
Foliar Application of ZnO-NPs Influences Chlorophyll Fluorescence and Antioxidants Pool in Capsicum annum L. under Salinity

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Preparation of ZnO-NPs
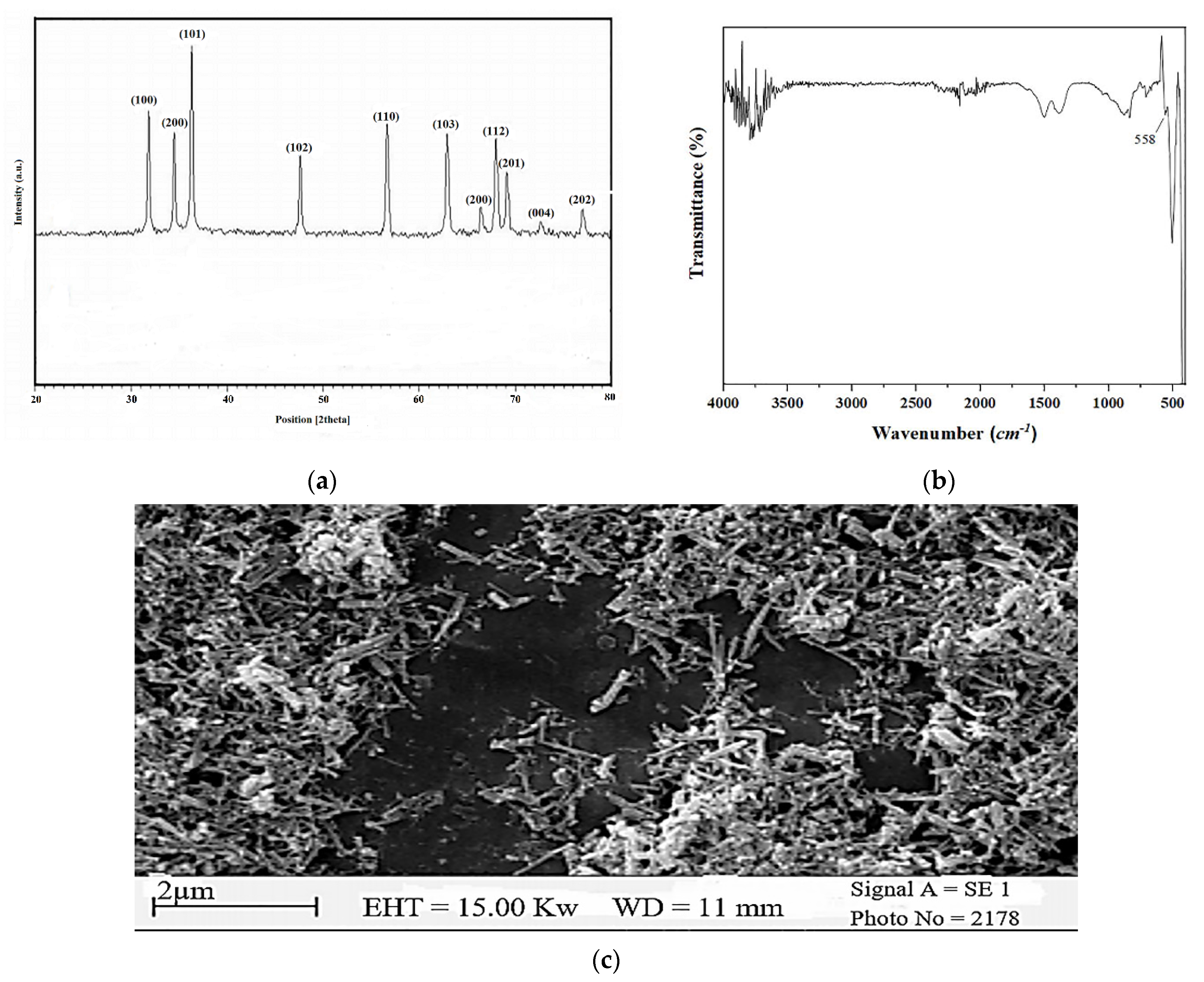
XRD Spectrum, FT-IR Spectrum, and FE-SEM Images
2.3. Chlorophyll Index
2.4. Measurement of Chlorophyll Fluorescence Parameters
2.5. Na+ and K+ Concentration
2.6. Proline
2.7. H2O2 Content
2.8. Electrolyte Leakage (EL)
2.9. Enzymatic Antioxidants Activity
2.9.1. Guaiacol Peroxidase (GPX)
2.9.2. Superoxide Dismutase Activity (SOD)
2.9.3. Ascorbate Peroxidase Activity (APX)
2.9.4. Glutathione Reductase (GR) Activity
2.10. Non-Enzymatic Antioxidants Assay
2.10.1. Ascorbate (AsA) Assay
2.10.2. Glutathione
2.11. Statistical Analysis
3. Results
3.1. Chlorophyll Fluorescence Parameters
3.2. Ionic Homeostasis
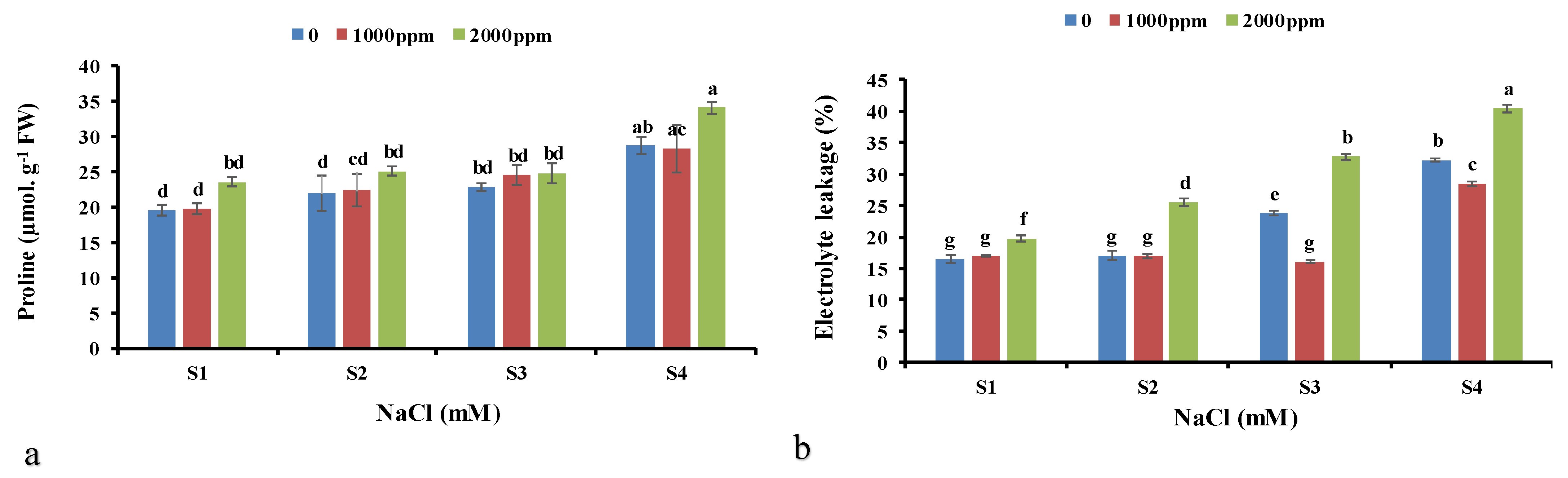
3.3. Proline
3.4. Electrolyte Leakage (EL)
3.5. Hydrogen Peroxide (H2O2)
3.6. The Activity of Enzymatic Antioxidants
3.7. The Activity of Non-Enzymatic Antioxidants
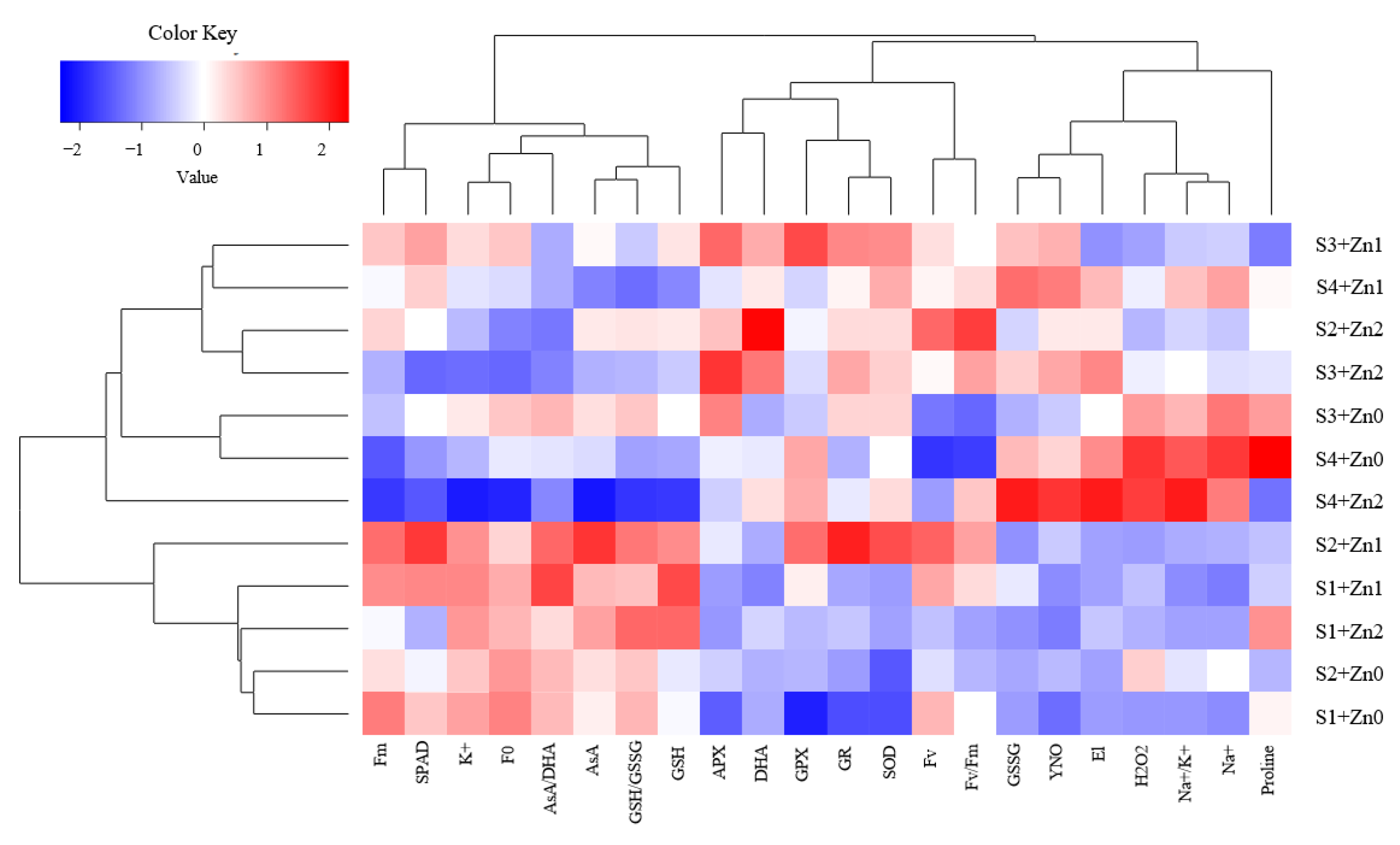
3.8. Correlation Matrix and Relative Expressions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Labrada, F.; López-Vargas, E.R.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Responses of tomato plants under saline stress to foliar application of copper nanoparticles. Plants 2019, 8, 151. [Google Scholar] [CrossRef] [PubMed]
- Jenks, M.A.; Hasegawa, P.M.; Jain, S.M.; Foolad, M. Advances in Molecular Breeding toward Drought and Salt Tolerant Crops; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Naveed, M.; Sajid, H.; Mustafa, A.; Niamat, B.; Ahmad, Z.; Yaseen, M.; Kamran, M.; Rafique, M.; Ahmar, S.; Chen, J.-T. Alleviation of salinity-induced oxidative stress, improvement in growth, physiology and mineral nutrition of canola (Brassica napus L.) through calcium-fortified composted animal manure. Sustainability 2020, 12, 846. [Google Scholar] [CrossRef]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New insights on plant salt tolerance mechanisms and their potential use for breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Li, R.-h.; Guo, P.-G.; Michael, B.; Stefania, G.; Salvatore, C. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Havaux, M.; Lannoye, R. Chlorophyll fluorescence induction: A sensitive indicator of water stress in maize plants. Irrig. Sci. 1983, 4, 147–151. [Google Scholar]
- Cerovic, Z.G.; Samson, G.; Morales, F.; Tremblay, N.; Moya, I. Ultraviolet-induced fluorescence for plant monitoring: Present state and prospects. Agronomie 1999, 19, 543–578. [Google Scholar] [CrossRef]
- Ouerghi, Z.; Cornic, G.; Roudani, M.; Ayadi, A.; Brulfert, J. Effect of NaCl on photosynthesis of two wheat species (Triticum durum and T. aestivum) differing in their sensitivity to salt stress. J. Plant Physiol. 2000, 156, 335–340. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Suleyman, I.A.; Allakhverdiev, A.S.; Yoshitaka, N.; Masami, I.; Norio, M. Ionic and osmotic effects of NaCl-induced inactivation of photosystems I and II in Synechococcus sp. Plant Physiol. 2000, 123, 1047–1056. [Google Scholar]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef]
