
Integrated Volatile Metabolomics and Transcriptomics Analyses Reveal the Influence of Infection TuMV to Volatile Organic Compounds in Brassica rapa

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Detection of TuMV Resistance in Brassica rapa
2.3. Material Cultivation
2.3.1. TuMV Inoculation
2.3.2. TuMV-ELISA Test
2.3.3. Isolation and Concentration of Volatiles
2.3.4. RNA Extraction and Library Construction
- Data Analyses
3. Results
3.1. Phenotype Analysis of Resistance and Susceptibility to TuMV in B. rapa
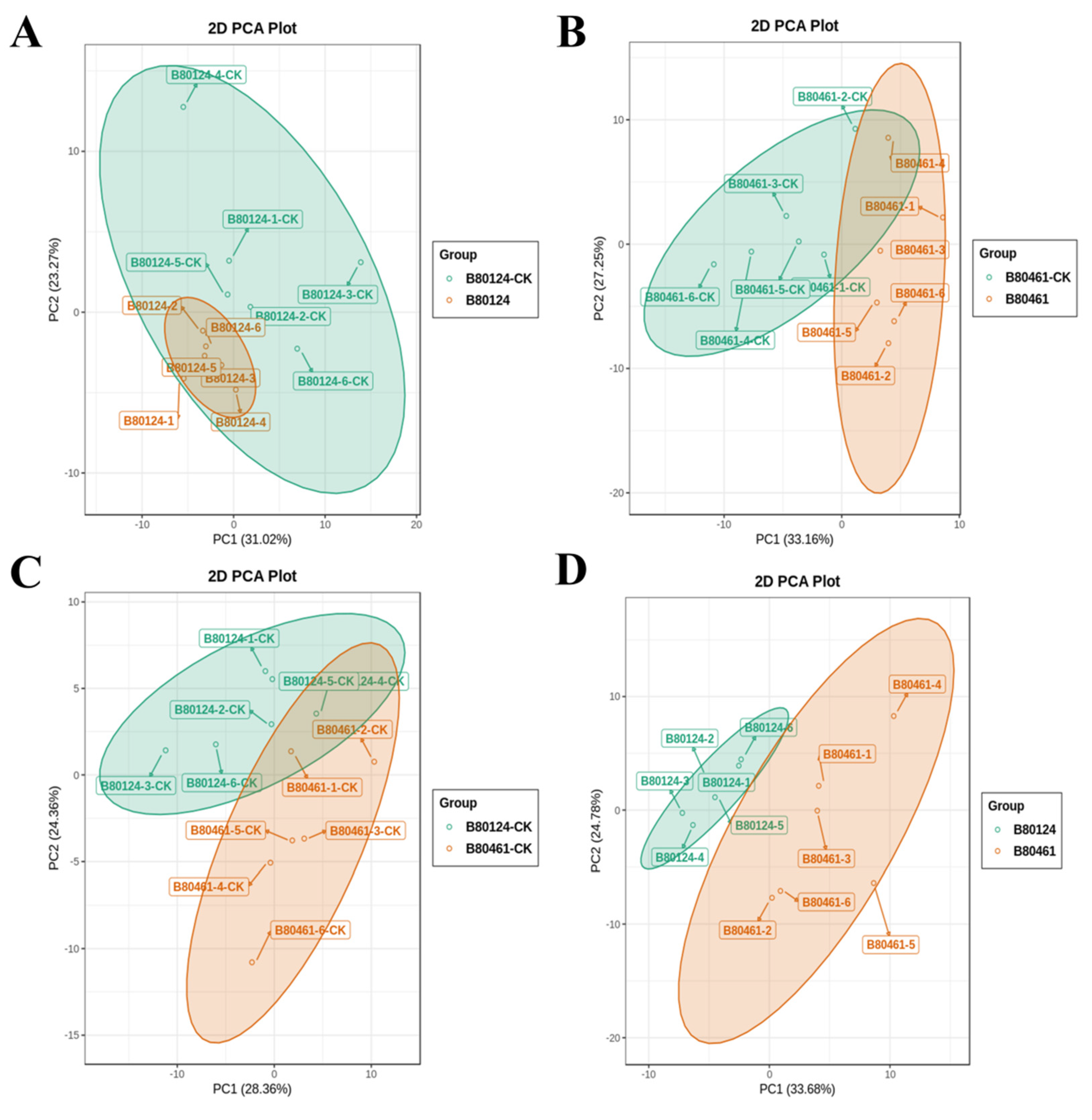
