

Genome-Wide Identification and Expression Profile Analysis of the WUSCHEL-Related Homeobox (WOX) Genes in Woodland Strawberry (Fragaria vesca)

Abstract
1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification of WOX Genes in Fragaria vesca
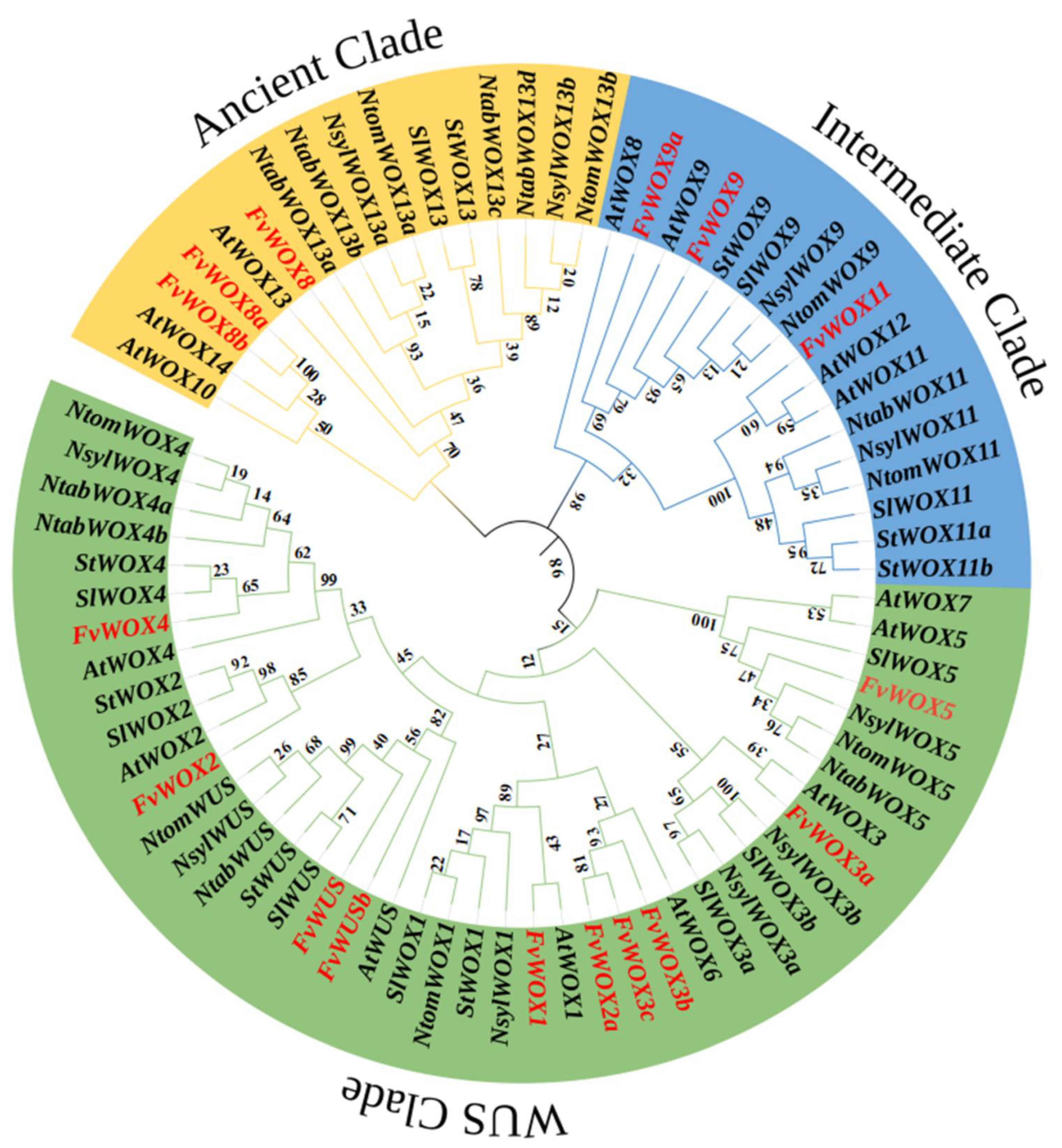
2.2. Protein Properties and Phylogenetic Tree Construction
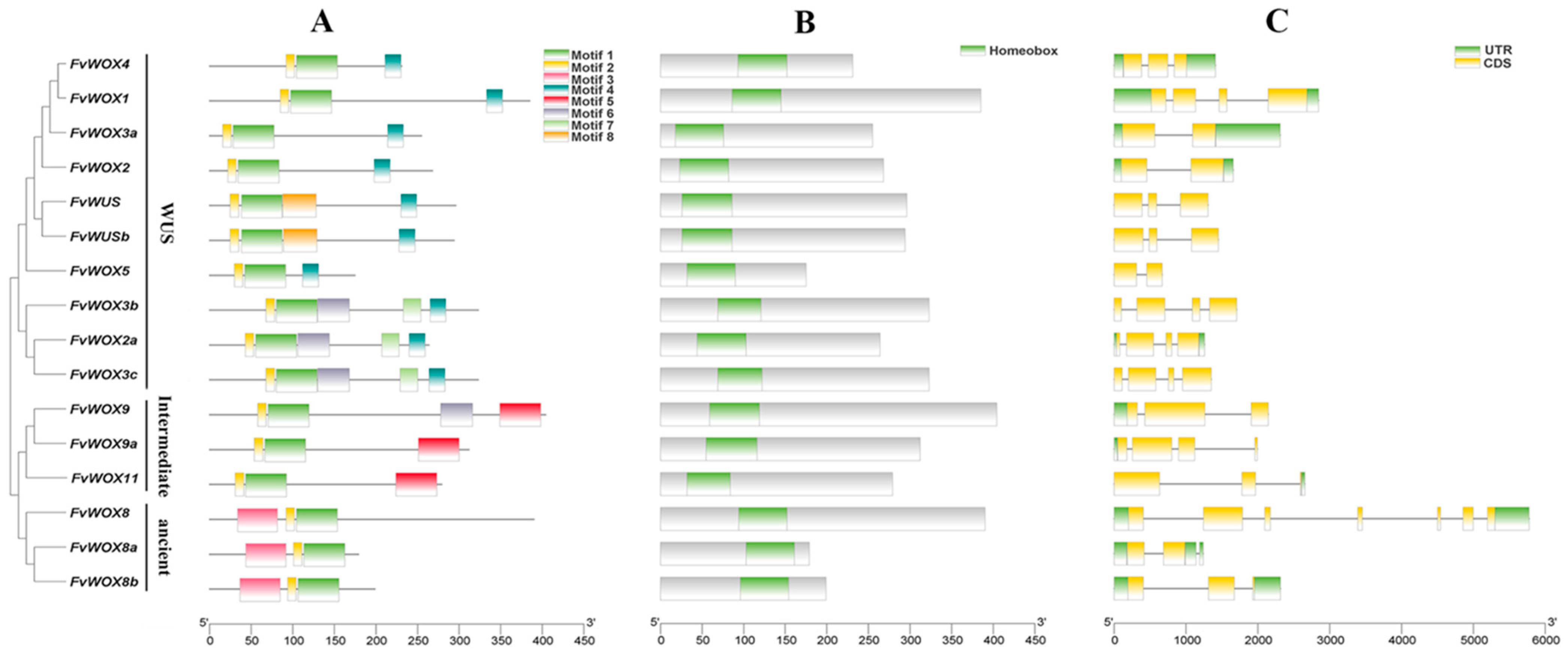
2.3. Gene Structure, Domain and Protein Motif Analysis
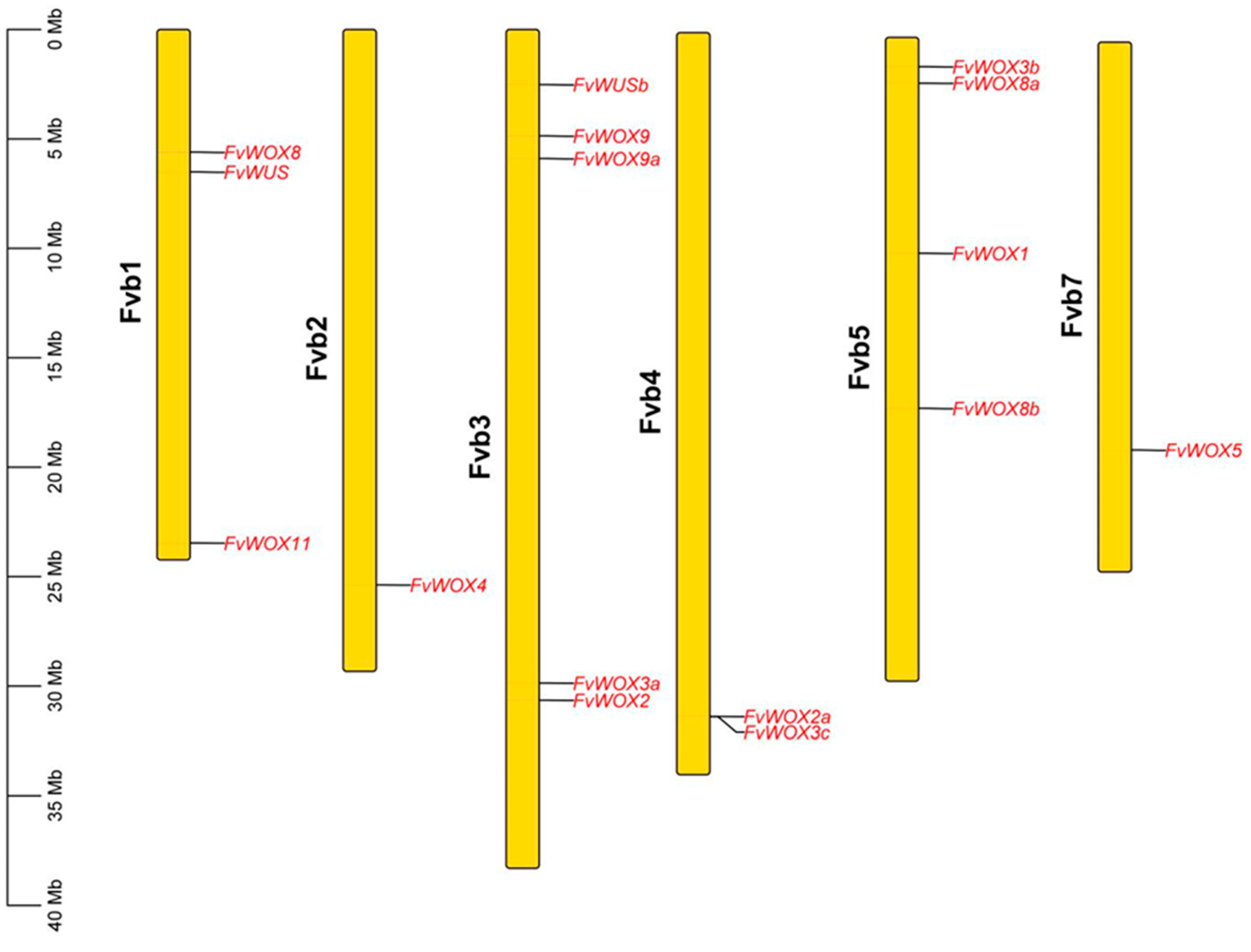
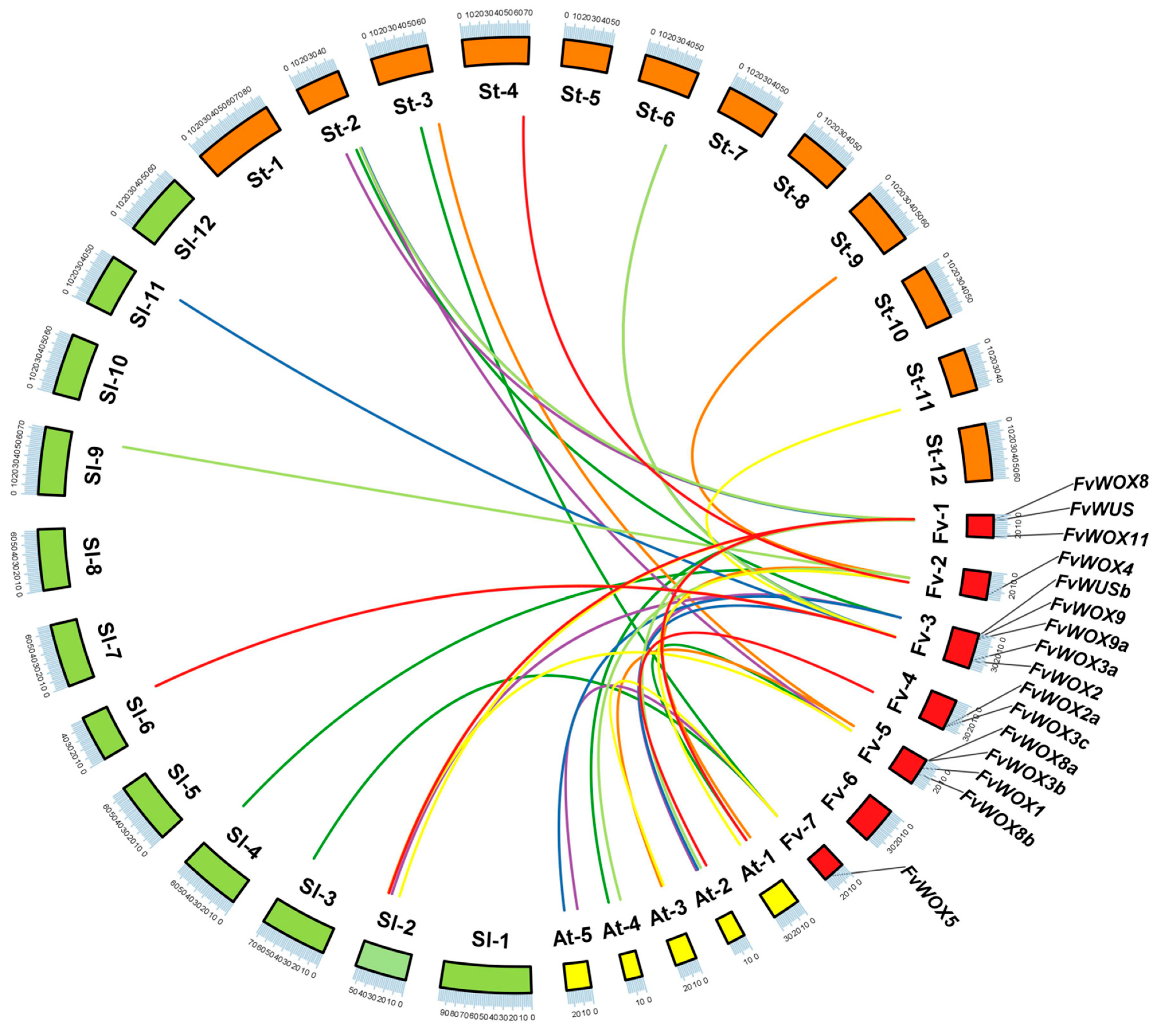
2.4. Chromosome Location and Collinear Analysis of FvWOX Genes
2.5. Cis-Elements Analysis
2.6. Plant Material and Expression Profile Analysis
2.7. Prediction of the Regulatory Networks Represented by FvWOX3a
2.8. Subcellular Localization Analysis
3. Result
3.1. Identification of WOX Genes in Fragaria vesca
3.2. Phylogenetic Analysis and Multiple Sequence Alignment
3.3. Conserved Motif and Gene Structure Analysis
3.4. Collinear Analysis of FvWOX Genes
3.5. Cis-Elements Analysis of FvWOX GENES
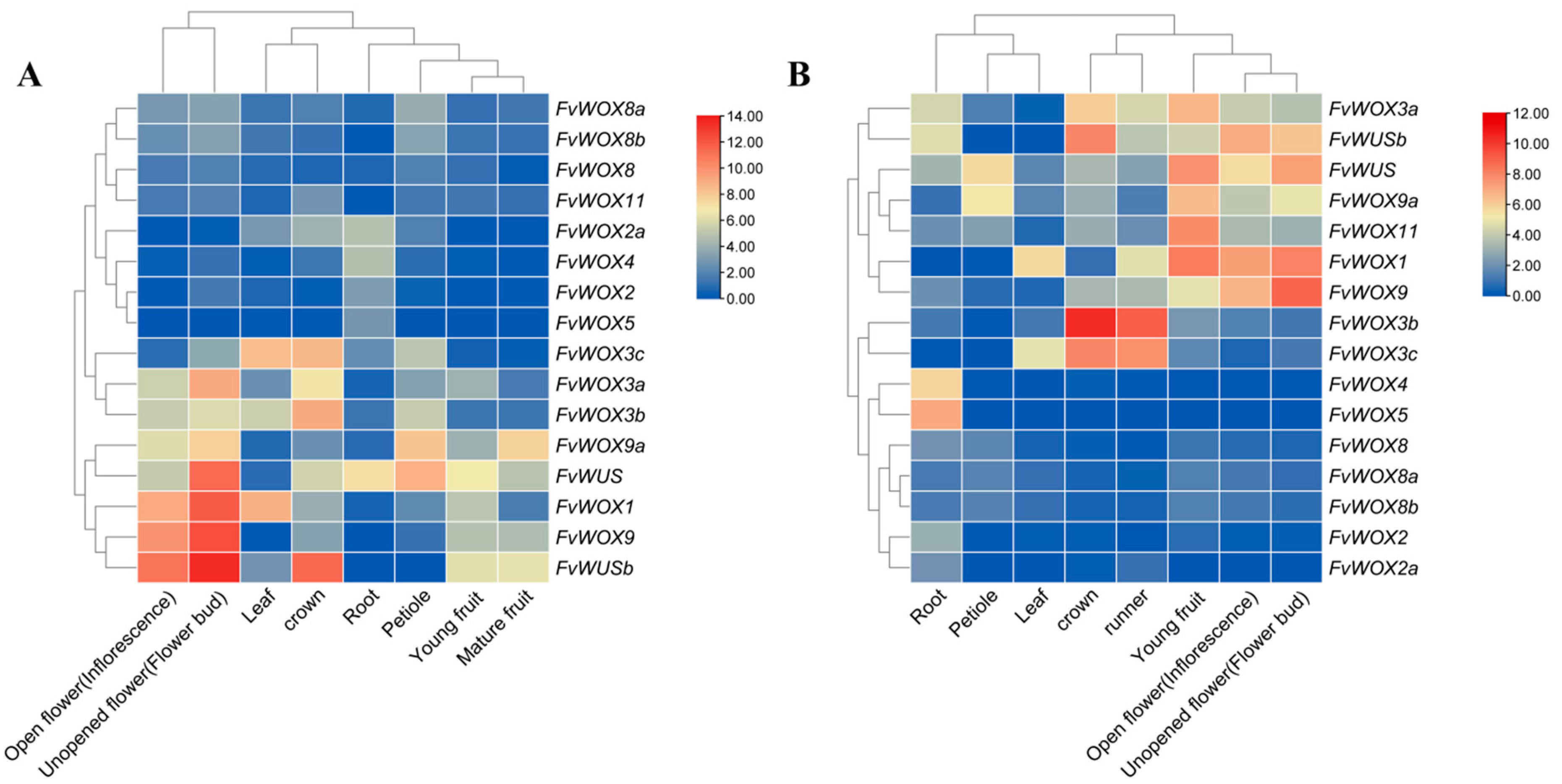
3.6. Expression Patterns of FvWOXs in Different Woodland Strawberry Tissues
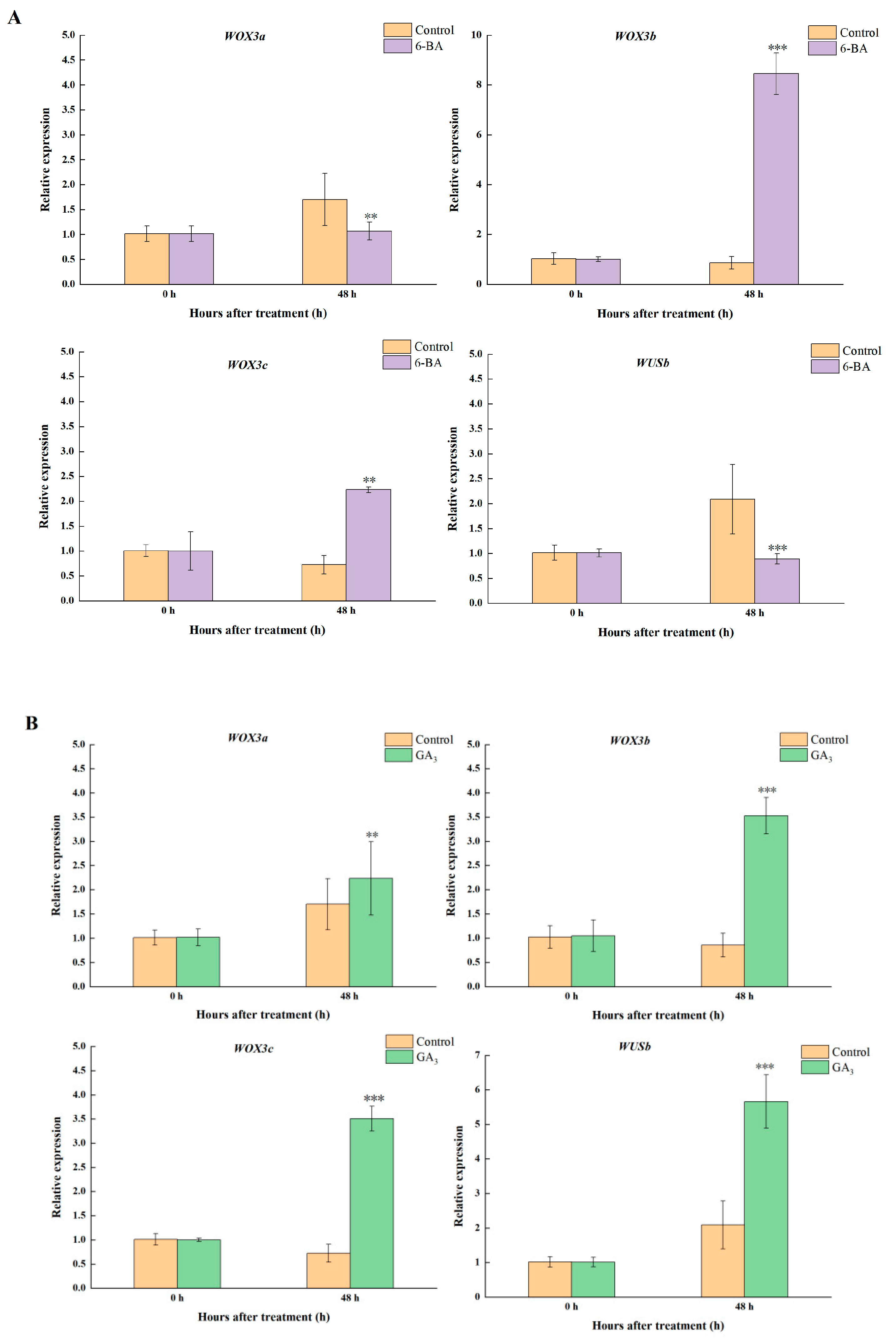
3.7. FvWOXs Highly Expressed in Crown Are Regulated by 6-BA and GA3
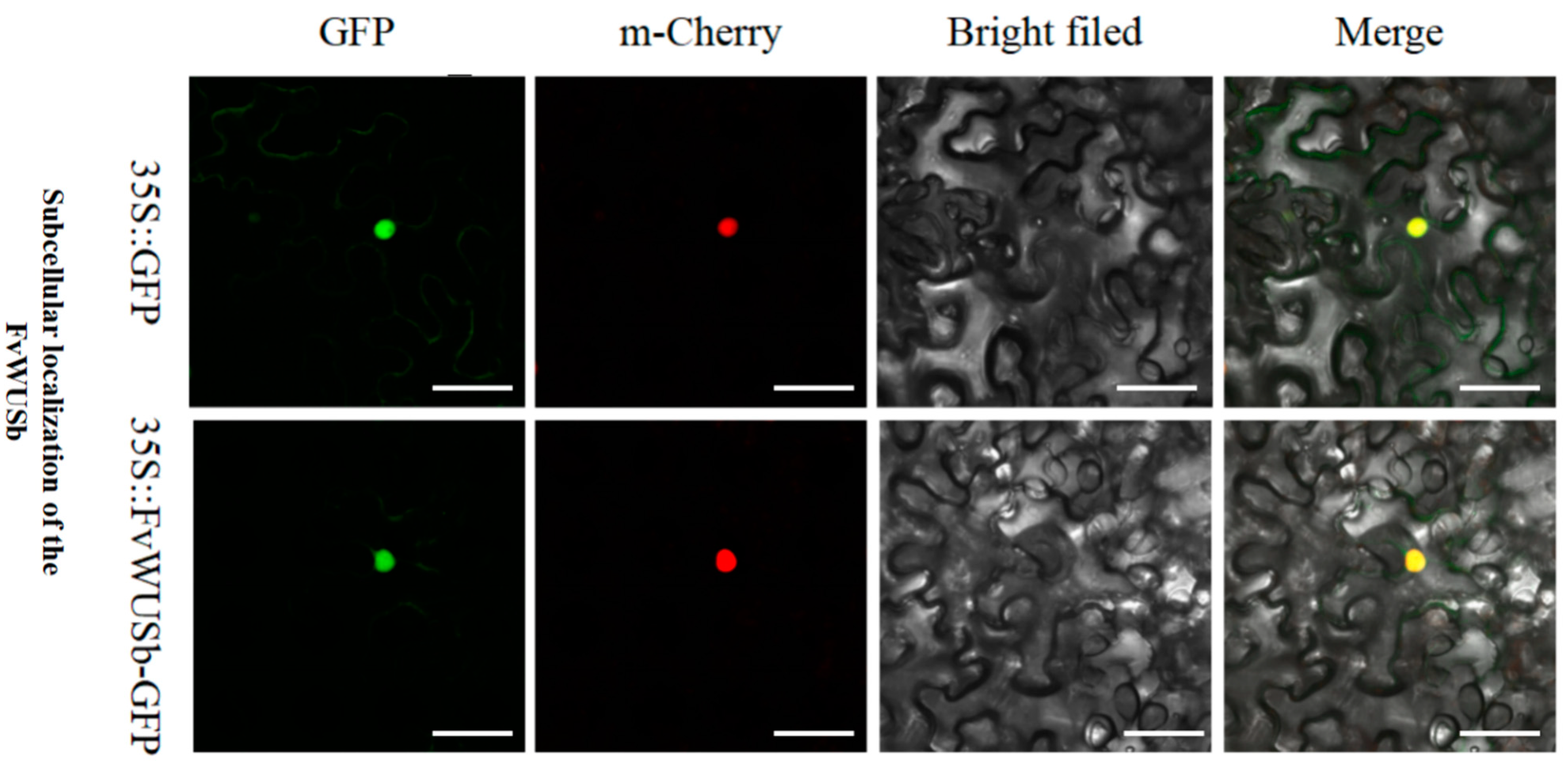
3.8. Subcellular Localization of FvWUSb
3.9. Protein Interaction Network Prediction of FvWOX3a
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jha, P.; Ochatt, S.J.; Kumar, V. WUSCHEL: A master regulator in plant growth signaling. Plant Cell Rep. 2020, 39, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Lian, G.; Ding, Z.; Wang, Q.; Zhang, D.; Xu, J. Origins and evolution of WUSCHEL-related homeobox protein family in plant kingdom. Sci. World J. 2014, 2014, 534140. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, D.; Xia, Y.; Li, Z.; Jing, D.; Du, J.; Niu, N.; Ma, S.; Wang, J.; Song, Y.; et al. Identification of the WUSCHEL-Related Homeobox (WOX) Gene Family, and Interaction and Functional Analysis of TaWOX9 and TaWUS in Wheat. Int. J. Mol. Sci. 2020, 21, 1581. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Li, F.W.; Kramer, E.M. Large-scale phylogenomic analysis suggests three ancient superclades of the WUSCHEL-RELATED HOMEOBOX transcription factor family in plants. PLoS ONE 2019, 14, e0223521. [Google Scholar] [CrossRef] [PubMed]
- Dolzblasz, A.; Nardmann, J.; Clerici, E.; Causier, B.; van der Graaff, E.; Chen, J.; Davies, B.; Werr, W.; Laux, T. Stem Cell Regulation by Arabidopsis WOX Genes. Mol. Plant 2016, 9, 1028–1039. [Google Scholar] [CrossRef]
- He, P.; Zhang, Y.; Liu, H.; Yuan, Y.; Wang, C.; Yu, J.; Xiao, G. Comprehensive analysis of WOX genes uncovers that WOX13 is involved in phytohormone-mediated fiber development in cotton. BMC Plant Biol. 2019, 19, 312. [Google Scholar] [CrossRef]
- Hao, Q.; Zhang, L.; Yang, Y.; Shan, Z.; Zhou, X.A. Genome-Wide Analysis of the WOX Gene Family and Function Exploration of GmWOX18 in Soybean. Plants 2019, 8, 215. [Google Scholar] [CrossRef]
- Mayer, K.F.; Schoof, H.; Haecker, A.; Lenhard, M.; Jürgens, G.; Laux, T. Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell 1998, 95, 805–815. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, R.; Qin, G.; Chen, Z.; Gu, H.; Qu, L.J. Over-expression of WOX1 leads to defects in meristem development and polyamine homeostasis in Arabidopsis. J. Integr. Plant Biol. 2011, 53, 493–506. [Google Scholar] [CrossRef]
