
Metabolomics Data Revealed Metabolite Changes during Endocarp Lignification in Kernel-Using Apricot
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sampling
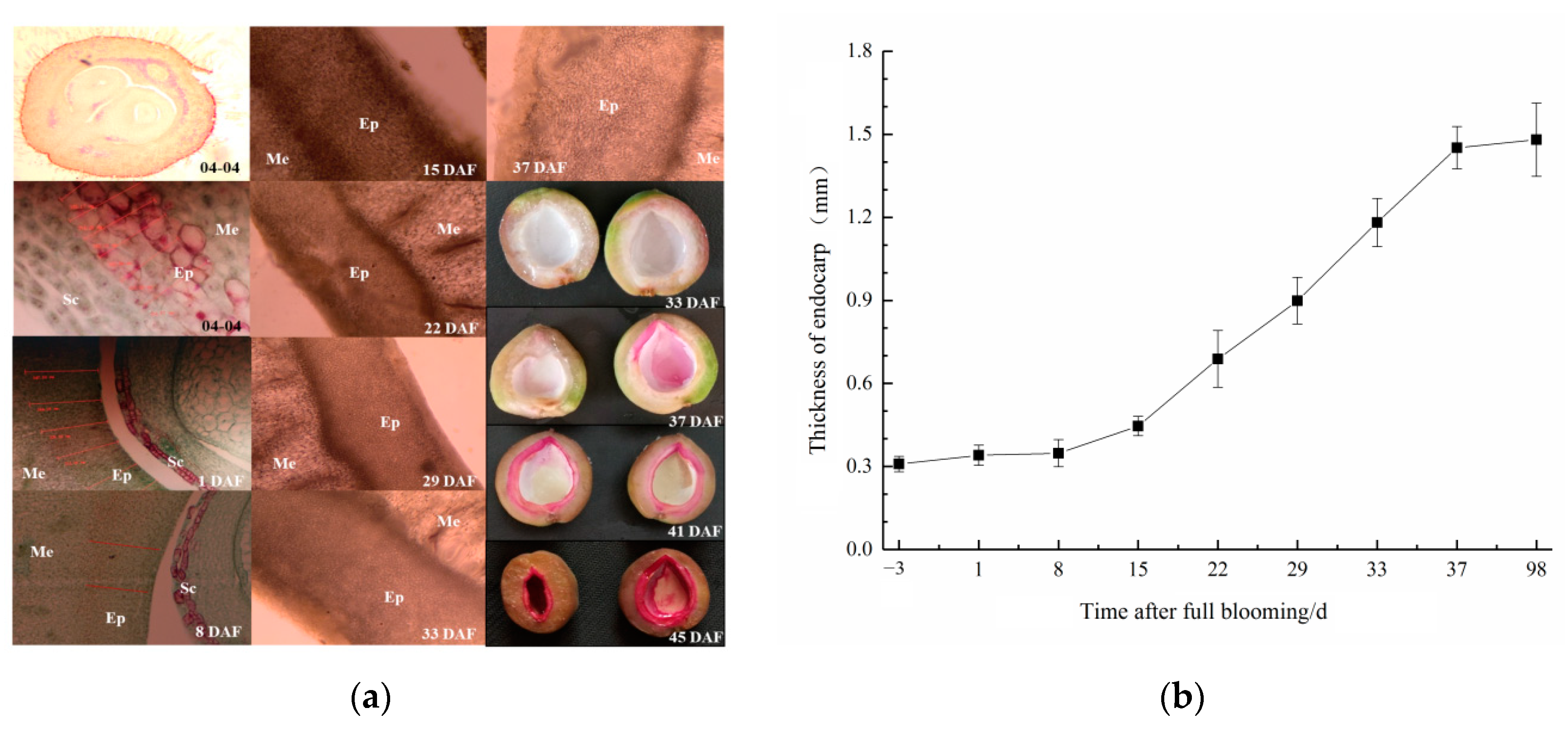
2.2. Observation of Endocarp Development and Lignin Staining
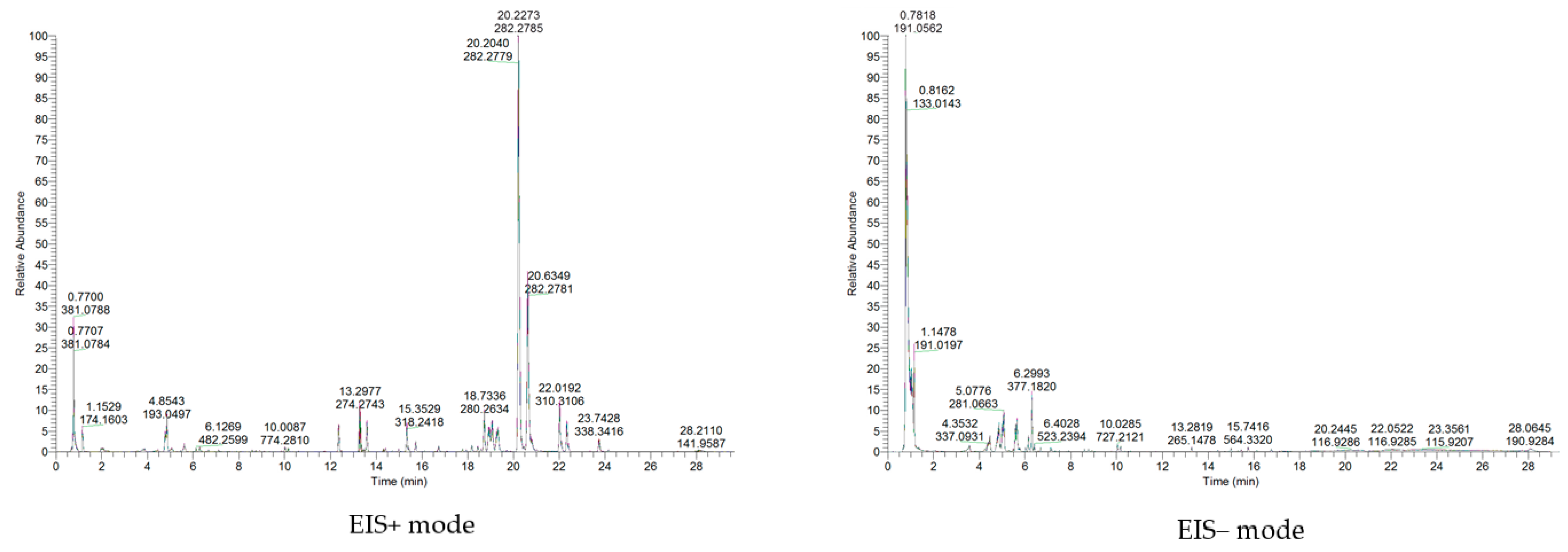
2.3. Sample Preparation and UPLC-MS/MS Analysis
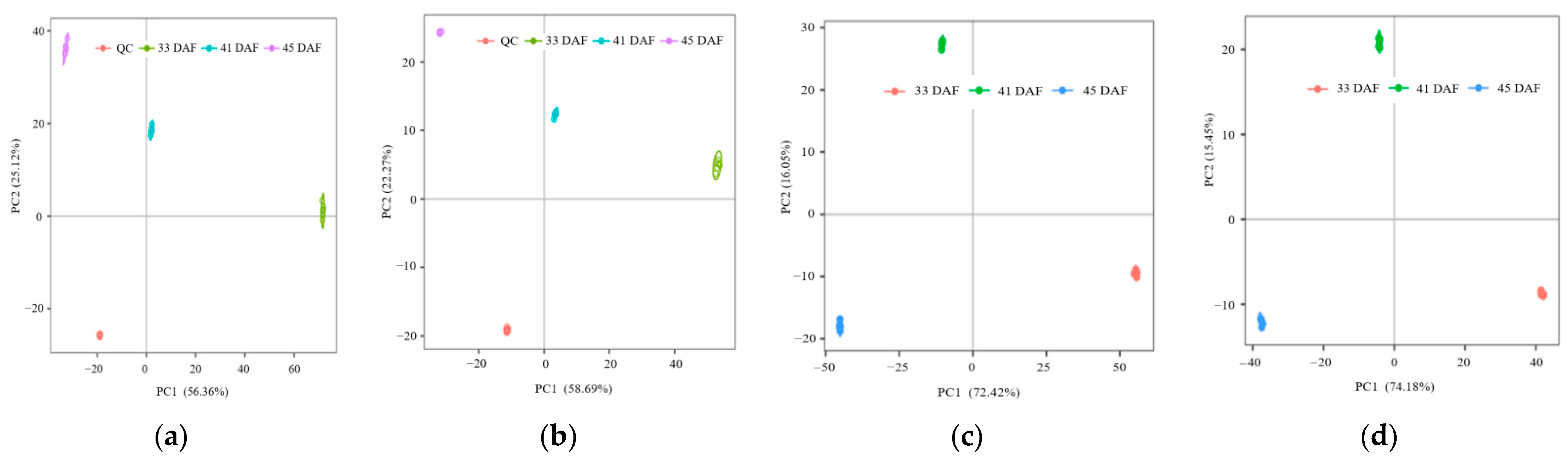
2.4. Metabolomics Data Processing and Statistical Analysis
2.5. RNA Extraction and Transcriptome Analysis
3. Results
3.1. Spatial/Temporal Pattern of Lignin Deposition in the Apricot Endocarp
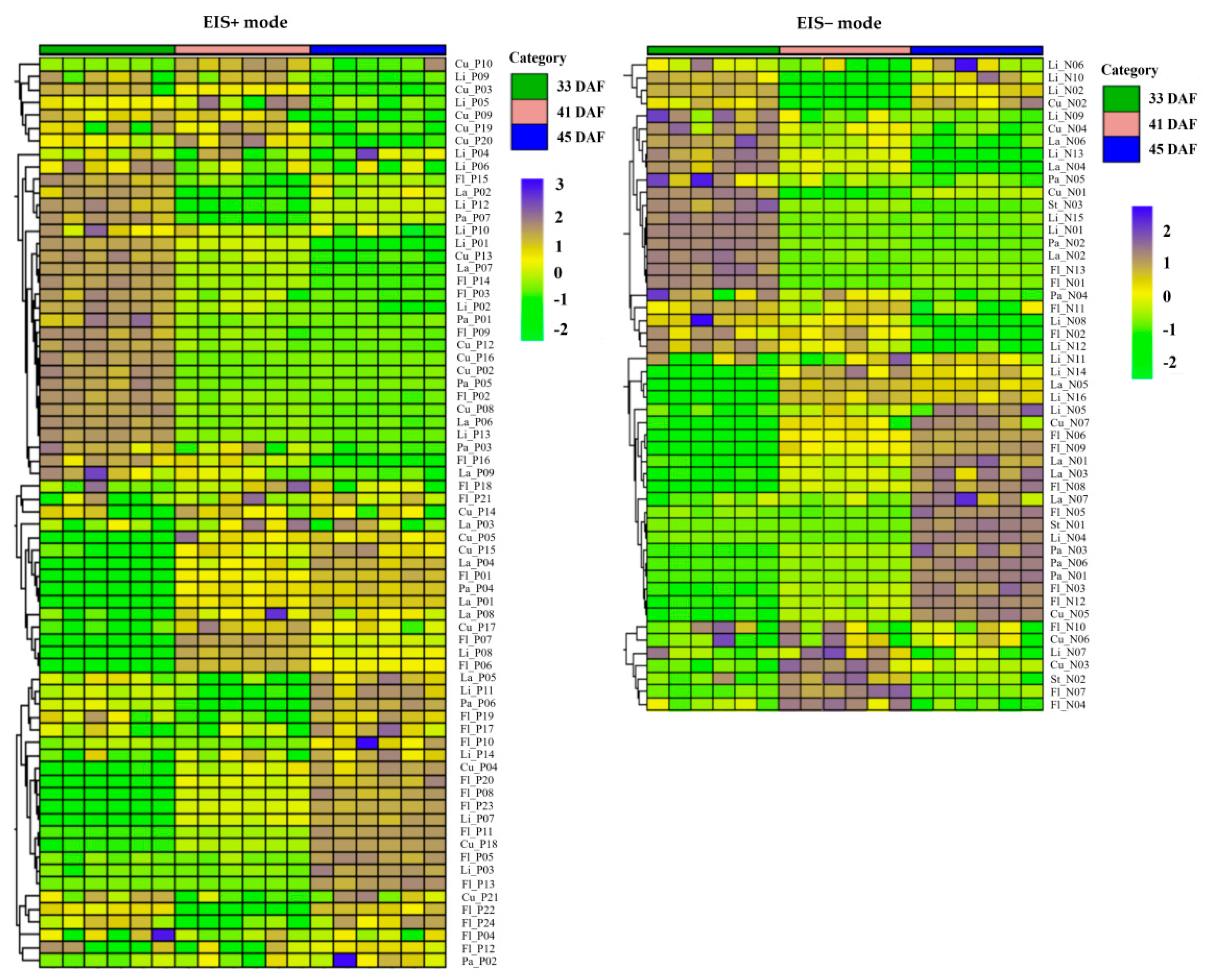
3.2. Metabolic Profiles and Metabolite Identification during Endocarp Lignification
3.3. Expression Patterns of Primary Metabolites during Endocarp Lignification
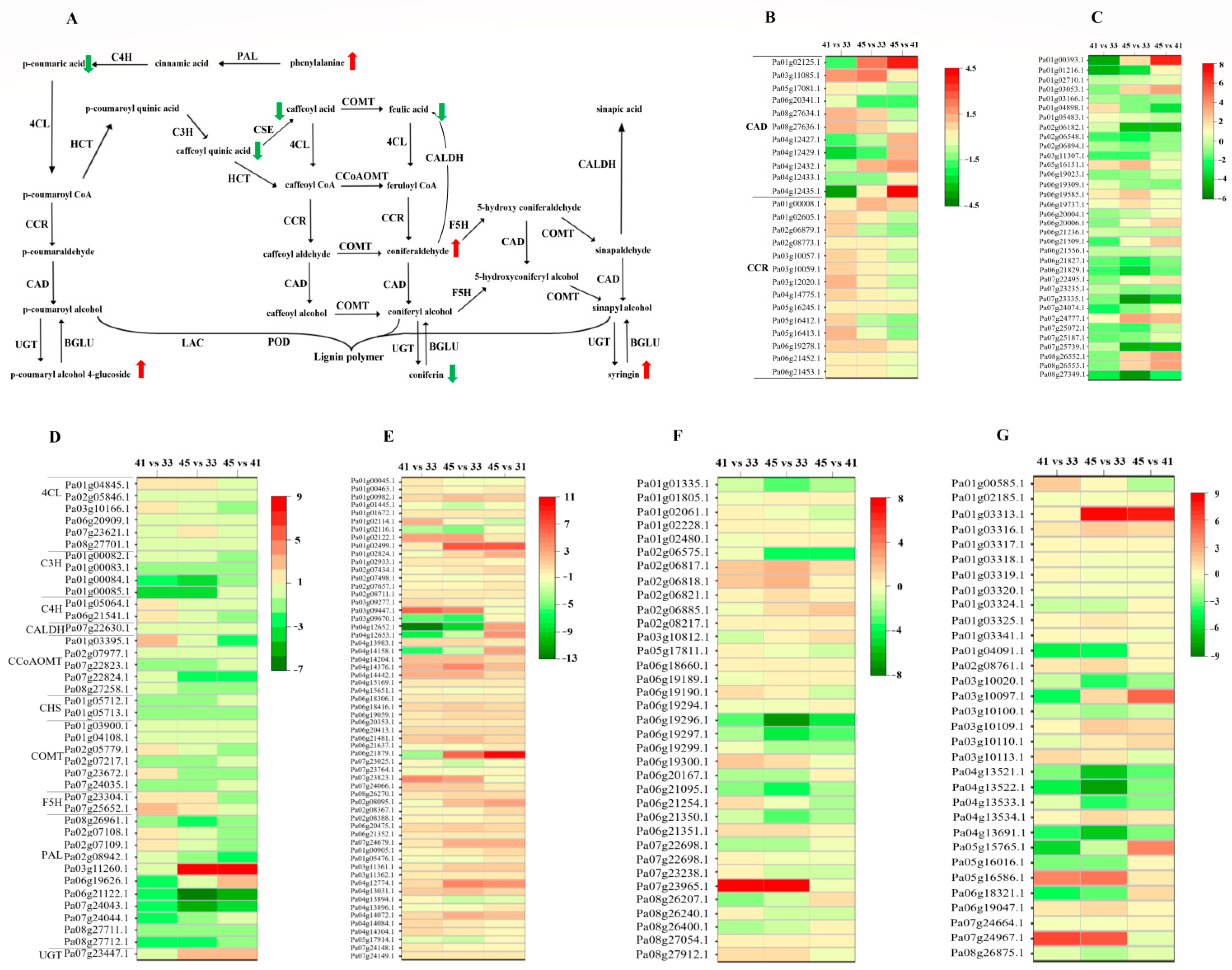
3.4. Expression Patterns of Phenylpropanoids Metabolites during Endocarp Lignification
3.5. Analysis of Lignin Metabolites before and after Lignification
3.6. Expression Patterns of Genes Associated with Lignin Metabolic Pathway during Endocarp Lignification
4. Discussion
4.1. Timing of Endocarp Lignification in Kernel-Using Apricot
4.2. Changes of Metabolites before and after Lignification of Endocarp in Kernel-Using Apricot
4.3. Regulation of a Lignified Endocarp in Kernel-Using Apricot
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, W.; Liu, N.; Zhang, Y.; Yu, X.; Sun, M.; Xu, M.; Zhang, Q.; Liu, S. Kernel-using apricot resources and its utilization. Acta Hortic. 2012, 966, 189–191. [Google Scholar] [CrossRef]
- Wang, R.H.; Li, J.R.; Chen, L.L. Prelimiary studies on tissue structure and developmental anatomy of apricot during fruit growth and development. Acta Univ. Agric. Boreali-Occident. 2000, 28, 45–50. [Google Scholar] [CrossRef]