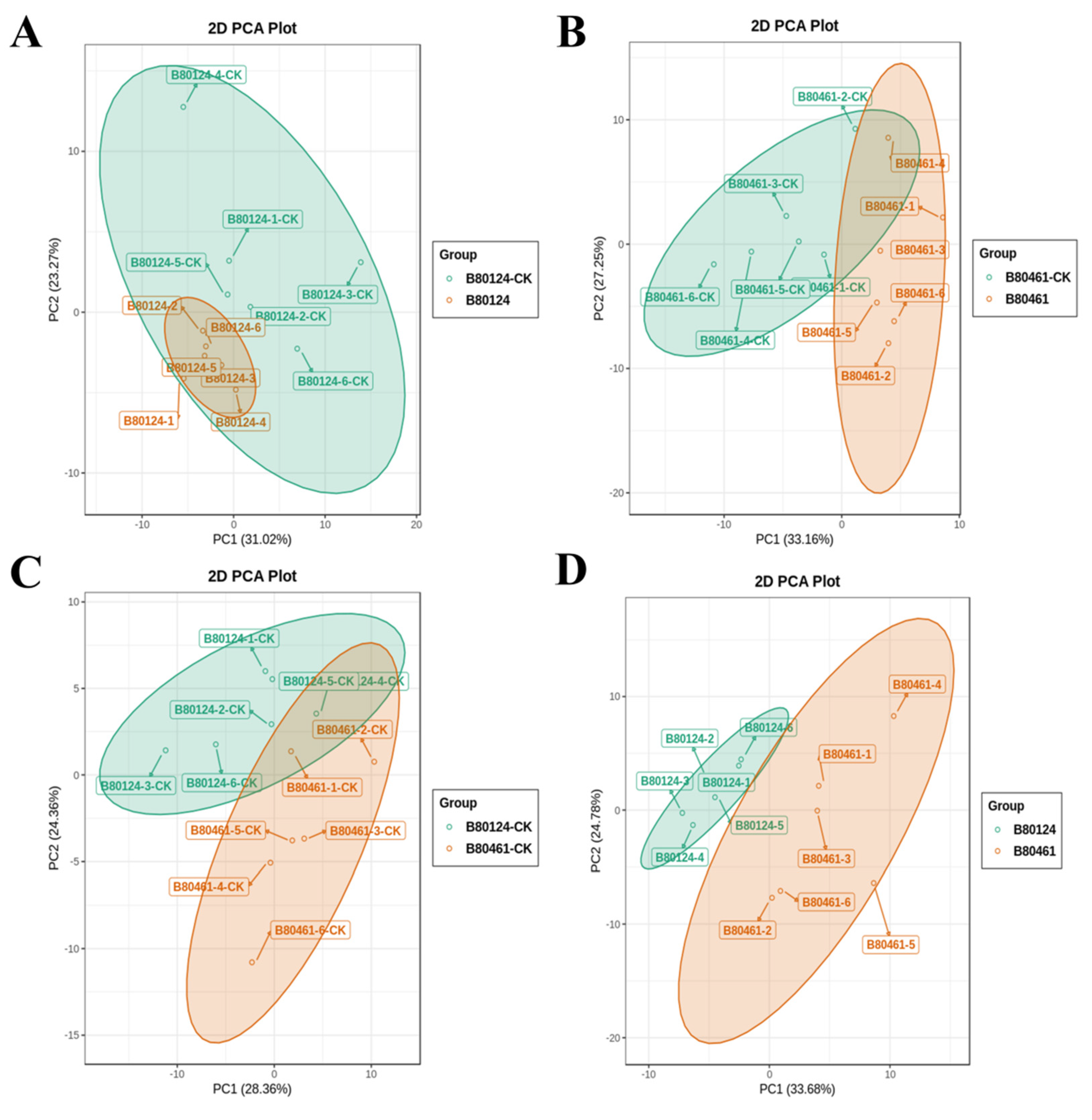
3.2. Volatile Metabolome Analysis of Volatile Organic Compound Accumulation in B. rapa