- Niu, H.; Liu, X.; Tong, C.; Wang, H.; Li, S.; Lu, L.; Pan, Y.; Zhang, X.; Weng, Y.; Li, Z. The WUSCHEL-related homeobox1 gene of cucumber regulates reproductive organ development. J. Exp. Bot. 2018, 69, 5373–5387. [Google Scholar] [CrossRef]
- Zhu, T.; Moschou, P.N.; Alvarez, J.M.; Sohlberg, J.J.; von Arnold, S. WUSCHEL-RELATED HOMEOBOX 2 is important for protoderm and suspensor development in the gymnosperm Norway spruce. BMC Plant Biol. 2016, 16, 19. [Google Scholar] [CrossRef]
- Shimizu, R.; Ji, J.; Kelsey, E.; Ohtsu, K.; Schnable, P.S.; Scanlon, M.J. Tissue specificity and evolution of meristematic WOX3 function. Plant Physiol. 2009, 149, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Tanaka, S.Y.; Masumoto, Y.; Nobori, N.; Ishii, H.; Hibara, K.; Itoh, J.; Tanisaka, T.; Taketa, S. Barley NARROW LEAFED DWARF1 encoding a WUSCHEL-RELATED HOMEOBOX 3 (WOX3) regulates the marginal development of lateral organs. Breed. Sci. 2016, 66, 416–424. [Google Scholar] [CrossRef][Green Version]
- Suer, S.; Agusti, J.; Sanchez, P.; Schwarz, M.; Greb, T. WOX4 imparts auxin responsiveness to cambium cells in Arabidopsis. Plant Cell 2011, 23, 3247–3259. [Google Scholar] [CrossRef] [PubMed]
- Kucukoglu, M.; Nilsson, J.; Zheng, B.; Chaabouni, S.; Nilsson, O. WUSCHEL-RELATED HOMEOBOX4 (WOX4)-like genes regulate cambial cell division activity and secondary growth in Populus trees. New Phytol. 2017, 215, 642–657. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Moya, F.; Escudero, N.; Zavala-Gonzalez, E.A.; Esteve-Bruna, D.; Blázquez, M.A.; Alabadí, D.; Lopez-Llorca, L.V. Induction of auxin biosynthesis and WOX5 repression mediate changes in root development in Arabidopsis exposed to chitosan. Sci. Rep. 2017, 7, 16813. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Hao, Y.; Cui, H. The WUSCHEL Related Homeobox Protein WOX7 Regulates the Sugar Response of Lateral Root Development in Arabidopsis thaliana. Mol. Plant 2016, 9, 261–270. [Google Scholar] [CrossRef]
- Wolabu, T.W.; Wang, H.; Tadesse, D.; Zhang, F.; Behzadirad, M.; Tvorogova, V.E.; Abdelmageed, H.; Liu, Y.; Chen, N.; Chen, J.; et al. WOX9 functions antagonistic to STF and LAM1 to regulate leaf blade expansion in Medicago truncatula and Nicotiana sylvestris. New Phytol. 2021, 229, 1582–1597. [Google Scholar] [CrossRef]
- Cheng, S.; Zhou, D.X.; Zhao, Y. WUSCHEL-related homeobox gene WOX11 increases rice drought resistance by controlling root hair formation and root system development. Plant Signal. Behav. 2016, 11, e1130198. [Google Scholar] [CrossRef]
- Wang, L.Q.; Wen, S.S.; Wang, R.; Wang, C.; Gao, B.; Lu, M.Z. PagWOX11/12a activates PagCYP736A12 gene that facilitates salt tolerance in poplar. Plant Biotechnol. J. 2021, 19, 2249–2260. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Iwase, A.; Ito, T.; Tanaka, H.; Favero, D.S.; Kawamura, A.; Sakamoto, S.; Wakazaki, M.; Tameshige, T.; Fujii, H.; et al. Wound-inducible WUSCHEL-RELATED HOMEOBOX 13 is required for callus growth and organ reconnection. Plant Physiol. 2022, 188, 425–441. [Google Scholar] [CrossRef] [PubMed]
- Xu, L. WUSCHEL: The versatile protein in the shoot apical meristem. Science China. Life Sci. 2021, 64, 177–178. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Feng, J.; Cheng, L.; Dai, C.; Gao, Q.; Liu, Z.; Kang, C. Gene Expression Profiling of the Shoot Meristematic Tissues in Woodland Strawberry Fragaria vesca. Front. Plant Sci. 2019, 10, 1624. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Mistry, J.; Schusterböckler, B.; Griffithsjones, S.; Hollich, V.; Lassmann, T.; Moxon, S.; Marshall, M.; Khanna, A.; Durbin, R. Pfam: Clans, web tools and services. Nucleic Acids Res. 2006, 34, 247–251. [Google Scholar] [CrossRef]
- Upadhyay, R.K.; Mattoo, A.K. Genome-wide identification of tomato (Solanum lycopersicum L.) lipoxygenases coupled with expression profiles during plant development and in response to methyl-jasmonate and wounding. J. Plant Physiol. 2018, 231, 318–328. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L. MEME Suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; He, Y.; Niu, S.; Yan, S.; Zhang, Y. Identification and characterization of the CONSTANS (CO)/CONSTANS-like (COL) genes related to photoperiodic signaling and flowering in tomato. Plant Sci. 2020, 301, 110653. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Tang, R.; Chen, X.; Xu, Z.; Ren, Z.; Wang, L. Genome-wide identification and characterization of WOX genes in Cucumis sativus. Genome 2021, 64, 761–776. [Google Scholar] [CrossRef]
- Wang, M.M.; Liu, M.M.; Ran, F.; Guo, P.C.; Ke, Y.Z.; Wu, Y.W.; Wen, J.; Li, P.F.; Li, J.N.; Du, H. Global Analysis of WOX Transcription Factor Gene Family in Brassica napus Reveals Their Stress- and Hormone-Responsive Patterns. Int. J. Mol. Sci. 2018, 19, 3470. [Google Scholar] [CrossRef]
- Li, M.; Wang, R.; Liu, Z.; Wu, X.; Wang, J. Genome-wide identification and analysis of the WUSCHEL-related homeobox (WOX) gene family in allotetraploid Brassica napus reveals changes in WOX genes during polyploidization. BMC Genom. 2019, 20, 317. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef]