- Baybordi, A. Zinc in Soils and Crop Nutrition; Parivar Press: Tehran, Iran, 2006; p. 179. [Google Scholar]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from plant tissues. In Plant Molecular Biology Manual; Springer: Berlin/Heidelberg, Germany, 1989; pp. 73–83. [Google Scholar]
- Aktaş, H.; ABAK, K.; Öztürk, L.; Çakmak, İ. The effect of zinc on growth and shoot concentrations of sodium and potassium in pepper plants under salinity stress. Turkish J. Agric. For. 2006, 30, 407–412. [Google Scholar]
- Saleh, J.; Maftoun, M.; Safarzadeh, S.; Gholami, A. Growth, mineral composition, and biochemical changes of broad bean as affected by sodium chloride and zinc levels and sources. Commun. Soil Sci. Plant Anal. 2009, 40, 3046–3060. [Google Scholar] [CrossRef]
- Nadeem, F.; Azhar, M.; Anwar-ul-Haq, M.; Sabir, M.; Samreen, T.; Tufail, A.; Awan, H.U.M.; Juan, W. Comparative response of two rice (Oryza sativa L.) cultivars to applied zinc and manganese for mitigation of salt stress. J. Soil Sci. Plant Nutr. 2020, 20, 2059–2072. [Google Scholar] [CrossRef]
- Tolay, I. The impact of different Zinc (Zn) levels on growth and nutrient uptake of Basil (Ocimum basilicum L.) grown under salinity stress. PLoS ONE 2021, 16, e0246493. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.N.; Rasheed, R.; Ashraf, M.Y.; Ashraf, M.A.; Hussain, I. Exogenously applied zinc and copper mitigate salinity effect in maize (Zea mays L.) by improving key physiological and biochemical attributes. Environ. Sci. Pollut. Res. 2018, 25, 23883–23896. [Google Scholar] [CrossRef]
- García-Sánchez, S.; Bernales, I.; Cristobal, S. Early response to nanoparticles in the Arabidopsis transcriptome compromises plant defence and root-hair development through salicylic acid signalling. BMC Genom. 2015, 16, 341. [Google Scholar] [CrossRef] [PubMed]
- Kisan, B.; Shruthi, H.; Sharanagouda, H.; Revanappa, S.; Pramod, N. Effect of nano-zinc oxide on the leaf physical and nutritional quality of spinach. Agrotechnology 2015, 5, 135. [Google Scholar]
- Reynolds, G.H. Nanotechnology and regulatory policy: Three futures. Harv. J. L. Tech. 2003, 17, 179. [Google Scholar]
- Prasad, R.Y.; McGee, J.K.; Killius, M.G.; Suarez, D.A.; Blackman, C.F.; DeMarini, D.M.; Simmons, S.O. Investigating oxidative stress and inflammatory responses elicited by silver nanoparticles using high-throughput reporter genes in HepG2 cells: Effect of size, surface coating, and intracellular uptake. Toxicol. Vitr. 2013, 27, 2013–2021. [Google Scholar] [CrossRef]
- Sávoly, Z.; Hrács, K.; Pemmer, B.; Streli, C.; Záray, G.; Nagy, P.I. Uptake and toxicity of nano-ZnO in the plant-feeding nematode, Xiphinema vuittenezi: The role of dissolved zinc and nanoparticle-specific effects. Environ. Sci. Pollut. Res. 2016, 23, 9669–9678. [Google Scholar] [CrossRef] [PubMed]
- Kahru, A.; Dubourguier, H.-C. From ecotoxicology to nanoecotoxicology. Toxicology 2010, 269, 105–119. [Google Scholar] [CrossRef]
- Faizan, M.; Bhat, J.A.; Chen, C.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P.; Yu, F. Zinc oxide nanoparticles (ZnO-NPs) induce salt tolerance by improving the antioxidant system and photosynthetic machinery in tomato. Plant Physiol. Biochem. 2021, 161, 122–130. [Google Scholar] [CrossRef]