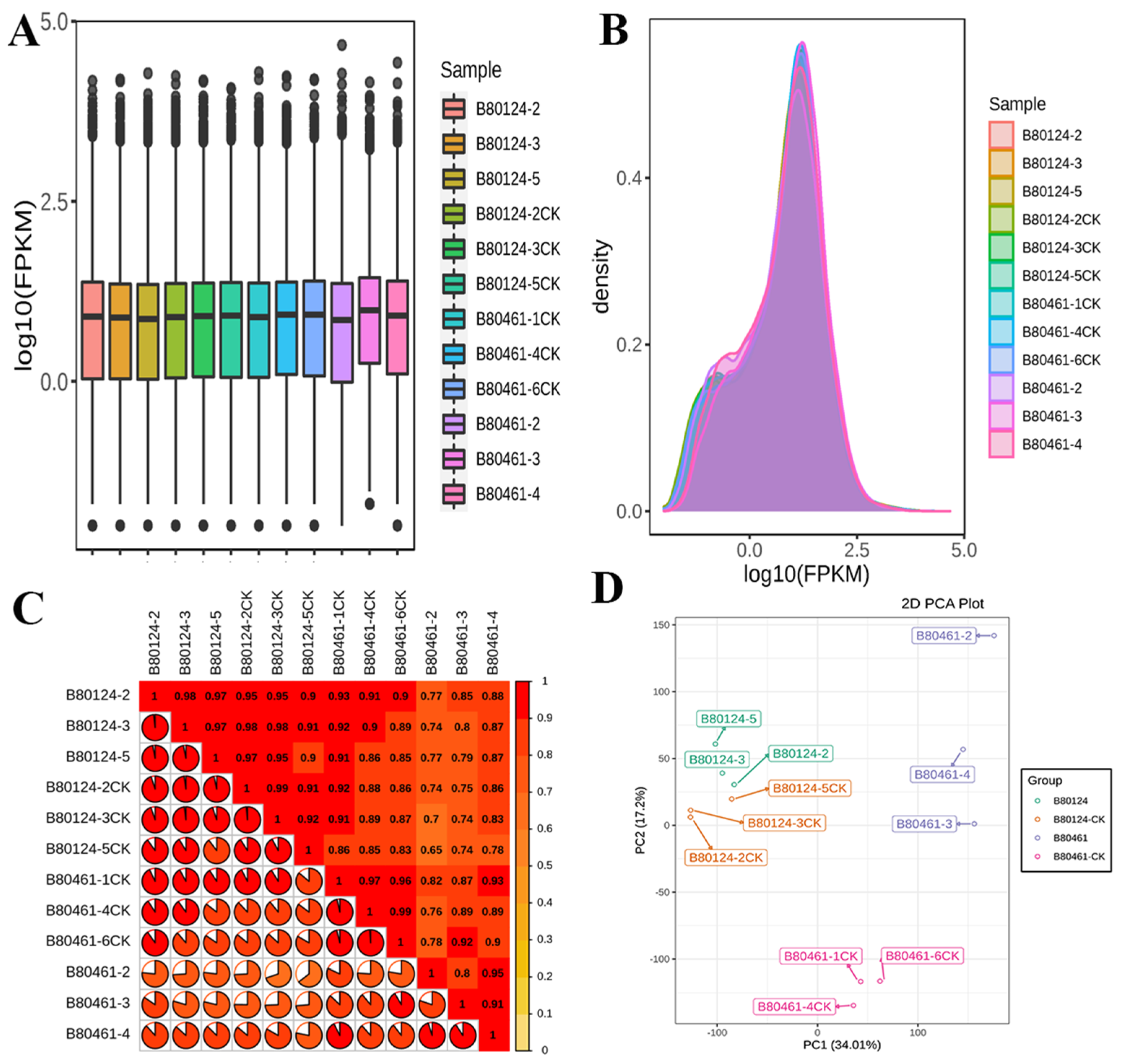
3.2.1. The Volatile Organic Compound Data Quality
3.2.2. VOC Data Analysis