- Shi, M.Y.; Li, Y.; Zhang, W.; Xu, J.; Liu, Y.P. Dynamic changes of lignin deposition in the endocarp of peach fruit. J. Beijing Agric. Coll. 2013, 28, 25–28. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Haji, T.; Miyake, M.; Yaegaki, H. Studies on the varietal differences and early deviation of mesocarp cell numbers and lengths and fruit weight among commercial peach [Prunus persica (L.) Batsch] cultivars and selections, wild types, and their hybrids. J. Jpn. Soc. Hortic. Sci. 2002, 71, 459–466. [Google Scholar] [CrossRef]
- Zhu, Q.P.; Guo, C.M.; Xu, J.; Tudi, M.; Gong, P.; Tuerxungu, A.; Yang, B.; Liao, K. Tissue anatomical observation of almond endocarp development. Nonwood For. Res. 2017, 35, 124–129. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, L.J.; Zhang, Q.P.; Xu, J.Y.; Liu, W.S.; Dong, W.X. Comparative transcriptome profiling and morphology provide insights into endocarp cleaving of apricot cultivar (Prunus armeniaca L.). BMC Plant Biol. 2017, 17, 72. [Google Scholar] [CrossRef]
- Lv, C.J.; Zhang, Q.P.; Liu, N.; Zhang, Y.P.; Xu, M.; Liu, S.; Ma, X.X.; Zhang, Y.J.; Liu, W.S. Correlations between physical properties and major chemical components of shells in apricot. J. Fruit Sci. 2021, 38, 1717–1724. [Google Scholar] [CrossRef]
- Vanholme, R.; De Meester, B.; Ralph, J.; Boerjan, W. Lignin biosynthesis and its integration into metabolism. Curr. Opin. Biotech. 2019, 56, 230–239. [Google Scholar] [CrossRef]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant develppment and plant-environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xie, M.; Tuskan, G.; Muchero, W.; Chen, J.G. Recent advances in the transcriptional regulation of secondary cell wall biosynthesis in the woody plants. Front. Plant Sci. 2018, 9, 1535. [Google Scholar] [CrossRef]
- Li, S.M.; Su, X.Q.; Abdullah, M.; Sun, Y.M.; Li, G.H.; Cheng, X.; Lin, Y.; Cai, Y.P.; Jin, Q. Effects of different pollens on primary metabolism and lignin biosynthesis in pear. Int. J. Mol. Sci. 2018, 19, 2273. [Google Scholar] [CrossRef]