- Cheema, A.; Padmanabhan, P.; Amer, A.; Parry, M.J.; Lim, L.-T.; Subramanian, J.; Paliyath, G. Postharvest hexanal vapor treatment delays ripening and enhances shelf life of greenhouse grown sweet bell pepper (Capsicum annum L.). Postharvest Biol. Technol. 2018, 136, 80–89. [Google Scholar] [CrossRef]
- Uddling, J.; Gelang-Alfredsson, J.; Piikki, K.; Pleijel, H. Evaluating the relationship between leaf chlorophyll concentration and SPAD-502 chlorophyll meter readings. Photosynth. Res. 2007, 91, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Al Abdullah, K.; Awad, S.; Zaraket, J.; Salame, C. Synthesis of ZnO nanopowders by using sol-gel and studying their structural and electrical properties at different temperature. Energy Procedia 2017, 119, 565–570. [Google Scholar] [CrossRef]
- Ghosh, K. Methods of Analysis of Soils, Plants, Waters and Fertilisers. J. Indian Soc. Soil Sci. 1993, 41, 814–815. [Google Scholar]
- Bates, J. Tell it how it is: Few school leavers understand what nursing is all about. Nurs. Stand. 2004, 18, 25–26. [Google Scholar]
- Sinha, S.; Saxena, R.; Singh, S. Chromium induced lipid peroxidation in the plants of Pistia stratiotes L.: Role of antioxidants and antioxidant enzymes. Chemosphere 2005, 58, 595–604. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.; Bouharmont, J. Effects of salt stress on growth, mineral nutrition and proline accumulation in relation to osmotic adjustment in rice (Oryza sativa L.) cultivars differing in salinity resistance. Plant Growth Regul. 1996, 19, 207–218. [Google Scholar] [CrossRef]
- Yoshimura, K.; Yabuta, Y.; Ishikawa, T.; Shigeoka, S. Expression of spinach ascorbate peroxidase isoenzymes in response to oxidative stresses. Plant Physiol. 2000, 123, 223–234. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Karuppanapandian, T.; Moon, J.-C.; Kim, C.; Manoharan, K.; Kim, W. Reactive oxygen species in plants: Their generation, signal transduction, and scavenging mechanisms. Aust. J. Crop Sci. 2011, 5, 709–725. [Google Scholar]
- Sairam, R.; Srivastava, G. Changes in antioxidant activity in sub-cellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Sci. 2002, 162, 897–904. [Google Scholar] [CrossRef]
- Law, M.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, M.; Song, Y.; Li, H.; Huang, H.; Shao, M.; Liu, Y.; Kang, Z. Carbon dots promote the growth and photosynthesis of mung bean sprouts. Carbon 2018, 136, 94–102. [Google Scholar] [CrossRef]
- Zhao, G.; Xu, H.; Zhang, P.; Su, X.; Zhao, H. Effects of 2, 4-epibrassinolide on photosynthesis and Rubisco activase gene expression in Triticum aestivum L. seedlings under a combination of drought and heat stress. Plant Growth Regul. 2017, 81, 377–384. [Google Scholar] [CrossRef]
- Gao, S.; Zhang, H.; Wang, X.; Deng, R.; Sun, D.; Zheng, G. ZnO-based hollow microspheres: Biopolymer-assisted assemblies from ZnO nanorods. J. Phys. Chem. B 2006, 110, 15847–15852. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Ashraf, M. Exogenous application of nitric oxide promotes growth and oxidative defense system in highly boron stressed tomato plants bearing fruit. Sci. Hortic. 2015, 185, 43–47. [Google Scholar] [CrossRef]