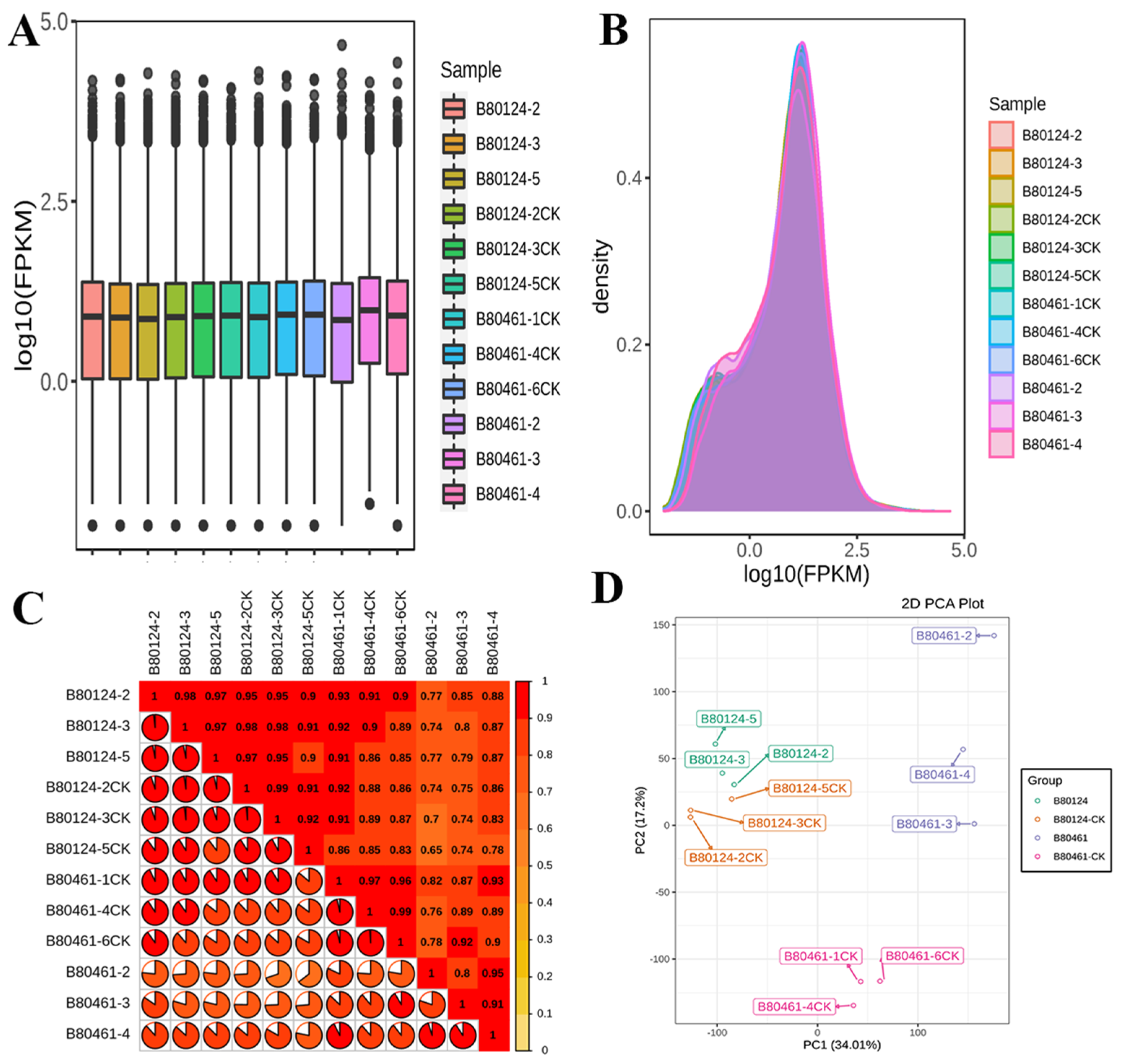
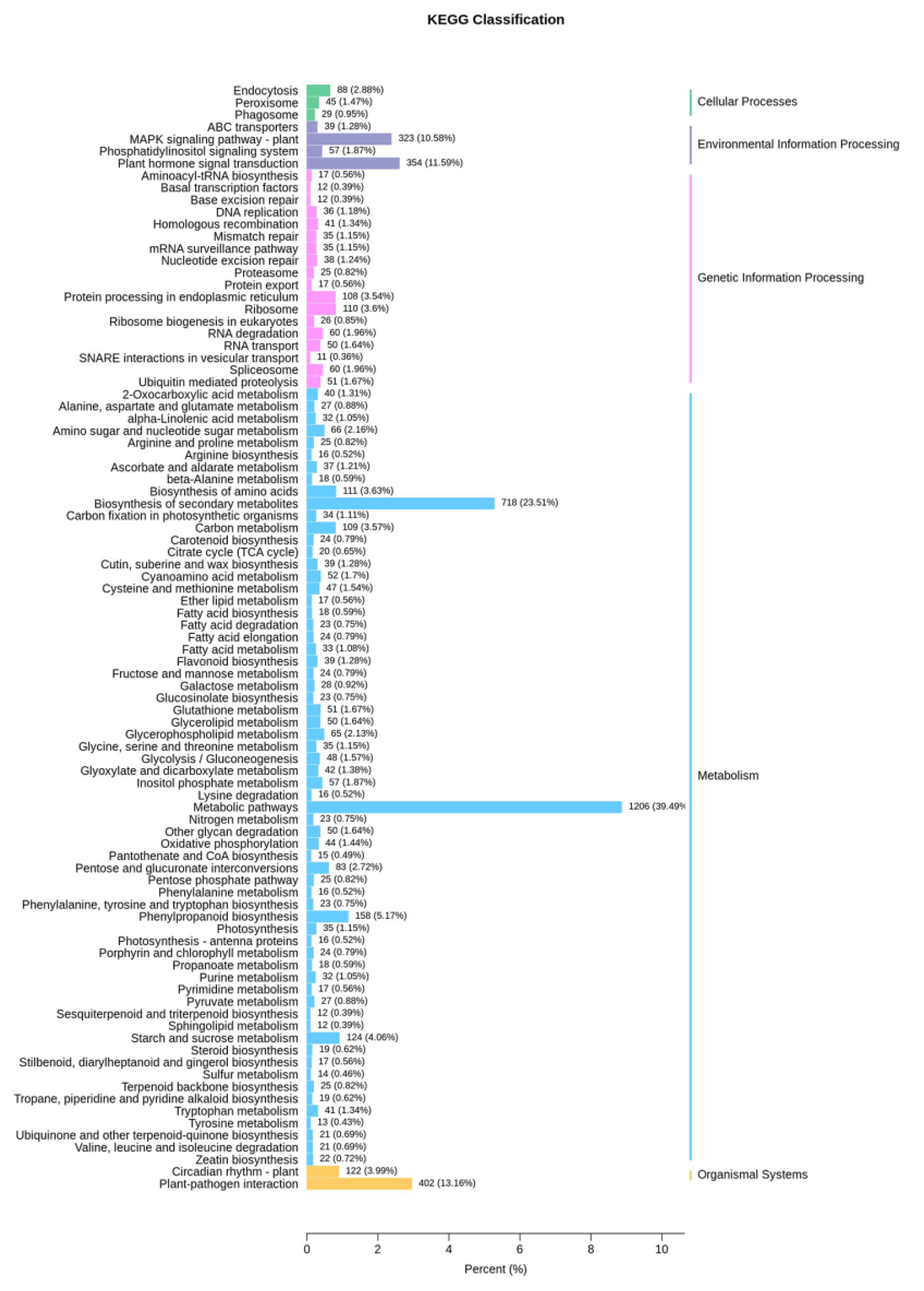
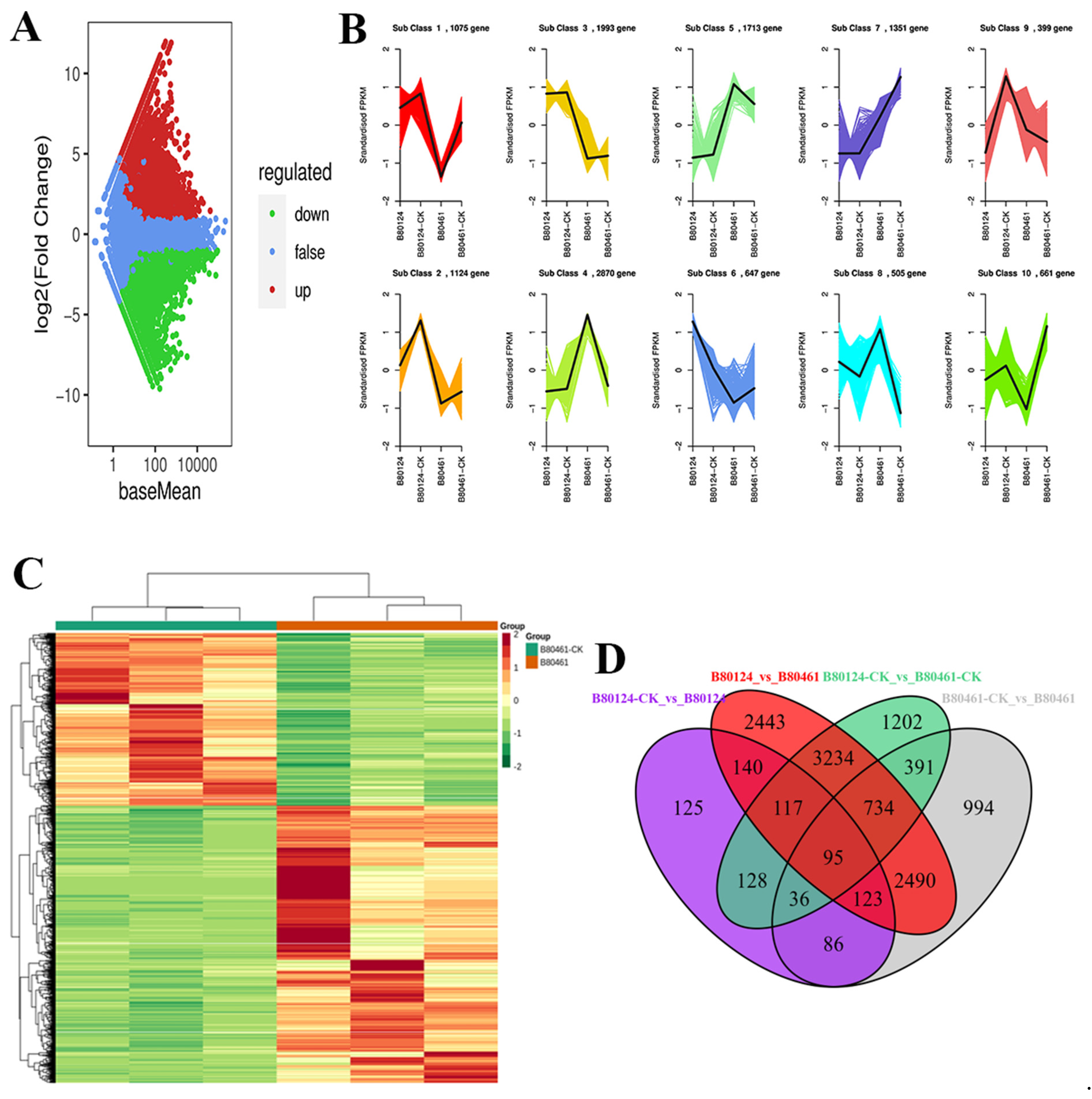
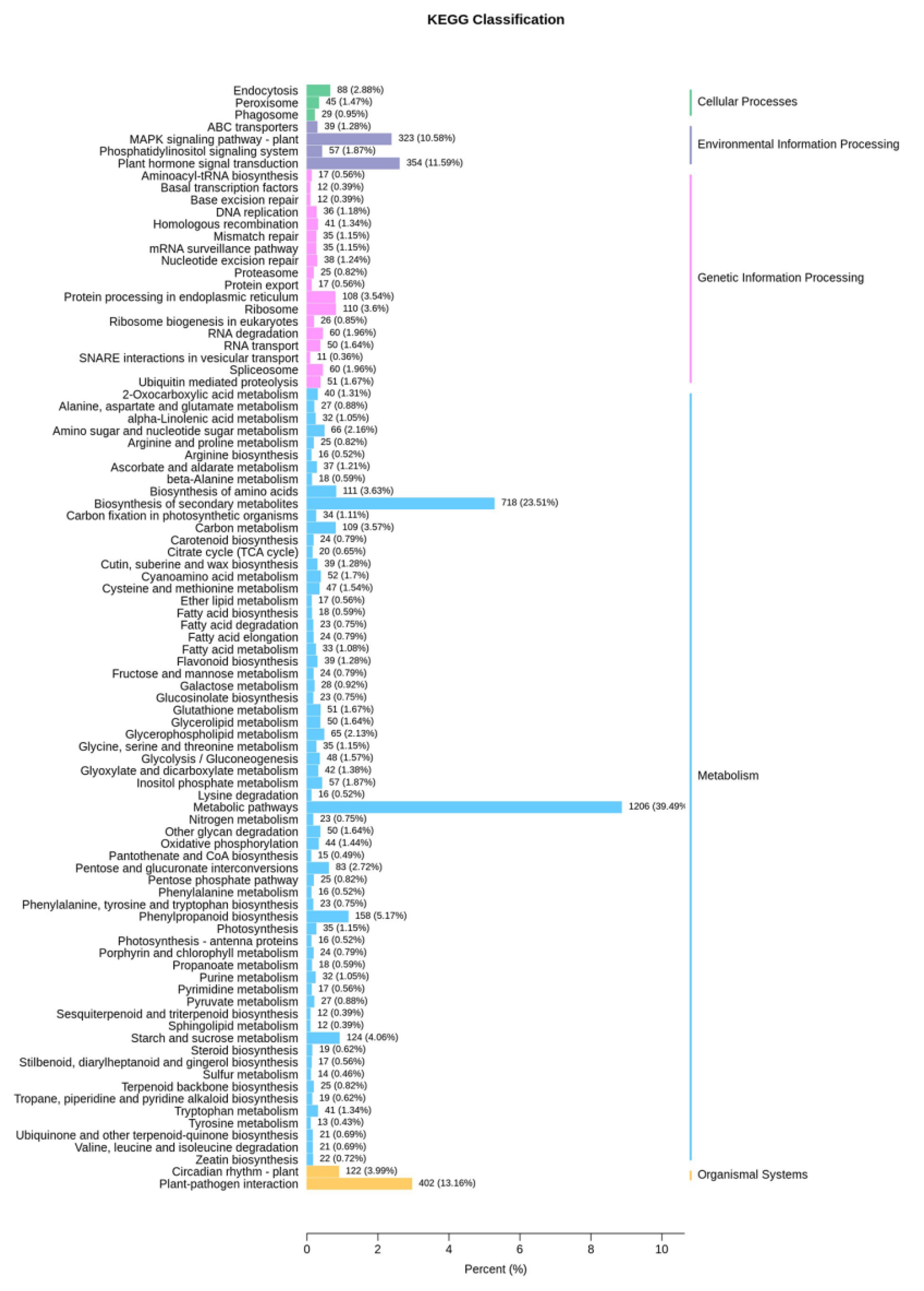
