78 ± 1.38 b | 21.38 ± 1.29 b | 0.22 ± 0.02 a | 0.88 ± 0.01 a | 63.13 ± 4.01 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bian, T.; Zheng, S.; Li, X.; Wang, S.; Zhang, X.; Wang, Z.; Li, X.; Fu, H.; Sun, Z. The Variation of Soil Phosphorus Fractions and Microbial Community Composition under Consecutive Cucumber Cropping in a Greenhouse. Horticulturae 2022, 8, 320. https://doi.org/10.3390/horticulturae8040320
Bian T, Zheng S, Li X, Wang S, Zhang X, Wang Z, Li X, Fu H, Sun Z. The Variation of Soil Phosphorus Fractions and Microbial Community Composition under Consecutive Cucumber Cropping in a Greenhouse. Horticulturae. 2022; 8(4):320. https://doi.org/10.3390/horticulturae8040320
Chicago/Turabian StyleBian, Ting, Shiwei Zheng, Xiao Li, Shuang Wang, Xiaolan Zhang, Zhen Wang, Xiaoxia Li, Hongdan Fu, and Zhouping Sun. 2022. "The Variation of Soil Phosphorus Fractions and Microbial Community Composition under Consecutive Cucumber Cropping in a Greenhouse" Horticulturae 8, no. 4: 320. https://doi.org/10.3390/horticulturae8040320
APA StyleBian, T., Zheng, S., Li, X., Wang, S., Zhang, X., Wang, Z., Li, X., Fu, H., & Sun, Z. (2022). The Variation of Soil Phosphorus Fractions and Microbial Community Composition under Consecutive Cucumber Cropping in a Greenhouse. Horticulturae, 8(4), 320. https://doi.org/10.3390/horticulturae8040320