
QTL Mapping of Resistance to Bacterial Wilt in Pepper Plants (Capsicum annuum) Using Genotyping-by-Sequencing (GBS)

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. DNA Extraction
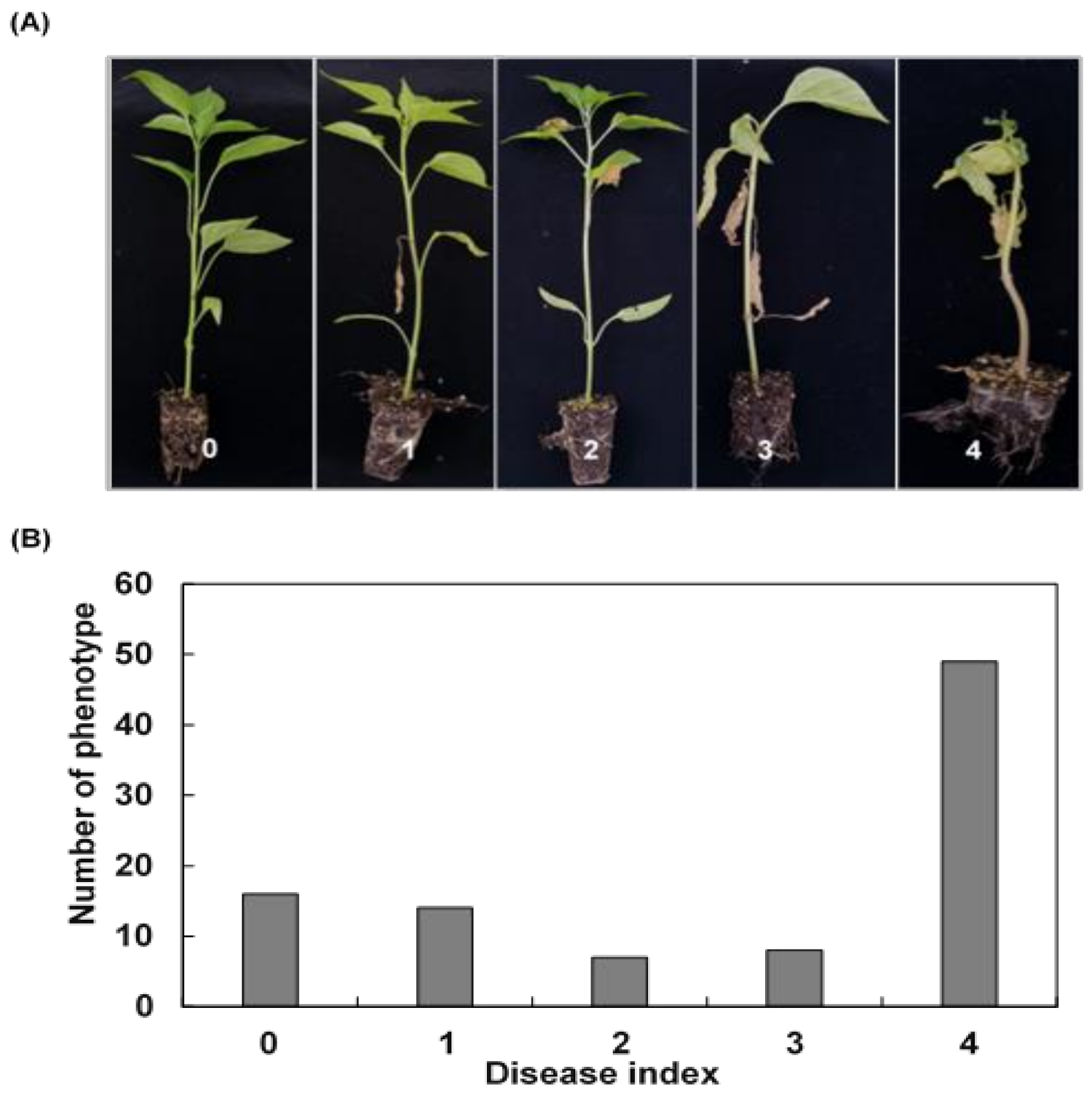
2.3. Disease Assay and Resistance Index Scoring
2.4. Preparation of Libraries for Genotype-by-Sequencing (GBS) Analysis
2.5. Sequencing, Alignment, and SNP Genotyping
2.6. Linkage Map Construction
2.7. QTL Analysis and Candidate Genes Prediction
2.8. Data Analysis
3. Results
3.1. Phenotyping of the Resistance for Bacterial Wilt Disease
3.2. GBS Analysis
3.3. Construction of Linkage Mapping
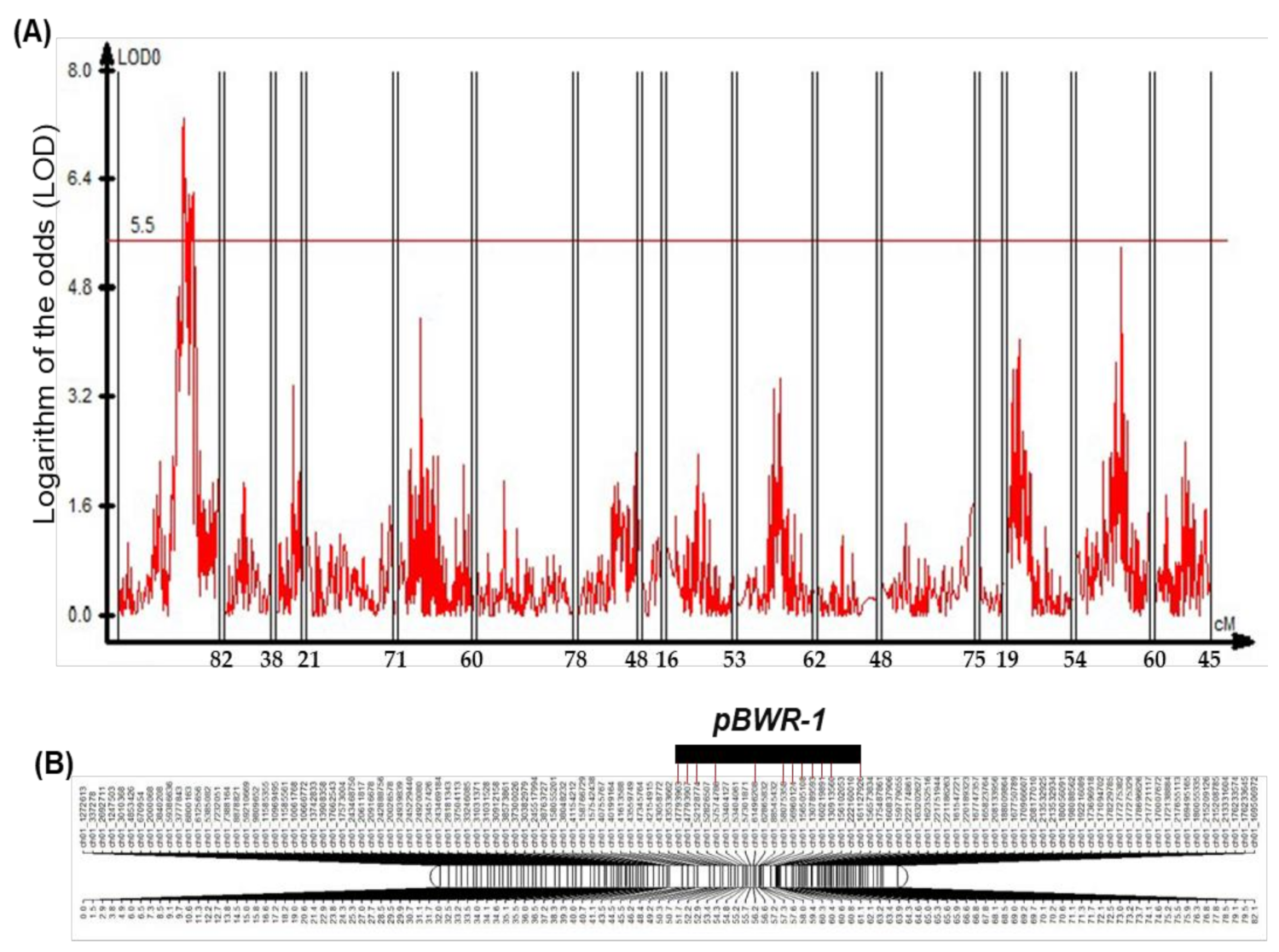
3.4. QTL Analysis for Bacterial Wilt (BW) Resistance
3.5. Prediction and Annotation of Candidate Genes
4. Discussion
4.1. Construction of Pepper Genetic Map
4.2. Genetic Inheritance of pBWR-1 QTL on Chromosome 01
4.3. Detection of Major QTL Controlling Resistance to R. solanacearum
4.4. pBWR-1 Candidate Genes for Resistance to R. solanacearum
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhutia, K.; Khanna, V.; Meetei, T.; Bhutia, N. Effects of climate change on growth and development of chilli. Agrotechnology 2018, 7, 2. [Google Scholar] [CrossRef]
- Prohens, J.; Nuez, F. Handbook of Plant Breeding. Vegetables II: Fabaceae, Liliaceae, Solanaceae and Umbelliferae; Springer: New York, NY, USA, 2008; Volume 3, pp. 30–40. [Google Scholar]