- Samadi, S.; Habibi, G.; Vaziri, A. Effects of exogenous salicylic acid on antioxidative responses, phenolic metabolism and photochemical activity of strawberry under salt stress. Iran. J. Plant Physiol. 2019, 9, 2685–2694. [Google Scholar]
- Zhou, Y.; Diao, M.; Chen, X.; Cui, J.; Pang, S.; Li, Y.; Hou, C.; Liu, H.-y. Application of exogenous glutathione confers salinity stress tolerance in tomato seedlings by modulating ions homeostasis and polyamine metabolism. Sci. Hortic. 2019, 250, 45–58. [Google Scholar] [CrossRef]
- Rahimi, E.; Nazari, F.; Javadi, T.; Samadi, S.; da Silva, J.A.T. Potassium-enriched clinoptilolite zeolite mitigates the adverse impacts of salinity stress in perennial ryegrass (Lolium perenne L.) by increasing silicon absorption and improving the K/Na ratio. J. Environ. Manag. 2021, 285, 112142. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Hussain, S.; Ali, B.; Ren, X.; Chen, X.; Li, Q.; Saqib, M.; Ahmad, N. Recent progress in understanding salinity tolerance in plants: Story of Na+/K+ balance and beyond. Plant Physiol. Biochem. 2021, 160, 239–256. [Google Scholar] [CrossRef]
- Noohpisheh, Z.; Amiri, H.; Mohammadi, A.; Farhadi, S. Effect of the foliar application of zinc oxide nanoparticles on some biochemical and physiological parameters of Trigonella foenum-graecum under salinity stress. Plant Biosyst.- Int. J. Deal. All Asp. Plant Biol. 2021, 155, 267–280. [Google Scholar] [CrossRef]
- Chen, Y.; Li, R.; Ge, J.; Liu, J.; Wang, W.; Xu, M.; Zhang, R.; Hussain, S.; Wei, H.; Dai, Q. Exogenous melatonin confers enhanced salinity tolerance in rice by blocking the ROS burst and improving Na+/K+ homeostasis. Environ. Exp. Bot. 2021, 189, 104530. [Google Scholar] [CrossRef]
- Yasmin, H.; Mazher, J.; Azmat, A.; Nosheen, A.; Naz, R.; Hassan, M.N.; Noureldeen, A.; Ahmad, P. Combined application of zinc oxide nanoparticles and biofertilizer to induce salt resistance in safflower by regulating ion homeostasis and antioxidant defence responses. Ecotoxicol. Environ. Saf. 2021, 218, 112262. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Responses of photosynthesis, nitrogen and proline metabolism to salinity stress in Solanum lycopersicum under different levels of nitrogen supplementation. Plant Physiol. Biochem. 2016, 109, 72–83. [Google Scholar] [CrossRef]
- Singh, P.; Arif, Y.; Siddiqui, H.; Sami, F.; Zaidi, R.; Azam, A.; Alam, P.; Hayat, S. Nanoparticles enhances the salinity toxicity tolerance in Linum usitatissimum L. by modulating the antioxidative enzymes, photosynthetic efficiency, redox status and cellular damage. Ecotoxicol. Environ. Saf. 2021, 213, 112020. [Google Scholar] [CrossRef] [PubMed]
- Sarabi, B.; Bolandnazar, S.; Ghaderi, N.; Ghashghaie, J. Genotypic differences in physiological and biochemical responses to salinity stress in melon (Cucumis melo L.) plants: Prospects for selection of salt tolerant landraces. Plant Physiol. Biochem. 2017, 119, 294–311. [Google Scholar] [CrossRef] [PubMed]