- Barros, J.; Serk, H.; Granlundz, I.; Pesquet, E. The cell biology of lignification in higher plants. Ann. Bot. 2015, 115, 1053–1074. [Google Scholar] [CrossRef] [PubMed]
- Dardick, C.D.; Callahan, A.M.; Chiozzotto, R.; Schaffer, R.J.; Piagnani, M.C.; Scorza, R. Stone formation in peach fruit exhibits spatial coordination of the lignin and flavonoid pathways and similarity to Arabidopsis dehiscence. BMC Biol. 2010, 8, 13. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Q.P.; Sun, X.Y.; Du, X.; Liu, W.S.; Dong, W.X. Differential expression of genes encoding phenylpropanoid enzymes in an apricot cultivar (Prunus armeniaca L.) with cleavable endocarp. Trees 2019, 33, 1695–1710. [Google Scholar] [CrossRef]
- Xu, J.; Yan, J.; Li, W.; Wang, Q.; Wang, C.; Guo, J.; Geng, D.; Guan, Q.; Ma, F. Integrative analyses of widely targeted metabolic profiling and transcriptome data reveals molecular insight into metabolomic variations during apple (Malus domestica) fruit development and ripening. Int. J. Mol. Sci. 2020, 21, 4797. [Google Scholar] [CrossRef]
- Li, Z.L. Plant Sectioning Technology, 2nd ed.; Science Press: Beijing, China, 1987; pp. 105–137. [Google Scholar]
- Guida, R.D.; Engel, J.; Allwood, J.W.; Weber, R.J.M.; Jones, M.R.; Sommer, U.; Viant, M.R.; Dunn, W.B. Non-targeted UHPLC-MS metabolomic data processing methods: A comparative investigation of normalisation, missing value imputation, transformation and scaling. Metabolomics 2016, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-Mcintyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Wen, B.; Mei, Z.L.; Zeng, C.W.; Liu, S.Q. MetaX: A flexible and comprehensive software for processing metabolomics data. BMC Bioinform. 2017, 18, 183. [Google Scholar] [CrossRef] [PubMed]
- Westerhuis, J.A.; Hoefsloot, H.C.J.; Smit, S.; Vis, D.J.; Smiled, A.K.; Van Velzen, E.J.J.; Van Duijnhoven, J.P.M.; Van Dorsten, F.A. Assessment of PLSDA cross validation. Metabolomics 2008, 4, 81–89. [Google Scholar] [CrossRef]