- Fraenkel, L.; Bogardus, S.T.; Concato, J.; Wittink, D.R. Treatment options in knee osteoarthritis: The patient’s perspective. Arch. Intern. Med. 2004, 164, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Xu, Z.; Wu, C.T.; Janes, M.; Prinyawiwatkul, W.; No, H. Antioxidant activities of different colored sweet bell peppers (Capsicum annuum L.). J. Food Sci. 2007, 72, S98–S102. [Google Scholar] [CrossRef] [PubMed]
- Blum, E.; Mazourek, M.; O’connell, M.; Curry, J.; Thorup, T.; Liu, K.; Jahn, M.; Paran, I. Molecular mapping of capsaicinoid biosynthesis genes and quantitative trait loci analysis for capsaicinoid content in Capsicum. Theor. Appl. Genet. 2003, 108, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Stewart Jr, C.; Mazourek, M.; Stellari, G.M.; O’Connell, M.; Jahn, M. Genetic control of pungency in C. chinense via the Pun1 locus. J. Exp. Bot. 2007, 58, 979–991. [Google Scholar] [CrossRef]
- Aza-González, C.; Núñez-Palenius, H.G.; Ochoa-Alejo, N. Molecular biology of capsaicinoid biosynthesis in chili pepper (Capsicum spp.). Plant Cell Rep. 2011, 30, 695–706. [Google Scholar] [CrossRef]
- Luo, X.-J.; Peng, J.; Li, Y.-J. Recent advances in the study on capsaicinoids and capsinoids. Eur. J. Pharmacol. 2011, 650, 1–7. [Google Scholar] [CrossRef]
- Faostat. 2021. Available online: http://www.fao.org (accessed on 9 August 2021).