- Laux, T.; Mayer, K.F.; Berger, J.; Jürgens, G. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 1996, 122, 87–96. [Google Scholar] [CrossRef]
- Yadav, R.K.; Perales, M.; Gruel, J.; Girke, T.; Jönsson, H.; Reddy, G.V. WUSCHEL protein movement mediates stem cell homeostasis in the Arabidopsis shoot apex. Genes Dev. 2011, 25, 2025–2030. [Google Scholar] [CrossRef]
- Andrés, J.; Caruana, J.; Liang, J.; Samad, S.; Monfort, A.; Liu, Z.; Hytönen, T.; Koskela, E.A. Woodland strawberry axillary bud fate is dictated by a crosstalk of environmental and endogenous factors. Plant Physiol. 2021, 187, 1221–1234. [Google Scholar] [CrossRef]
- Cao, Y.; Han, Y.; Meng, D.; Li, G.; Li, D.; Abdullah, M.; Jin, Q.; Lin, Y.; Cai, Y. Genome-Wide Analysis Suggests the Relaxed Purifying Selection Affect the Evolution of WOX Genes in Pyrus bretschneideri, Prunus persica, Prunus mume, and Fragaria vesca. Front. Genet. 2017, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.K.; Luijten, M.; Miyashima, S.; Lenhard, M.; Hashimoto, T.; Nakajima, K.; Scheres, B.; Heidstra, R.; Laux, T. Conserved factors regulate signalling in Arabidopsis thaliana shoot and root stem cell organizers. Nature 2007, 446, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Somssich, M.; Je, B.I.; Simon, R.; Jackson, D. CLAVATA-WUSCHEL signaling in the shoot meristem. Development 2016, 143, 3238–3248. [Google Scholar] [CrossRef]
- Zhao, Z.; Andersen, S.U.; Ljung, K.; Dolezal, K.; Miotk, A.; Schultheiss, S.J.; Lohmann, J.U. Hormonal control of the shoot stem-cell niche. Nature 2010, 465, 1089–1092. [Google Scholar] [CrossRef]
- Li, W.; Zhang, J.; Sun, H.; Wang, S.; Chen, K.; Liu, Y.; Li, H.; Ma, Y.; Zhang, Z. FveRGA1, encoding a DELLA protein, negatively regulates runner production in Fragaria vesca. Planta 2018, 247, 941–951. [Google Scholar] [CrossRef]