- Bakhoum, G.S.; Sadak, M.S.; Badr, E.A.E.M. Mitigation of adverse effects of salinity stress on sunflower plant (Helianthus annuus L.) by exogenous application of chitosan. Bull. Natl. Res. Cent. 2020, 44, 1–11. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. The role of endogenous nitric oxide in salicylic acid-induced up-regulation of ascorbate-glutathione cycle involved in salinity tolerance of pepper (Capsicum annuum L.) plants. Plant Physiol. Biochem. 2020, 147, 10–20. [Google Scholar] [CrossRef]
- Panda, S. Physiological impact of Zinc nanoparticle on germination of rice (Oryza sativa L) seed. J. Plant Sci. Phytopathol. 2017, 1, 062–070. [Google Scholar]
- Yusefi-Tanha, E.; Fallah, S.; Rostamnejadi, A.; Pokhrel, L.R. Zinc oxide nanoparticles (ZnONPs) as a novel nanofertilizer: Influence on seed yield and antioxidant defense system in soil grown soybean (Glycine max cv. Kowsar). Sci. Total Environ. 2020, 738, 140240. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Faraz, A.; Yusuf, M.; Khan, S.; Hayat, S. Zinc oxide nanoparticle-mediated changes in photosynthetic efficiency and antioxidant system of tomato plants. Photosynthetica 2018, 56, 678–686. [Google Scholar] [CrossRef]
- Tuna, A.L.; Kaya, C.; Dikilitaş, M.; Yokaş, İ.; Burun, B.; Altunlu, H. Comparative effects of various salicylic acid derivatives on key growth parameters and some enzyme activities in salinity stressed maize (Zea mays L.) plants. Pak. J. Bot. 2007, 39, 787–798. [Google Scholar]
- Hniličková, H.; Hnilička, F.; Orsák, M.; Hejnák, V. Effect of salt stress on growth, electrolyte leakage, Na+ and K+ content in selected plant species. Plant Soil Environ. 2019, 65, 90–96. [Google Scholar] [CrossRef]
- El-Badri, A.M.; Batool, M.; Mohamed, I.A.; Khatab, A.; Sherif, A.; Wang, Z.; Salah, A.; Nishawy, E.; Ayaad, M.; Kuai, J. Modulation of salinity impact on early seedling stage via nano-priming application of zinc oxide on rapeseed (Brassica napus L.). Plant Physiol. Biochem. 2021, 166, 376–392. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; ur Rehman, M.Z.; Adrees, M.; Arshad, M.; Qayyum, M.F.; Ali, L.; Hussain, A.; Chatha, S.A.S.; Imran, M. Alleviation of cadmium accumulation in maize (Zea mays L.) by foliar spray of zinc oxide nanoparticles and biochar to contaminated soil. Environ. Pollut. 2019, 248, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Zeeshan, M.; Hu, Y.X.; Iqbal, A.; Salam, A.; Liu, Y.X.; Muhammad, I.; Ahmad, S.; Khan, A.H.; Hale, B.; Wu, H.Y. Amelioration of AsV toxicity by concurrent application of ZnO-NPs and Se-NPs is associated with differential regulation of photosynthetic indexes, antioxidant pool and osmolytes content in soybean seedling. Ecotoxicol. Environ. Saf. 2021, 225, 112738. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Gharsallah, C.; Fakhfakh, H.; Grubb, D.; Gorsane, F. Effect of salt stress on ion concentration, proline content, antioxidant enzyme activities and gene expression in tomato cultivars. AoB Plants 2016, 8, 1–21. [Google Scholar] [CrossRef]
- Naheed, R.; Aslam, H.; Kanwal, H.; Farhat, F.; Gamar, M.I.A.; Al-Mushhin, A.A.; Jabborova, D.; Ansari, M.J.; Shaheen, S.; Aqeel, M. Growth attributes, biochemical modulations, antioxidant enzymatic metabolism and yield in Brassica napus varieties for salinity tolerance. Saudi J. Biol. Sci. 2021, 28, 5469–5479. [Google Scholar] [CrossRef]