- Rajametov, S.N.; Lee, K.; Jeong, H.-B.; Cho, M.-C.; Nam, C.-W.; Yang, E.-Y. The Effect of Night Low Temperature on Agronomical Traits of Thirty-Nine Pepper Accessions (Capsicum annuum L.). Agronomy 2021, 11, 1986. [Google Scholar] [CrossRef]
- Comtrade UN. UN Comtrade Database. Available online: http://comtrade.un.org (accessed on 15 October 2020).
- APS. Common Names of Plant Disease. Available online: https://www.apsnet.org/edcenter/resources/commonnames/Pages/default.aspx (accessed on 24 October 2020).
- Jeong, Y.; Kim, J.; Kang, Y.; Lee, S.; Hwang, I. Genetic diversity and distribution of Korean isolates of Ralstonia solanacearum. Plant Dis. 2007, 91, 1277–1287. [Google Scholar] [CrossRef]
- Lee, Y.K.; Kang, H.W. Physiological, biochemical and genetic characteristics of Ralstonia solanacearum strains isolated from pepper plants in Korea. Res. Plant Dis. 2013, 19, 265–272. [Google Scholar] [CrossRef]
- Jiang, G.; Peyraud, R.; Remigi, P.; Guidot, A.; Ding, W.; Genin, S.; Peeters, N. Modeling and experimental determination of infection bottleneck and within-host dynamics of a soil-borne bacterial plant pathogen. bioRxiv 2016, 061408. [Google Scholar] [CrossRef]
- Jiang, G.; Wei, Z.; Xu, J.; Chen, H.; Zhang, Y.; She, X.; Macho, A.P.; Ding, W.; Liao, B. Bacterial wilt in China: History, current status, and future perspectives. Front. Plant Sci. 2017, 8, 1549. [Google Scholar] [CrossRef]
- Kang, Y.J.; Ahn, Y.-K.; Kim, K.-T.; Jun, T.-H. Resequencing of Capsicum annuum parental lines (YCM334 and Taean) for the genetic analysis of bacterial wilt resistance. BMC Plant Biol. 2016, 16, 235. [Google Scholar] [CrossRef]
- Hayward, A. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol. 1991, 29, 65–87. [Google Scholar] [CrossRef]
- Denny, T. Plant pathogenic Ralstonia species. In Plant-Associated Bacteria; Springer: Berlin/Heidelberg, Germany, 2007; pp. 573–644. [Google Scholar]
- Hayward, A. The hosts of Pseudomonas solanacearum. In Bacterial Wilt: The Disease and Its Causative Agent, Pseudomonas solanacearum; CAB International Press: Wallingford, UK, 1994; pp. 9–24. [Google Scholar]
- Denny, T.P. Ralstonia solanacearum—A plant pathogen in touch with its host. Trends Microbiol. 2000, 8, 486–489. [Google Scholar] [CrossRef]
- Vasse, J.; Frey, P.; Trigalet, A. Microscopic studies of intercellular infection and protoxylem invasion of tomato roots by Pseudomonas solanacearum. Mol. Plant Microbe Interact. 1995, 8, 241–251. [Google Scholar] [CrossRef]
- Rahman, M.; Abdullah, H.; Vanhaecke, M. Histopathology of susceptible and resistant Capsicum annuum cultivars infected with Ralstonia solanacearum. J. Phytopathol. 1999, 147, 129–140. [Google Scholar] [CrossRef]
- Wang, J.-F.; Olivier, J.; Thoquet, P.; Mangin, B.; Sauviac, L.; Grimsley, N.H. Resistance of tomato line Hawaii7996 to Ralstonia solanacearum Pss4 in Taiwan is controlled mainly by a major strain-specific locus. Mol. Plant Microbe Interact. 2000, 13, 6–13. [Google Scholar] [CrossRef]
- Winstead, N. Inoculation techniques for evluating resistance to Pseudomonas solanacearum. Phytopathology 1952, 42, 623–634. [Google Scholar]
- Mamphogoro, T.; Babalola, O.; Aiyegoro, O. Sustainable management strategies for bacterial wilt of sweet peppers (Capsicum annuum) and other Solanaceous crops. J. Appl. Microbiol. 2020, 129, 496–508. [Google Scholar] [CrossRef]
- Buddenhagen, I. Bacterial wilt of certain seed-bearing Musa spp. caused by tomato strain of Pseudomonas solanacearum. Phytopathology 1962, 52, 286. [Google Scholar]
- Hayward, A. Characteristics of Pseudomonas solanacearum. J. Appl. Bacteriol. 1964, 27, 265–277. [Google Scholar] [CrossRef]
- Matsunaga, H.; Sato, T.; Monma, S. In Inheritance of bacterial wilt resistance in the sweet pepper cv. Mie-Midori. In Proceedings of the 10th Eucarpia Meeting on Genetics and Breeding of Capsicum and Eggplant, Avignon, France, 7–11 September 1998; p. 172. [Google Scholar]
- Lopes, C.A.; Boiteux, L.S. Biovar-specific and broad-spectrum sources of resistance to bacterial wilt (Ralstonia solanacearum) in Capsicum. Embrapa Hortaliças-Artig. Periódico Indexado (ALICE) 2004, 4, 350–355. [Google Scholar] [CrossRef][Green Version]
- Mimura, Y.; Yoshikawa, M.; Hirai, M. Pepper accession LS2341 is highly resistant to Ralstonia solanacearum strains from Japan. HortScience 2009, 44, 2038–2040. [Google Scholar] [CrossRef]