- Li, Y.; Hu, J.; Wei, H.; Jeong, B.R. A Long-Day Photoperiod and 6-Benzyladenine Promote Runner Formation through Upregulation of Soluble Sugar Content in Strawberry. Int. J. Mol. Sci. 2020, 21, 4917. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Paek, N.C. Regulatory role of the OsWOX3A transcription factor in rice root development. Plant Signal. Behav. 2016, 11, e1184807. [Google Scholar] [CrossRef]
- Hytönen, T.; Elomaa, P.; Moritz, T.; Junttila, O. Gibberellin mediates daylength-controlled differentiation of vegetative meristems in strawberry (Fragaria × ananassa Duch). BMC Plant Biol. 2009, 9, 18. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene Locus ID | Number of Amino Acids | Molecular Weight (MW) | Isoelectric Point (pI) | Instability Index | Aliphatic Index | Hydropathicity (GRAVY) |
|---|---|---|---|---|---|---|---|
| FvWUS | FvH4_1g11910.1 | 891 | 73,066.77 | 5.09 | 60.27 | 26.04 | −0.764 |
| FvWUSb | FvH4_3g04400.1 | 885 | 72,820.74 | 5.09 | 59.21 | 27.68 | −0.808 |
| FvWOX1 | FvH4_5g17270.1 | 1158 | 95,638.53 | 5.01 | 49.94 | 29.88 | −0.936 |
| FvWOX2 | FvH4_3g35730.1 | 807 | 65,343.68 | 5.12 | 41.45 | 26.39 | −0.728 |
| FvWOX2a | FvH4_4g32650.1 | 795 | 65,214.18 | 5.12 | 46.02 | 31.32 | −0.856 |
| FvWOX3a | FvH4_3g34410.1 | 768 | 65,226.52 | 5.08 | 54.45 | 28.13 | −0.947 |
| FvWOX3b | FvH4_5g02150.1 | 972 | 79,499.19 | 5.08 | 54.78 | 32.2 | −0.869 |
| FvWOX3c | FvH4_4g32660.1 | 972 | 79,784.03 | 5.07 | 55.51 | 33.13 | −0.931 |
| FvWOX4 | FvH4_2g34030.1 | 696 | 57,459.4 | 5.11 | 57.53 | 29.31 | −0.958 |
| FvWOX5 | FvH4_7g24100.1 | 528 | 42,815.14 | 5.23 | 38.05 | 28.79 | −0.663 |