- Ates, M.T.; Yildirim, A.B.; Turker, A.U. Enhancement of alkaloid content (galanthamine and lycorine) and antioxidant activities (enzymatic and non-enzymatic) unders salt stress in summer snowflake (Leucojum aestivum L.). S. Afr. J. Bot. 2021, 140, 182–188. [Google Scholar] [CrossRef]
- Hussain, I.; Singh, A.; Singh, N.; Singh, P. Plant-nanoceria interaction: Toxicity, accumulation, translocation and biotransformation. S. Afr. J. Bot. 2019, 121, 239–247. [Google Scholar] [CrossRef]
- Mohammadi, M.H.Z.; Panahirad, S.; Navai, A.; Bahrami, M.K.; Kulak, M.; Gohari, G. Cerium oxide nanoparticles (CeO2-NPs) improve growth parameters and antioxidant defense system in Moldavian Balm (Dracocephalum moldavica L.) under salinity stress. Plant Stress 2021, 1, 100006. [Google Scholar] [CrossRef]
- Lacerda, J.S.; Martinez, H.E.; Pedrosa, A.W.; Clemente, J.M.; Santos, R.H.; Oliveira, G.L.; Jifon, J.L. Importance of zinc for arabica coffee and its effects on the chemical composition of raw grain and beverage quality. Crop Sci. 2018, 58, 1360–1370. [Google Scholar] [CrossRef]
- Adrees, M.; Khan, Z.S.; Hafeez, M.; Rizwan, M.; Hussain, K.; Asrar, M.; Alyemeni, M.N.; Wijaya, L.; Ali, S. Foliar exposure of zinc oxide nanoparticles improved the growth of wheat (Triticum aestivum L.) and decreased cadmium concentration in grains under simultaneous Cd and water deficient stress. Ecotoxicol. Environ. Saf. 2021, 208, 111627. [Google Scholar] [CrossRef]
- Molnár, Á.; Papp, M.; Kovács, D.Z.; Bélteky, P.; Oláh, D.; Feigl, G.; Szőllősi, R.; Rázga, Z.; Ördög, A.; Erdei, L. Nitro-oxidative signalling induced by chemically synthetized zinc oxide nanoparticles (ZnO NPs) in Brassica species. Chemosphere 2020, 251, 126419. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Reichheld, J.-P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2018; pp. 3–12. [Google Scholar]
- Singh, M.; Singh, V.P.; Prasad, S.M. Nitrogen alleviates salinity toxicity in Solanum lycopersicum seedlings by regulating ROS homeostasis. Plant Physiol. Biochem. 2019, 141, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Khanna-Chopra, R.; Selote, D.S. Acclimation to drought stress generates oxidative stress tolerance in drought-resistant than-susceptible wheat cultivar under field conditions. Environ. Exp. Bot. 2007, 60, 276–283. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, J.; Wang, K.; Xu, H.; Qu, M.; Gao, Z.; Guo, L.; Xie, J. Facile and sensitive measurement of GSH/GSSG in cells by surface-enhanced Raman spectroscopy. Talanta 2021, 224, 121852. [Google Scholar] [CrossRef]
- Kataria, S.; Baghel, L.; Jain, M.; Guruprasad, K. Magnetopriming regulates antioxidant defense system in soybean against salt stress. Biocatal. Agric. Biotechnol. 2019, 18, 101090. [Google Scholar] [CrossRef]
- Chen, H.; Liu, T.; Xiang, L.; Hu, L.; Hu, X. GABA enhances muskmelon chloroplast antioxidants to defense salinity-alkalinity stress. Russ. J. Plant Physiol. 2018, 65, 674–679. [Google Scholar] [CrossRef]