- Kim, B.; Cheung, J.; Cha, Y.; Hwang, H. Resistance to bacterial wilt of introduced peppers. Korean J. Plant Pathol. 1998, 14, 217–219. [Google Scholar]
- Tung, P.X.; Rasco, E.T.; Vander Zaag, P.; Schmiediche, P. Resistance to Pseudomonas solanacearum in the potato: I. Effects of sources of resistance and adaptation. Euphytica 1990, 45, 203–210. [Google Scholar] [CrossRef]
- Lafortune, D.; Béramis, M.; Daubèze, A.-M.; Boissot, N.; Palloix, A. Partial resistance of pepper to bacterial wilt is oligogenic and stable under tropical conditions. Plant Dis. 2005, 89, 501–506. [Google Scholar] [CrossRef]
- Mimura, Y.; Kageyama, T.; Minamiyama, Y.; Hirai, M. QTL analysis for resistance to Ralstonia solanacearum in Capsicum accession ‘LS2341’. J. Jpn. Soc. Hortic. Sci. 2009, 78, 307–313. [Google Scholar] [CrossRef]
- Du, H.; Wen, C.; Zhang, X.; Xu, X.; Yang, J.; Chen, B.; Geng, S. Identification of a major QTL (qRRs-10.1) that confers resistance to Ralstonia solanacearum in pepper (Capsicum annuum) using SLAF-BSA and QTL mapping. Int. J. Mol. Sci. 2019, 20, 5887. [Google Scholar] [CrossRef]
- Sonah, H.; Bastien, M.; Iquira, E.; Tardivel, A.; Légaré, G.; Boyle, B.; Normandeau, É.; Laroche, J.; Larose, S.; Jean, M. An improved genotyping by sequencing (GBS) approach offering increased versatility and efficiency of SNP discovery and genotyping. PLoS ONE 2013, 8, e54603. [Google Scholar] [CrossRef]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S. QTL-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef]
- Varshney, R.K.; Nayak, S.N.; May, G.D.; Jackson, S.A. Next-generation sequencing technologies and their implications for crop genetics and breeding. Trends Biotecnol. 2009, 27, 522–530. [Google Scholar] [CrossRef]
- Hayward, A.; Mason, A.; Dalton-Morgan, J.; Zander, M.; Edwards, D.; Batley, J. SNP discovery and applications in Brassica napus. J. Plant Biotechnol. 2012, 39, 49–61. [Google Scholar] [CrossRef]
- Elshire, R.; Glaubitz, J.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Baldwin, S.; Pither-Joyce, M.; Wright, K.; Chen, L.; McCallum, J. Development of robust genomic simple sequence repeat markers for estimation of genetic diversity within and among bulb onion (Allium cepa L.) populations. Mol. Breed. 2012, 30, 1401–1411. [Google Scholar] [CrossRef]
- Davey, J.W.; Hohenlohe, P.A.; Etter, P.D.; Boone, J.Q.; Catchen, J.M.; Blaxter, M.L. Genome-wide genetic marker discovery and genotyping using next-generation sequencing. Nat. Rev. Genet. 2011, 12, 499–510. [Google Scholar] [CrossRef]
- Romay, M.C.; Millard, M.J.; Glaubitz, J.C.; Peiffer, J.A.; Swarts, K.L.; Casstevens, T.M.; Elshire, R.J.; Acharya, C.B.; Mitchell, S.E.; Flint-Garcia, S.A. Comprehensive genotyping of the USA national maize inbred seed bank. Genome Biol. 2013, 14, R55. [Google Scholar] [CrossRef]
- Li, H.; Vikram, P.; Singh, R.P.; Kilian, A.; Carling, J.; Song, J.; Burgueno-Ferreira, J.A.; Bhavani, S.; Huerta-Espino, J.; Payne, T. A high density GBS map of bread wheat and its application for dissecting complex disease resistance traits. BMC Genom. 2015, 16, 216. [Google Scholar] [CrossRef]
- Iquira, E.; Humira, S.; François, B. Association mapping of QTLs for sclerotinia stem rot resistance in a collection of soybean plant introductions using a genotyping by sequencing (GBS) approach. BMC Plant Biol. 2015, 15, 5. [Google Scholar] [CrossRef]
- Jaganathan, D.; Thudi, M.; Kale, S.; Azam, S.; Roorkiwal, M.; Gaur, P.M.; Kishor, P.K.; Nguyen, H.; Sutton, T.; Varshney, R.K. Genotyping-by-sequencing based intra-specific genetic map refines a ‘‘QTL-hotspot” region for drought tolerance in chickpea. Mol. Genet. Genom. 2015, 290, 559–571. [Google Scholar] [CrossRef]
- Siddique, M.I.; Lee, H.-Y.; Ro, N.-Y.; Han, K.; Venkatesh, J.; Solomon, A.M.; Patil, A.S.; Changkwian, A.; Kwon, J.-K.; Kang, B.-C. Identifying candidate genes for Phytophthora capsici resistance in pepper (Capsicum annuum) via genotyping-by-sequencing-based QTL mapping and genome-wide association study. Sci. Rep. 2019, 9, 9962. [Google Scholar] [CrossRef]
- Kang, Y.-J.; Lee, B.-M.; Nam, M.; Oh, K.-W.; Lee, M.-H.; Kim, T.-H.; Jo, S.-H.; Lee, J.-H. Identification of quantitative trait loci associated with flowering time in perilla using genotyping-by-sequencing. Mol. Biol. Rep. 2019, 46, 4397–4407. [Google Scholar] [CrossRef]