- Cock, P.J.A.; Fields, C.J.; Goto, N.; Heuer, M.L.; Rice, P.M. The Sanger FASTQ file format for sequences with quality scores, and the Solexa/Illumina FASTQ variants. Nucleic Acids Res. 2010, 38, 1767–1771. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.P.; Zhang, D.Y.; Yu, K.; Ji, J.J.; Liu, N.; Zhang, Y.P.; Xu, M.; Zhang, Y.J.; Ma, X.X.; Liu, S.; et al. Frequent germplasm exchanges drive the high genetic diversity of Chinese-cultivated common apricot germplasm. Hortic. Res. 2021, 8, 215. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef]
- Dardick, C.D.; Callahan, A.M. Evolution of the fruit endocarp: Molecular mechanisms underlying adaptations in seed protection and dispersal strategies. Front. Plant Sci. 2014, 5, 284. [Google Scholar] [CrossRef]
- Ge, S.L.; Wang, Y.N.; Meng, H.L.; Wang, Y.M.; Guan, W.; Lu, P. Study on anatomy and pit-splitting during peach fruit development. J. Beijing Agric. Coll. 2006, 21, 1–4. [Google Scholar] [CrossRef]
- Meng, Z.; Yu, J.; Yang, A.Z.; Shi, G.L.; Zhang, T.Q.; Wang, Y.N. The lignin deposition process in fruit endocarp of different jujube cultivars. J. Agric. 2016, 6, 82–87. [Google Scholar] [CrossRef]
- Canton, M.; Drincovich, M.F.; Lara, M.V.; Vizzotto, G.; Walker, R.P.; Famiani, F.; Bonghi, C. Metabolism of stone fruits: Reciprocal contribution between primary metabolism and cell wall. Front. Plant Sci. 2020, 11, 1054. [Google Scholar] [CrossRef]
- Yang, A.Z.; Zhang, Z.Y.; Cao, A.J.; Meng, H.L.; Wang, Y.N. Studies of changes in sugar accumulation and lignin deposition during peach fruit endocarp development. Acta Hortic. Sin. 2009, 36, 1113–1119. [Google Scholar] [CrossRef]
- Zhu, Q.P.; Guo, C.M.; Mubarak, A.; Gong, P.; Shu, R.; Yang, B.; Liao, K. Changes in relative enzyme activities during the lignification in the almond endocarp. J. Fruit Sci. 2018, 35, 1079–1086. [Google Scholar] [CrossRef]