- Huang, C.; Ding, G.; Gu, C.; Zhou, J.; Kuang, M.; Ji, Y.; He, Y.; Kondo, T.; Fan, J. Decreased Selenium-Binding Protein 1 Enhances Glutathione Peroxidase 1 Activity and Downregulates HIF-1α to Promote Hepatocellular Carcinoma InvasivenessDecreased SBP1 Promotes HCC through GPX1 and HIF-1α. Clin. Cancer Res. 2012, 18, 3042–3053. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Hossain, M.; Mahmud, J.-A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Manganese-induced salt stress tolerance in rice seedlings: Regulation of ion homeostasis, antioxidant defense and glyoxalase systems. Physiol. Mol. Biol. Plants 2016, 22, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Kang, G.; Li, G.; Liu, G.; Xu, W.; Peng, X.; Wang, C.; Zhu, Y.; Guo, T. Exogenous salicylic acid enhances wheat drought tolerance by influence on the expression of genes related to ascorbate-glutathione cycle. Biol. Plant. 2013, 57, 718–724. [Google Scholar] [CrossRef]
- Ahmad, P.; Alyemeni, M.N.; Al-Huqail, A.A.; Alqahtani, M.A.; Wijaya, L.; Ashraf, M.; Kaya, C.; Bajguz, A. Zinc oxide nanoparticles application alleviates arsenic (As) toxicity in soybean plants by restricting the uptake of as and modulating key biochemical attributes, antioxidant enzymes, ascorbate-glutathione cycle and glyoxalase system. Plants 2020, 9, 825. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NaCl (mM) | Zn | SPAD | Fm | F0 | Fv | Fv/Fm | Y (NO) | Na+ (mmol·g−1 DW) | K+ (mmol·g−1 DW) | Na+/K+ |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 0 | 41.6 ± 2.8 c | 5.22 ± 0.19 c | 0.86 ± 0.19 i | 4.35 ± 0.137 b | 0.83 ± 0.0053 ab | 0.33 ± 0.016 g | 0.467 ± 0.032 i | 5.573 ± 0.053 b | 0.083 ± 0.0024 gh |
| 1000 | 48.8 ± 0.9 b | 5.65 ± 0.18 a | 0.74 ± 0.18 j | 4.91 ± 0.117 a | 0.86 ± 0.0224 a | 0.37 ± 0.014 fg | 0.400 ± 0.033 g | 5.777 ± 0.047 a | 0.070 ± 0.0134 h | |
| 2000 | 24.8 ± 1.1 e | 4.87 ± 0.129 f | 0.96 ± 0.129 h | 3.91 ± 0.079 cd | 0.81 ± 0.0041 bc | 0.35 ± 0.014 g | 0.570 ± 0.024 f | 5.655 ± 0.041 ab | 0.100 ± 0.0042 g | |
| 25 | 0 | 33.1 ± 3.3 d | 5.07 ± 0.04 de | 1.10 ± 0.04 fg | 3.96 ± 0.085 c | 0.78 ± 0.0120 c | 0.44 ± 0.017 ef | 1.082 ± 0.042 d | 5.307 ± 0.066 c | 0.203 ± 0.0033 e |
| 1000 | 57.8 ± 1.6 a | 5.39 ± 0.112 b | 1.07 ± 0.112 g | 4.32 ± 0.089 b | 0.80 ± 0.0094 bc | 0.46 ± 0.032 e | 0.647 ± 0.050 f | 5.705 ± 0.064 ab | 0.113 ± 0.0129 fg | |
| 2000 | 34.3 ± 1.1 d | 4.94 ± 0.097 ef | 1.14 ± 0.097 f | 3.80 ± 0.114 d | 0.76 ± 0.0116 cd | 0.57 ± 0.018 d | 0.755 ± 0.015 e | 4.265 ± 0.068 f | 0.177 ± 0.009 e | |
| 50 | 0 | 34.4 ± 1.5 d | 4.65 ± 0.13 d | 1.24 ± 0.13 de | 3.41 ± 0.031 f | 0.73 ± 0.0037 de | 0.46 ± 0.01 ef | 1.77 ± 0.063 b | 5.02 ± 0.078 d | 0.352 ± 0.0047 c |
| 1000 | 45.6 ± 1.0 bc | 5.16 ± 0.178 cd | 1.20 ± 0.178 e | 3.96 ± 0.074 c | 0.76 ± 0.0047 cd | 0.65 ± 0.031 bd | 0.820 ± 0.022 e | 5.125 ± 0.079 d | 0.160 ± 0.0169 f | |
| 2000 | 16.4 ± 4.5 f | 4.71 ± 0.17 d | 1.28 ± 0.17 cd | 3.43 ± 0.064 f | 0.72 ± 0.0071 de | 0.67 ± 0.036 bc | 0.890 ± 0.035 e | 3.655 ± 0.079 g | 0.243 ± 0.0063 d | |
| 75 | 0 | 21.9 ± 4.5 ef | 4.47 ± 0.107 h | 1.34 ± 0.107 b | 3.13 ± 0.084 g | 0.70 ± 0.0011 e | 0.60 ± 0.034 cd | 2.085 ± 0.071 a | 4.212 ± 0.071 f | 0.495 ± 0.0021 b |
| 1000 | 40.6 ± 0.8 c | 4.95 ± 0.113 ef | 1.31 ± 0.113 bc | 3.64 ± 0.067 e | 0.73 ± 0.0087 de | 0.73 ± 0.029 b | 1.567 ± 0.065 c | 4.608 ± 0.067 e | 0.340 ± 0.0191 cd | |