- Jo, J.; Purushotham, P.M.; Han, K.; Lee, H.-R.; Nah, G.; Kang, B.-C. Development of a genetic map for onion (Allium cepa L.) using reference-free genotyping-by-sequencing and SNP assays. Front. Plant Sci. 2017, 8, 1606. [Google Scholar] [CrossRef]
- Reyes, V.P.; Angeles-Shim, R.B.; Mendioro, M.S.; Manuel, M.; Carmina, C.; Lapis, R.S.; Shim, J.; Sunohara, H.; Nishiuchi, S.; Kikuta, M. Marker-Assisted Introgression and Stacking of Major QTLs Controlling Grain Number (Gn1a) and Number of Primary Branching (WFP) to NERICA Cultivars. Plants 2021, 10, 844. [Google Scholar] [CrossRef]
- Kitony, J.K.; Sunohara, H.; Tasaki, M.; Mori, J.-I.; Shimazu, A.; Reyes, V.P.; Yasui, H.; Yamagata, Y.; Yoshimura, A.; Yamasaki, M. Development of an Aus-Derived Nested Association Mapping (Aus-NAM) Population in Rice. Plants 2021, 10, 1255. [Google Scholar] [CrossRef]
- Rajametov, S.N.; Lee, K.; Jeong, H.B.; Cho, M.C.; Nam, C.W.; Yang, E.Y. Physiological Traits of Thirty-Five Tomato Accessions (Solanum lycopersicum L.) in Response to Low Temperature. Agriculture 2021, 11, 792. [Google Scholar] [CrossRef]
- Allen, G.; Flores-Vergara, M.; Krasynanski, S.; Kumar, S.; Thompson, W. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Kim, J.-E.; Oh, S.-K.; Lee, J.-H.; Lee, B.-M.; Jo, S.-H. Genome-wide SNP calling using next generation sequencing data in tomato. Mol. Cells 2014, 37, 36. [Google Scholar] [CrossRef]
- Eun, M.H.; Han, J.-H.; Yoon, J.B.; Lee, J. QTL mapping of resistance to the Cucumber mosaic virus P1 strain in pepper using a genotyping-by-sequencing analysis. Hortic. Environ. Biotechnol. 2016, 57, 589–597. [Google Scholar] [CrossRef]
- Kosambi, D. The estimation of map distance. Ann. Eugen. 1944, 12, 505–525. [Google Scholar]
- Voorrips, R. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Wang, S.; Basten, C.; Zeng, Z. Windows QTL Cartographer; Bioinformatics Research Center, North Carolina State University: Raleigh, NC, USA, 2007. [Google Scholar]
- Tran, N.H.; Kim, B.S. Sources of resistance to bacterial wilt found in Vietnam collections of pepper (Capsicum annuum) and their nuclear fertility restorer genotypes for cytoplasmic male sterility. Plant. Pathol. J. 2012, 28, 418–422. [Google Scholar] [CrossRef]
- Deschamps, S.; Llaca, V.; May, G.D. Genotyping-by-sequencing in plants. Biology 2012, 1, 460–483. [Google Scholar] [CrossRef]
- Poland, J.A.; Rife, T.W. Genotyping-by-sequencing for plant breeding and genetics. Plant Genome 2012, 5. [Google Scholar] [CrossRef]
- Bielenberg, D.G.; Rauh, B.; Fan, S.; Gasic, K.; Abbott, A.G.; Reighard, G.L.; Okie, W.R.; Wells, C.E. Genotyping by sequencing for SNP-based linkage map construction and QTL analysis of chilling requirement and bloom date in peach [Prunus persica (L.) Batsch]. PLoS ONE 2015, 10, e0139406. [Google Scholar]
- Boutet, G.; Carvalho, S.A.; Falque, M.; Peterlongo, P.; Lhuillier, E.; Bouchez, O.; Lavaud, C.; Pilet-Nayel, M.-L.; Rivière, N.; Baranger, A. SNP discovery and genetic mapping using genotyping by sequencing of whole genome genomic DNA from a pea RIL population. BMC Genom. 2016, 17, 121. [Google Scholar] [CrossRef]
- Iqbal, M.M.; Huynh, M.; Udall, J.A.; Kilian, A.; Adhikari, K.N.; Berger, J.D.; Erskine, W.; Nelson, M.N. The first genetic map for yellow lupin enables genetic dissection of adaptation traits in an orphan grain legume crop. BMC Genet. 2019, 20, 68. [Google Scholar] [CrossRef]
- Manivannan, A.; Choi, S.; Jun, T.-H.; Yang, E.-Y.; Kim, J.-H.; Lee, E.-S.; Lee, H.-E.; Kim, D.-S.; Ahn, Y.-K. Genotyping by Sequencing-Based Discovery of SNP Markers and Construction of Linkage Map from F5 Population of Pepper with Contrasting Powdery Mildew Resistance Trait. BioMed Res. Int. 2021, 2021, 6673010. [Google Scholar] [CrossRef]
- Thakur, P.P.; Mathew, D.; Nazeem, P.; Abida, P.; Indira, P.; Girija, D.; Shylaja, M.; Valsala, P. Identification of allele specific AFLP markers linked with bacterial wilt [Ralstonia solanacearum (Smith) Yabuuchi et al.] resistance in hot peppers (Capsicum annuum L.). Physiol. Mol. Plant Pathol. 2014, 87, 19–24. [Google Scholar] [CrossRef]