- Stewart, J.J.; Akiyama, T.; Chapple, C.C.S.; Ralph, J.; Mansfield, S.D. The effects on lignin structure on overexpression of ferulate 5-hydroxylase in hybrid poplar. Plant Physiol. 2009, 150, 621–635. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Davis, E.; Gardner, D.; Cai, X.; Wu, Y. Involvement of AtLAC15 in lignin synthesis in seeds and in root elongation of Arabidopsis. Planta 2006, 224, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Lillo, C.; Lea, U.S.; Ruoff, P. Nutrient depletion as a key factor for manipulating gene expression and product formation in different branches of the flavonoid pathway. Plant Cell Environ. 2008, 31, 582–601. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ion Mode | Total | Amino Acids | Sugars | Organic Acids | Coumarins | Lignins | Lignans | Phenolic Acids | Flavonoids | Others |
|---|---|---|---|---|---|---|---|---|---|---|
| EIS+ | 390 | 12 | 6 | 2 | 21 | 14 | 9 | 7 | 24 | 295 |
| EIS− | 236 | 8 | 15 | 12 | 7 | 16 | 7 | 6 | 13 | 152 |
| Metabolites | Ion | 33 DAF | 41 DAF | 45 DAF | ANOVA_P | KW_P |
|---|---|---|---|---|---|---|
| DL-Arginine | + | 73.11 | 4.06 | 9.19 | 0.000 | 0.001 |
| L-Aspartic acid | + | 0.88 | 1.57 | 0.84 | 0.000 | 0.003 |
| L-Aspartic acid | − | 7.05 | 11.85 | 10.48 | 0.000 | 0.002 |
| Asparagine | + | 23.04 | 16.99 | 26.11 | 0.033 | 0.018 |
| Asparagine | − | 83.89 | 54.07 | 82.69 | 0.000 | 0.004 |
| D-Glutamic acid | + | 22.89 | 25.75 | 13.98 | 0.000 | 0.001 |
| L-Glutamic acid | − | 12.76 | 14.35 | 8.43 | 0.000 | 0.001 |
| L-Pyroglutamic acid | + | 7.54 | 8.14 | 6.92 | 0.001 | 0.002 |
| D-(−)-Glutamine | + | 2.46 | 2.59 | 2.78 | 0.067 | 0.045 |
| D-(−)-Glutamine | − | 1.88 | 2.00 | 1.96 | 0.281 | 0.241 |
| L-Histidine | + | 3.00 | 1.34 | 4.61 | 0.000 | 0.001 |
| L-Histidine | − | 4.50 | 1.18 | 4.58 | 0.000 | 0.003 |