| FvWOX8 | FvH4_1g10280.1 | 1173 | 94,627.87 | 5.07 | 36.63 | 30.86 | −0.756 |
| FvWOX8a | FvH4_5g03570.1 | 540 | 43,069.95 | 5.21 | 35.86 | 33.15 | −0.843 |
| FvWOX8b | FvH4_5g25470.1 | 600 | 47,987.51 | 5.19 | 39.52 | 33.67 | −0.86 |
| FvWOX9 | FvH4_3g08310.1 | 1215 | 102,295.82 | 4.98 | 59.31 | 26.17 | −0.929 |
| FvWOX9a | FvH4_3g10130.1 | 939 | 78,480.07 | 5.04 | 58.91 | 24.49 | −0.855 |
| FvWOX11 | FvH4_1g29830.1 | 840 | 69,986.65 | 5.09 | 39.08 | 25.48 | −0.775 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Zhao, X.; Miao, Y.; Wang, D.; Zhang, Z.; Liu, Y. Genome-Wide Identification and Expression Profile Analysis of the WUSCHEL-Related Homeobox (WOX) Genes in Woodland Strawberry (Fragaria vesca). Horticulturae 2022, 8, 1043. https://doi.org/10.3390/horticulturae8111043
Yang X, Zhao X, Miao Y, Wang D, Zhang Z, Liu Y. Genome-Wide Identification and Expression Profile Analysis of the WUSCHEL-Related Homeobox (WOX) Genes in Woodland Strawberry (Fragaria vesca). Horticulturae. 2022; 8(11):1043. https://doi.org/10.3390/horticulturae8111043
Chicago/Turabian StyleYang, Xu, Xinyong Zhao, Yanan Miao, Dongxue Wang, Zhihong Zhang, and Yuexue Liu. 2022. "Genome-Wide Identification and Expression Profile Analysis of the WUSCHEL-Related Homeobox (WOX) Genes in Woodland Strawberry (Fragaria vesca)" Horticulturae 8, no. 11: 1043. https://doi.org/10.3390/horticulturae8111043
APA StyleYang, X., Zhao, X., Miao, Y., Wang, D., Zhang, Z., & Liu, Y. (2022). Genome-Wide Identification and Expression Profile Analysis of the WUSCHEL-Related Homeobox (WOX) Genes in Woodland Strawberry (Fragaria vesca). Horticulturae, 8(11), 1043. https://doi.org/10.3390/horticulturae8111043