- Kwon, J.-S.; Nam, J.-Y.; Yeom, S.-I.; Kang, W.-H. Leaf-to-whole plant spread bioassay for pepper and Ralstonia solanacearum interaction determines inheritance of resistance to bacterial wilt for further breeding. Int. J. Mol. Sci. 2021, 22, 2279. [Google Scholar] [CrossRef]
- Mimura, Y.; Inoue, T.; Minamiyama, Y.; Kubo, N. An SSR-based genetic map of pepper (Capsicum annuum L.) serves as an anchor for the alignment of major pepper maps. Breed. Sci. 2012, 62, 93–98. [Google Scholar] [CrossRef]
- Mathew, D. Analysis of QTL Bw1 and marker CAMS451 associated with the bacterial wilt resistance in hot pepper (Capsicum annuum L.). Plant Gene 2020, 24, 100260. [Google Scholar] [CrossRef]
- Kim, S.B.; Kang, W.H.; Huy, H.N.; Yeom, S.I.; An, J.T.; Kim, S.; Kang, M.Y.; Kim, H.J.; Jo, Y.D.; Ha, Y. Divergent evolution of multiple virus-resistance genes from a progenitor in Capsicum spp. New Phytol. 2017, 213, 886–899. [Google Scholar] [CrossRef]
- Tai, T.H.; Dahlbeck, D.; Clark, E.T.; Gajiwala, P.; Pasion, R.; Whalen, M.C.; Stall, R.E.; Staskawicz, B.J. Expression of the Bs2 pepper gene confers resistance to bacterial spot disease in tomato. Proc. Natl. Acad. Sci. USA 1999, 96, 14153–14158. [Google Scholar] [CrossRef]
- Kearney, B.; Staskawicz, B.J. Widespread distribution and fitness contribution of Xanthomonas campestris avirulence gene avrBs2. Nature 1990, 346, 385–386. [Google Scholar] [CrossRef]
- Kunwar, S.; Iriarte, F.; Fan, Q.; Evaristo da Silva, E.; Ritchie, L.; Nguyen, N.S.; Freeman, J.H.; Stall, R.E.; Jones, J.B.; Minsavage, G.V. Transgenic expression of EFR and Bs2 genes for field management of bacterial wilt and bacterial spot of tomato. Phytopathology 2018, 108, 1402–1411. [Google Scholar] [CrossRef]
- Boutrot, F.; Zipfel, C. Function, discovery, and exploitation of plant pattern recognition receptors for broad-spectrum disease resistance. Annu. Rev. Phytopathol. 2017, 55, 257–286. [Google Scholar] [CrossRef]
- Grant, M.R.; Godiard, L.; Straube, E.; Ashfield, T.; Lewald, J.; Sattler, A.; Innes, R.W.; Dangl, J.L. Structure of the Arabidopsis RPM1 gene enabling dual specificity disease resistance. Science 1995, 269, 843–846. [Google Scholar] [CrossRef]
- Boyes, D.C.; Nam, J.; Dangl, J.L. The Arabidopsis thaliana RPM1 disease resistance gene product is a peripheral plasma membrane protein that is degraded coincident with the hypersensitive response. Proc. Natl. Acad. Sci. USA 1998, 95, 15849–15854. [Google Scholar] [CrossRef]
- Al-Daoude, A.; de Torres Zabala, M.; Ko, J.-H.; Grant, M. RIN13 is a positive regulator of the plant disease resistance protein RPM1. Plant Cell 2005, 17, 1016–1028. [Google Scholar] [CrossRef]
- Van Ooijen, G.; Mayr, G.; Kasiem, M.M.; Albrecht, M.; Cornelissen, B.J.; Takken, F.L. Structure–function analysis of the NB-ARC domain of plant disease resistance proteins. J. Exp. Bot. 2008, 59, 1383–1397. [Google Scholar] [CrossRef]
- Qi, D.; Innes, R.W. Recent advances in plant NLR structure, function, localization, and signaling. Front. Immunol. 2013, 4, 348. [Google Scholar] [CrossRef]
- de Araújo, A.C.; Fonseca, F.C.D.A.; Cotta, M.G.; Alves, G.S.C.; Miller, R.N.G. Plant NLR receptor proteins and their potential in the development of durable genetic resistance to biotic stresses. Biotechnol. Res. Innov. 2019, 3, 80–94. [Google Scholar] [CrossRef]
- Singh, N.K.; Kumar, K.R.R.; Kumar, D.; Shukla, P.; Kirti, P. Characterization of a pathogen induced thaumatin-like protein gene AdTLP from Arachis diogoi, a wild peanut. PLoS ONE 2013, 8, e83963. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, F.; Liang, F.; Zhang, Y.; Ma, L.; Wang, H.; Liu, D. Functional analysis of a pathogenesis-related thaumatin-like protein gene TaLr35PR5 from wheat induced by leaf rust fungus. BMC Plant Biol. 2018, 18, 76. [Google Scholar] [CrossRef]
- Ali, G.S.; Hu, X.; Reddy, A. Overexpression of the Arabidopsis thaumatin-like protein 1 in transgenic potato plants enhances resistance against early and late blights. BioRxiv 2019, 621649. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | F2 Population (n = 94) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Max | Min | Range | Wilt Rate (%) a | Average b | Standard Error | Variation | Skewness | Kurtosis | |
| Disease index | 4 | 0 | 0–4 | 68.085 | 2.638 | 0.167 | 2.620 | −0.604 | −1.352 |
| Chromosome | Linkage Group | Number of SNP Marker | Genetic Distance (cM) | Correlation Coefficient |