| DL-β-Leucine | − | 0.14 | 0.46 | 0.92 | 0.000 | 0.001 |
| L-Isoleucine | + | 2.42 | 5.24 | 15.16 | 0.000 | 0.001 |
| Isoleucine | + | 0.67 | 1.43 | 3.59 | 0.000 | 0.001 |
| L-Norleucine | + | 1.11 | 2.72 | 7.10 | 0.000 | 0.001 |
| L-Phenylalanine | − | 3.67 | 4.64 | 21.22 | 0.000 | 0.002 |
| L-Tyrosine | + | 0.63 | 1.03 | 2.65 | 0.000 | 0.004 |
| D-(+)-Tryptophan | + | 0.92 | 6.47 | 32.28 | 0.000 | 0.001 |
| D-(+)-Tryptophan | − | 0.33 | 2.59 | 12.69 | 0.000 | 0.001 |
| α-Oxoglutaric acid | − | 210.29 | 177.71 | 386.87 | 0.000 | 0.001 |
| α-Hydroxyglutaric acid | − | 3.21 | 4.08 | 1.29 | 0.486 | 0.103 |
| Citric acid | − | 105.56 | 65.82 | 213.98 | 0.000 | 0.001 |
| Isocitric acid | − | 19.59 | 14.55 | 23.65 | 0.000 | 0.001 |
| Fumaric acid | − | 61.35 | 55.14 | 47.78 | 0.053 | 0.030 |
| Levulinic acid | − | 0.09 | 0.15 | 0.31 | 0.000 | 0.001 |
| DL-Malic acid | − | 2502.17 | 2037.39 | 1760.69 | 0.000 | 0.003 |
| 2-Isopropylmalic acid | − | 6.62 | 9.79 | 27.44 | 0.000 | 0.001 |
| D-(−)-Quinic acid | + | 3.08 | 2.38 | 1.33 | 0.000 | 0.001 |
| D-(−)-Quinic acid | − | 536.46 | 440.19 | 364.91 | 0.033 | 0.045 |
| Shikimic acid | − | 0.48 | 0.29 | 0.25 | 0.000 | 0.001 |
| Succinic acid | − | 17.94 | 11.28 | 7.70 | 0.000 | 0.001 |
| trans-Aconitic acid | + | 2.54 | 2.12 | 5.06 | 0.000 | 0.001 |
| trans-Aconitic acid | − | 12.64 | 9.09 | 14.45 | 0.000 | 0.001 |
| 5-Hydroxymethylfurfural | + | 1.72 | 0.86 | 0.41 | 0.000 | 0.001 |
| α-Lactose | + | 8.57 | 5.68 | 2.90 | 0.000 | 0.001 |
| α-Lactose | − | 15.67 | 11.05 | 8.24 | 0.001 | 0.001 |
| Galactitol | + | 8.75 | 4.56 | 1.85 | 0.000 | 0.001 |
| D-(−)-Lyxose | − | 0.89 | 0.67 | 0.66 | 0.124 | 0.034 |
| D-(−)-Fructose | − | 0.26 | 0.42 | 0.70 | 0.000 | 0.001 |
| D-Fructose-1,6-diphosphate | + | 0.52 | 0.46 | 0.39 | 0.000 | 0.001 |
| D-Fructose-1,6-diphosphate | − | 8.81 | 7.50 | 6.09 | 0.000 | 0.001 |
| D-(+)-Glucose | + | 2.93 | 1.61 | 0.75 | 0.000 | 0.001 |