| 2000 | 14.9 ± 2.4 f | 4.05 ± 0.101 i | 1.45 ± 0.101 a | 2.60 ± 0.034 h | 0.64 ± 0.0014 f | 0.83 ± 0.016 a | 1.742 ± 0.039 b | 2.963 ± 0.058 h | 0.587 ± 0.0088 a | |
| LSD at 0.05% | 0.75 | 0.0.039 | 0.17 | 0.15 | 0.045 | 0.090 | 0.074 | 0.13 | 0.052 | |
| S.O.V. | ||||||||||
| NaCl.3220 | 110.68 * | 1.366 ** | 0.576 ** | 3.582 ** | 0.044 ** | 0.244 ** | 5.970 ** | 10.02 ** | 0.535 ** | |
| ZnO-NPs | 1481.61 ** | 1.720 ** | 0.067 ** | 2.440 ** | 0.013 ** | 0.004 ns | 2.008 ** | 1.67 ** | 0.214 ** | |
| NaCl × ZnO-NPs | 732.81 ** | 0.071 ** | 0.005 * | 0.102 ** | 0.001 ** | 0.070 ** | 0.374 ** | 0.339 ** | 0.057 ** | |
| Error | 143.3 | 0.011 | 0.001 | 0.012 | 0.000 | 0.003 | 0.002 | 0.007 | 0.002 | |
| C.V. | 12.32 | 3.38 | 8.66 | 2.90 | 10.21 | 10.86 | 8.80 | 11.74 | 12.56 | |
| Mean square | ||||||||
|---|---|---|---|---|---|---|---|---|
| S.O.V. | df | Proline | Electrolyte Leakage | H2O2 Content | GPX Activity | APX Activity | SOD Activity | GR Activity |
| NaCl | 3 | 25.99 ns | 600.3 ** | 5.896 ** | 3.396 ** | 0.055 ** | 2.002 * | 0.052 ** |
| ZnO-NPs | 2 | 99.37 ** | 232.5 * | 2.764 ** | 1.363 ** | 0.017 * | 0.658 ** | 0.041 * |
| NaCl × ZnO-NPs | 5 | 77.20 ** | 36.50 ** | 0.439 ** | 0.240 ** | 0.001 ** | 0.117 ** | 0.003 ** |
| Error | 36 | 13.76 | 1.04 | 0.016 | 0.001 | 0.002 | 0.002 | 0.003 |
| C.V. | 15.07 | 7.38 | 8.38 | 13.02 | 11.42 | 14.35 | 12.84 | |
| Mean Square | |||||||
|---|---|---|---|---|---|---|---|
| S.O.V. | df | AsA | DHA | ASA/DHA | GSH | GSSG | GSH/GSG |
| NaCl | 3 | 5092.6 ** | 19,276.8 ** | 11.92 ** | 1838.2 ** | 3165.4 ** | 1.074 ** |
| ZnO-NPs | 2 | 10,354.3 ** | 7359.8 ** | 7.408 ** | 7267.1 * | 6300.5 ** | 3.555 * |
| NaCl × ZnO-NPs | 5 | 1028.02 ** | 1194.8 ** | 1.207 ** | 818.9 ** | 1066.6 ** | 0.541 ** |
| Error | 36 | 16.79 | 12.77 | 0.010 | 10.77 | 7.299 | 0.004 |
| C.V. | 9.19 | 11.92 | 13.51 | 9.11 | 10.98 | 11.94 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasouli, F.; Asadi, M.; Hassanpouraghdam, M.B.; Aazami, M.A.; Ebrahimzadeh, A.; Kakaei, K.; Dokoupil, L.; Mlcek, J. Foliar Application of ZnO-NPs Influences Chlorophyll Fluorescence and Antioxidants Pool in Capsicum annum L. under Salinity. Horticulturae 2022, 8, 908. https://doi.org/10.3390/horticulturae8100908
Rasouli F, Asadi M, Hassanpouraghdam MB, Aazami MA, Ebrahimzadeh A, Kakaei K, Dokoupil L, Mlcek J. Foliar Application of ZnO-NPs Influences Chlorophyll Fluorescence and Antioxidants Pool in Capsicum annum L. under Salinity. Horticulturae. 2022; 8(10):908. https://doi.org/10.3390/horticulturae8100908
Chicago/Turabian StyleRasouli, Farzad, Mohammad Asadi, Mohammad Bagher Hassanpouraghdam, Mohammad Ali Aazami, Asghar Ebrahimzadeh, Karim Kakaei, Libor Dokoupil, and Jiri Mlcek. 2022. "Foliar Application of ZnO-NPs Influences Chlorophyll Fluorescence and Antioxidants Pool in Capsicum annum L. under Salinity" Horticulturae 8, no. 10: 908. https://doi.org/10.3390/horticulturae8100908
APA StyleRasouli, F., Asadi, M., Hassanpouraghdam, M. B., Aazami, M. A., Ebrahimzadeh, A., Kakaei, K., Dokoupil, L., & Mlcek, J. (2022). Foliar Application of ZnO-NPs Influences Chlorophyll Fluorescence and Antioxidants Pool in Capsicum annum L. under Salinity. Horticulturae, 8(10), 908. https://doi.org/10.3390/horticulturae8100908