|---|---|---|---|---|
| Chr.01 | LG_01-1 | 123 | 82.134 | 0.755 |
| LG_01-2 | 49 | 38.254 | 0.385 | |
| Chr.02 | LG_02-1 | 60 | 20.799 | 0.096 |
| LG_02-2 | 90 | 70.704 | 0.644 | |
| Chr.03 | LG_03 | 172 | 59.561 | 0.790 |
| Chr.04 | LG_04 | 117 | 78.045 | 0.867 |
| Chr.05 | LG_05-1 | 91 | 47.582 | 0.840 |
| LG_05-2 | 12 | 15.952 | 0.437 | |
| Chr.06 | LG_06 | 146 | 52.635 | 0.867 |
| Chr.07 | LG_07 | 136 | 61.809 | 0.876 |
| Chr.08 | LG_08 | 54 | 47.681 | 0.513 |
| Chr.09 | LG_09 | 114 | 74.982 | 0.786 |
| Chr.10 | LG_10-1 | 11 | 19.247 | 0.603 |
| LG_10-2 | 139 | 54.224 | 0.857 | |
| Chr.11 | LG_11 | 123 | 59.579 | 0.867 |
| Chr.12 | LG_12 | 113 | 45.261 | 0.541 |
| Total | 16 | 1550 | 828.449 | - |
| Average | - | 96.875 | 51.778 | 0.670 |
| Chr. | Marker (Chr_pos) | QTL Position (cM) | LOD | Additive | Dominant | a R2 (%) | b |d/a| | c |d/a| Value | d Number of Gene |
|---|---|---|---|---|---|---|---|---|---|
| Chr. 01-1 | ch01_47793963 | 51.73 | 7.065493 | −0.85811 | 0.701208 | 23.9114 | D | 0.82 | 1 |
| ch01_47793907 | 52.21 | 7.323219 | −0.88307 | 0.690647 | 25.1692 | PD | 0.78 | 1 | |
| ch01_52128774 | 52.94 | 6.962209 | −0.88383 | 0.603339 | 24.0817 | PD | 0.68 | 5 | |
| ch01_57574786 | 54.32 | 6.440147 | −0.85645 | 0.651621 | 23.1222 | PD | 0.76 | 0 | |
| ch01_61496208 | 56.25 | 6.180448 | −0.89693 | 0.496732 | 24.5181 | PD | 0.55 | 2 | |
| ch01_58075358 | 57.31 | 5.682495 | −0.79976 | 0.700647 | 20.9856 | D | 0.88 | 0 | |
| ch01_56960124 | 57.65 | 5.983697 | −0.93776 | 0.399064 | 23.0298 | PD | 0.43 | 0 | |
| ch01_156505108 | 58.98 | 6.161348 | −0.88705 | 0.631756 | 24.0344 | PD | 0.71 | 11 | |
| ch01_130789593 | 59.46 | 5.602784 | −0.81563 | 0.56539 | 20.171 | PD | 0.69 | 0 | |
| ch01_160219891 | 60.27 | 5.506165 | −0.95201 | 0.162341 | 22.5886 | A | 0.17 | 2 | |
| ch01_130913560 | 60.36 | 6.216155 | −0.92824 | 0.283328 | 22.2838 | PD | 0.31 | 0 | |
| ch01_161127926 | 61.1 | 5.692397 | −0.85691 | 0.394691 | 20.1396 | PD | 0.46 | 10 |
| Gene Name | Gene Start | Gene End | CM334(v.1.55) | KEGG/GO | SwissProt | NR |
|---|---|---|---|---|---|---|
| CA01g12080 | 47862177 | 47863580 | Putative amino acid transporter | - | Putative polyamine transporter | Putative polyamine transporter |
| CA01g12450 | 52035941 | 52041393 | Calcium-dependent protein kinase 17 | K13412 (calcium-dependent protein kinase) | Calcium-dependent protein kinase 17 | Calcium-dependent protein kinase 17-like |
| CA01g12460 | 52113432 | 52115689 | Rho GTPase-activating protein | - | Rho GTPase-activating protein 7 | rho GTPase-activating protein 7 |
| CA01g12470 | 52119767 | 52121954 | Putative rho GTPase activator | - | Rho GTPase-activating protein 7 | PREDICTED: rho GTPase-activating protein 7-like |
| CA01g12480 | 52128303 | 52129214 | Rho GTPase-activating protein 7-like isoform X2 (Solanum tuberosum) | - | Rho GTPase-activating protein 7 | PREDICTED: rho GTPase-activating protein 7-like |
| CA01g12490 | 52147172 | 52148002 | Phospholipase A22 | - | Uncharacterized protein | Putative rho GTPase-activating protein 7-like isoform X1 |
| CA01g13370 | 61438907 | 61440480 | Arsenite-resistance protein, putative | - | Serrate RNA effector molecule | Serrate RNA effector molecule |
| CA01g13380 | 61446486 | 61447578 | Arsenite-resistance protein, putative | - | Serrate RNA effector molecule | Serrate RNA effector molecule |
| CA01g18610 | 156433050 | 156436475 | Detected protein of unknown function | - | O-fucosyltransferase family protein | Putative siroheme synthase-like |
| CA01g18620 | 156438428 | 156442076 | AT5g40850/MHK7_8 | K02303 (uroporphyrin-III C-methyltransferase) | Siroheme synthase | PREDICTED: siroheme synthase |
| CA01g18630 | 156454093 | 156456205 | Putative glutathione S-transferase T1 | K00799 (glutathione S-transferase) | Putative glutathione S-transferase | PREDICTED: probable glutathione S-transferase |
| CA01g18640 | 156457355 | 156460628 | Cohesin subunit rad21, putative | K06670 (RAD21; cohesin complex subunit SCC1) | Uncharacterized protein | PREDICTED: sister chromatid cohesion 1 protein 3 |
| CA01g18650 | 156487695 | 156490046 | Anthranilate phosphoribosyltransferase-like protein | - | Uncharacterized protein | Putative disease-resistance protein RPM1-like |