| Glucose 1-phosphate | − | 10.44 | 8.99 | 5.40 | 0.000 | 0.001 |
| D-Glucosamine | + | 0.65 | 0.41 | 0.59 | 0.000 | 0.001 |
| Gluconic acid | − | 14.07 | 10.94 | 8.76 | 0.000 | 0.001 |
| D-(+)-Maltose | − | 1.61 | 1.48 | 1.14 | 0.004 | 0.008 |
| Mannitol | − | 91.64 | 61.25 | 35.55 | 0.000 | 0.001 |
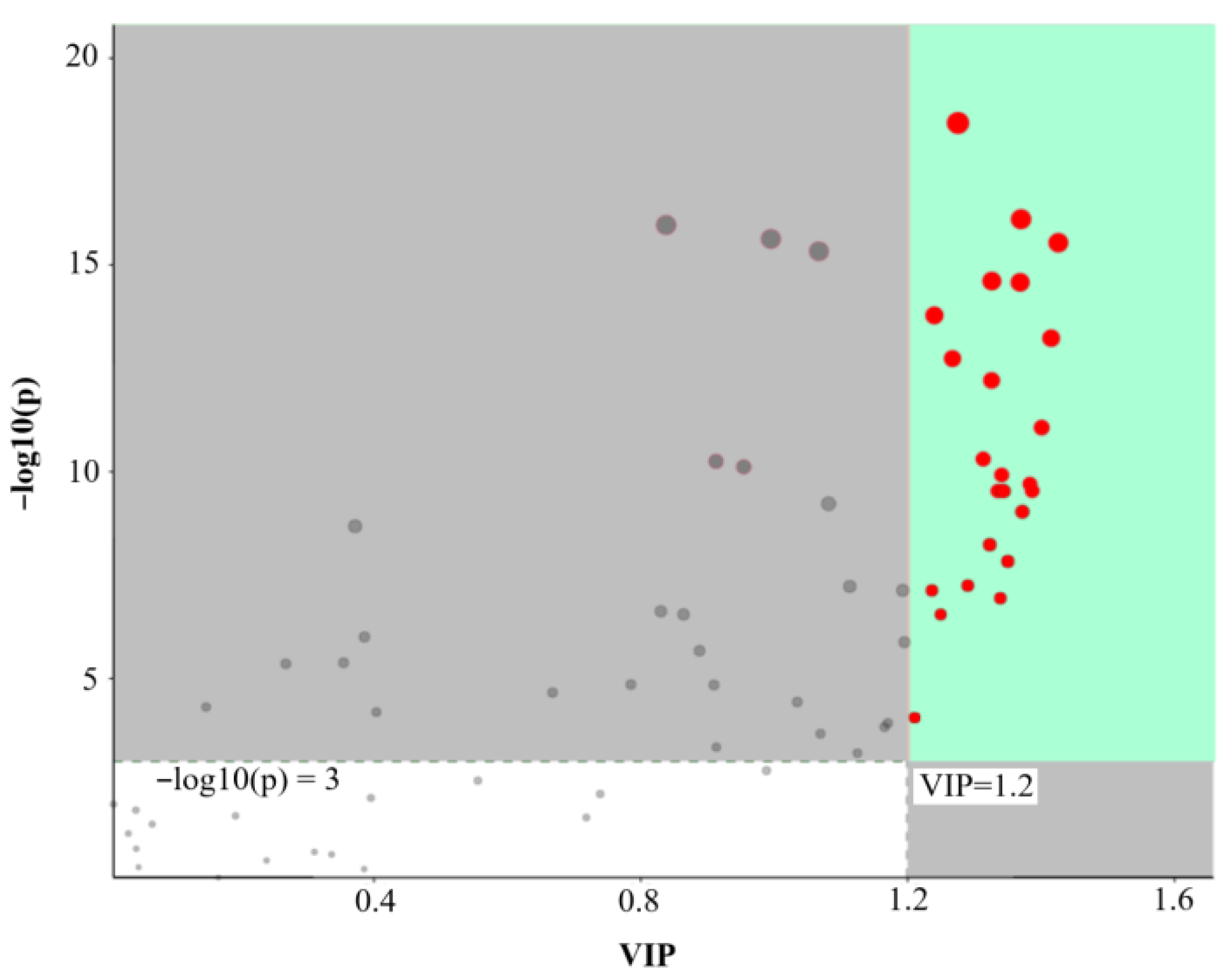
| Code | Metabolite | Regulation | VIP | p Value | 41 vs. 33 | 45 vs. 33 | ||
|---|---|---|---|---|---|---|---|---|
| FC | log2(FC) | FC | log2(FC) | |||||
| La_N01 | forsythin | up | 1.36 | 6.18 × 10−15 | 3.08 | 1.62 | 9.12 | 3.19 |
| Li_N14 | sibiricose A6 | up | 1.36 | 3.06 × 10−13 | 3.06 | 1.61 | 6.11 | 2.61 |
| Li_P07 | rosavin | up | 1.32 | 2.56 × 10−15 | 14.07 | 3.81 | 29.72 | 4.89 |
| Li_N04 | p-coumaryl alcohol 4-glucoside | up | 1.31 | 9.24 × 10−9 | 1.91 | 0.93 | 2.46 | 1.30 |
| La_P03 | l-asarinin | up | 1.28 | 7.02 × 10−15 | 5.87 | 2.55 | 7.11 | 2.83 |
| Cu_P18 | fraxinol | up | 1.27 | 5.58 × 10−11 | 3.63 | 1.86 | 4.11 | 2.04 |
| Li_N15 | tenuifoliside B | up | 1.25 | 1.40 × 10−7 | 2.11 | 1.08 | 2.39 | 1.25 |
| Li_P08 | isochlorogenic acid C | up | 1.22 | 1.56 × 10−18 | 54.16 | 5.76 | 60.32 | 5.91 |
| Cu_N03 | fraxetin | up | 1.21 | 1.26 × 10−12 | 10.32 | 3.37 | 10.48 | 3.39 |
| Cu_P19 | dihydrocoumarin | up | 1.20 | 1.05 × 10−7 | 1.33 | 0.41 | 3.37 | 1.75 |
| Li_P11 | coniferin | down | 1.34 | 1.31 × 10−10 | 0.74 | −0.44 | 0.38 | −1.40 |
| La_N03 | cycloolivil | down | 1.33 | 4.64 × 10−11 | 0.45 | −1.15 | 0.22 | −2.18 |
| Li_P04 | 2-hydroxycinnamic acid | down | 1.33 | 2.55 × 10−16 | 0.60 | −0.74 | 0.11 | −3.23 |
| Li_P02 | ethyl 4-methoxycinnamate | down | 1.32 | 1.47 × 10−10 | 0.67 | −0.59 | 0.43 | −1.21 |
| La_P02 | lariciresinol 4-o-glucoside | down | 1.32 | 1.07 × 10−10 | 0.57 | −0.80 | 0.35 | −1.52 |
| La_N06 | pinoresinol 4-o-glucoside | down | 1.31 | 3.60 × 10−10 | 0.80 | −0.31 | 0.34 | −1.54 |
| La_N02 | (+)-pinoresinol | down | 1.31 | 8.28 × 10−10 | 0.80 | −0.31 | 0.34 | −1.57 |
| Cu_P13 | 4-methylumbelliferyl-α-d-glucopyranoside | down | 1.30 | 4.78 × 10−10 | 0.61 | −0.71 | 0.40 | −1.33 |
| Li_P05 | 3-hydroxycinnamic acid | down | 1.30 | 1.03 × 10−11 | 0.80 | −0.33 | 0.24 | −2.09 |
| Cu_P08 | dicumarol | down | 1.29 | 2.46 × 10−8 | 0.47 | −1.08 | 0.22 | −2.16 |
| La_N04 | lariciresinol 4-o-glucoside | down | 1.28 | 6.03 × 10−11 | 0.47 | −1.08 | 0.31 | −1.67 |
| Li_N02 | p-coumaric acid | down | 1.26 | 3.06 × 10−9 | 0.88 | −0.19 | 0.32 | −1.63 |
| Li_N11 | ferulic acid | down | 1.24 | 2.65 × 10−8 | 0.98 | −0.02 | 0.47 | −1.09 |
| Li_N10 | neochlorogenic acid | down | 1.22 | 7.12 × 10−6 | 0.59 | −0.77 | 0.33 | −1.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Ma, X.; Liu, W.; Liu, N.; Zhang, Y.; Xu, M.; Liu, S.; Zhang, Y.; Zhao, H.; Liu, J. Metabolomics Data Revealed Metabolite Changes during Endocarp Lignification in Kernel-Using Apricot. Horticulturae 2022, 8, 967. https://doi.org/10.3390/horticulturae8100967
Zhang Q, Ma X, Liu W, Liu N, Zhang Y, Xu M, Liu S, Zhang Y, Zhao H, Liu J. Metabolomics Data Revealed Metabolite Changes during Endocarp Lignification in Kernel-Using Apricot. Horticulturae. 2022; 8(10):967. https://doi.org/10.3390/horticulturae8100967
Chicago/Turabian StyleZhang, Qiuping, Xiaoxue Ma, Weisheng Liu, Ning Liu, Yuping Zhang, Ming Xu, Shuo Liu, Yujun Zhang, Haijuan Zhao, and Jiacheng Liu. 2022. "Metabolomics Data Revealed Metabolite Changes during Endocarp Lignification in Kernel-Using Apricot" Horticulturae 8, no. 10: 967. https://doi.org/10.3390/horticulturae8100967
APA StyleZhang, Q., Ma, X., Liu, W., Liu, N., Zhang, Y., Xu, M., Liu, S., Zhang, Y., Zhao, H., & Liu, J. (2022). Metabolomics Data Revealed Metabolite Changes during Endocarp Lignification in Kernel-Using Apricot. Horticulturae, 8(10), 967. https://doi.org/10.3390/horticulturae8100967