| CA01g18660 | 156497587 | 156514448 | DNA-directed RNA polymerase | - | DNA-directed RNA polymerase subunit beta | PREDICTED: DNA-directed RNA polymerase I subunit 2 |
| CA01g18670 | 156515197 | 156518484 | Putative synaptotagmin | - | Protein QUIRKY | Protein QUIRKY |
| CA01g18680 | 156539078 | 156540145 | Detected protein of unknown function | K22733 (NIPA, SLC57A2S, magnesium transporter) | Magnesium transporter | Putative magnesium transporter NIPA8 |
| CA01g18690 | 156550800 | 156555986 | DnaJ-like protein | K03686 (DnaJ, molecular chaperone DnaJ) | Uncharacterized protein | Hypothetical protein T459_02496 |
| CA01g18700 | 156563245 | 156580138 | Eukaryotic translation initiation factor 3 subunit, putative | K03255 (TIF31, CLU1, protein TIF31) | Clustered mitochondria protein | Clustered mitochondria protein |
| CA01g18710 | 156602485 | 156602895 | Detected protein of unknown function | K11253 (Histone H3) | Histone H3 | PREDICTED: histone H3.2-like |
| CA01g19920 | 160217017 | 160229898 | Ankyrin repeat family protein (Populus trichocarpa) | K24810 (IBTK, inhibitor of Bruton tyrosine kinase) | Ankyrin repeat domain-containing protein | Hypothetical protein FXO37_07210 |
| CA01g19930 | 160315849 | 160316316 | Polypeptide with a gag-like domain | - | LRRNT-2 domain-containing protein | Uncharacterized protein |
| CA01g20080 | 161050160 | 161051870 | Transcription factor bHLH130-like isoform X1 (Citrus sinensis) | - | Transcription factor bHLH130 | PREDICTED: transcription factor bHLH130-like |
| CA01g20090 | 161051871 | 161052295 | DNA binding protein, putative | - | Transcription factor bHLH130 | PREDICTED: transcription factor bHLH130-like |
| CA01g20100 | 161113518 | 161117641 | Ras-related protein RGP1-like (Solanum tuberosum) | K07904 (RAB11A, Ras-related protein Rab-11A) | Ras-related protein RABA4c | Ras-related protein RABA4c |
| CA01g20110 | 161118723 | 161120113 | Thaumatin-like protein SE39b | -/GO (0006952): defense response | Uncharacterized protein | PREDICTED: thaumatin-like protein 1b isoform X1 |
| CA01g20120 | 161154079 | 161154489 | Histone H3 family protein (Populus trichocarpa) | K11253 (H3, histone H3) | Histone H3 | PREDICTED: histone H3.2-like |
| CA01g20130 | 161161494 | 161162582 | Disease resistance protein RPM1-like (Fragaria vesca) subsp. vesca | K13457 (RPM1, RPS3; disease-resistance protein RPM1) | Uncharacterized protein | Hypothetical protein T459_01449 |
| CA01g20140 | 161184987 | 161187221 | Detected protein of unknown function | - | NB-ARC domain-containing protein | PREDICTED: putative late blight-resistance protein homolog R1B-23 |
| CA01g20150 | 161212363 | 161212704 | Detected protein of unknown function | - | Uncharacterized protein | Putative cellulose synthase A catalytic subunit 3 (UDP-forming)-like |
| CA01g20160 | 161215973 | 161218180 | Detected protein of unknown function | - | Uncharacterized protein | Uncharacterized protein LOC107839230 isoform X2 |
| CA01g20170 | 161221263 | 161222494 | Detected protein of unknown function | - | Uncharacterized protein | Uncharacterized protein LOC107839213 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chae, S.-Y.; Lee, K.; Do, J.-W.; Hong, S.-C.; Lee, K.-H.; Cho, M.-C.; Yang, E.-Y.; Yoon, J.-B. QTL Mapping of Resistance to Bacterial Wilt in Pepper Plants (Capsicum annuum) Using Genotyping-by-Sequencing (GBS). Horticulturae 2022, 8, 115. https://doi.org/10.3390/horticulturae8020115
Chae S-Y, Lee K, Do J-W, Hong S-C, Lee K-H, Cho M-C, Yang E-Y, Yoon J-B. QTL Mapping of Resistance to Bacterial Wilt in Pepper Plants (Capsicum annuum) Using Genotyping-by-Sequencing (GBS). Horticulturae. 2022; 8(2):115. https://doi.org/10.3390/horticulturae8020115
Chicago/Turabian StyleChae, Soo-Young, Kwanuk Lee, Jae-Wahng Do, Sun-Cheul Hong, Kang-Hyun Lee, Myeong-Cheoul Cho, Eun-Young Yang, and Jae-Bok Yoon. 2022. "QTL Mapping of Resistance to Bacterial Wilt in Pepper Plants (Capsicum annuum) Using Genotyping-by-Sequencing (GBS)" Horticulturae 8, no. 2: 115. https://doi.org/10.3390/horticulturae8020115
APA StyleChae, S.-Y., Lee, K., Do, J.-W., Hong, S.-C., Lee, K.-H., Cho, M.-C., Yang, E.-Y., & Yoon, J.-B. (2022). QTL Mapping of Resistance to Bacterial Wilt in Pepper Plants (Capsicum annuum) Using Genotyping-by-Sequencing (GBS). Horticulturae, 8(2), 115. https://doi.org/10.3390/horticulturae8020115