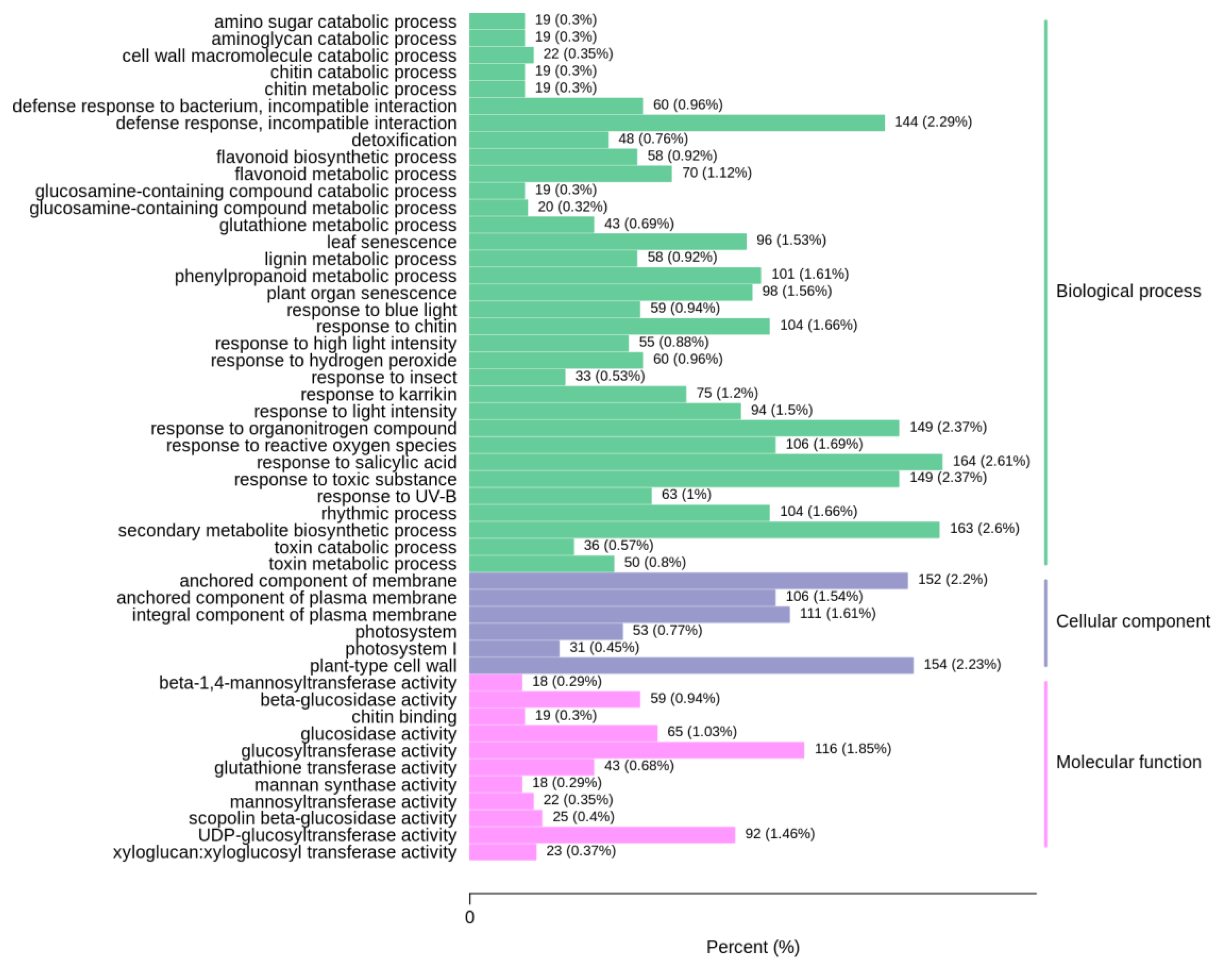
3.3. Transcriptome Analysis of Volatile Organic Compound Accumulation in B. rapa
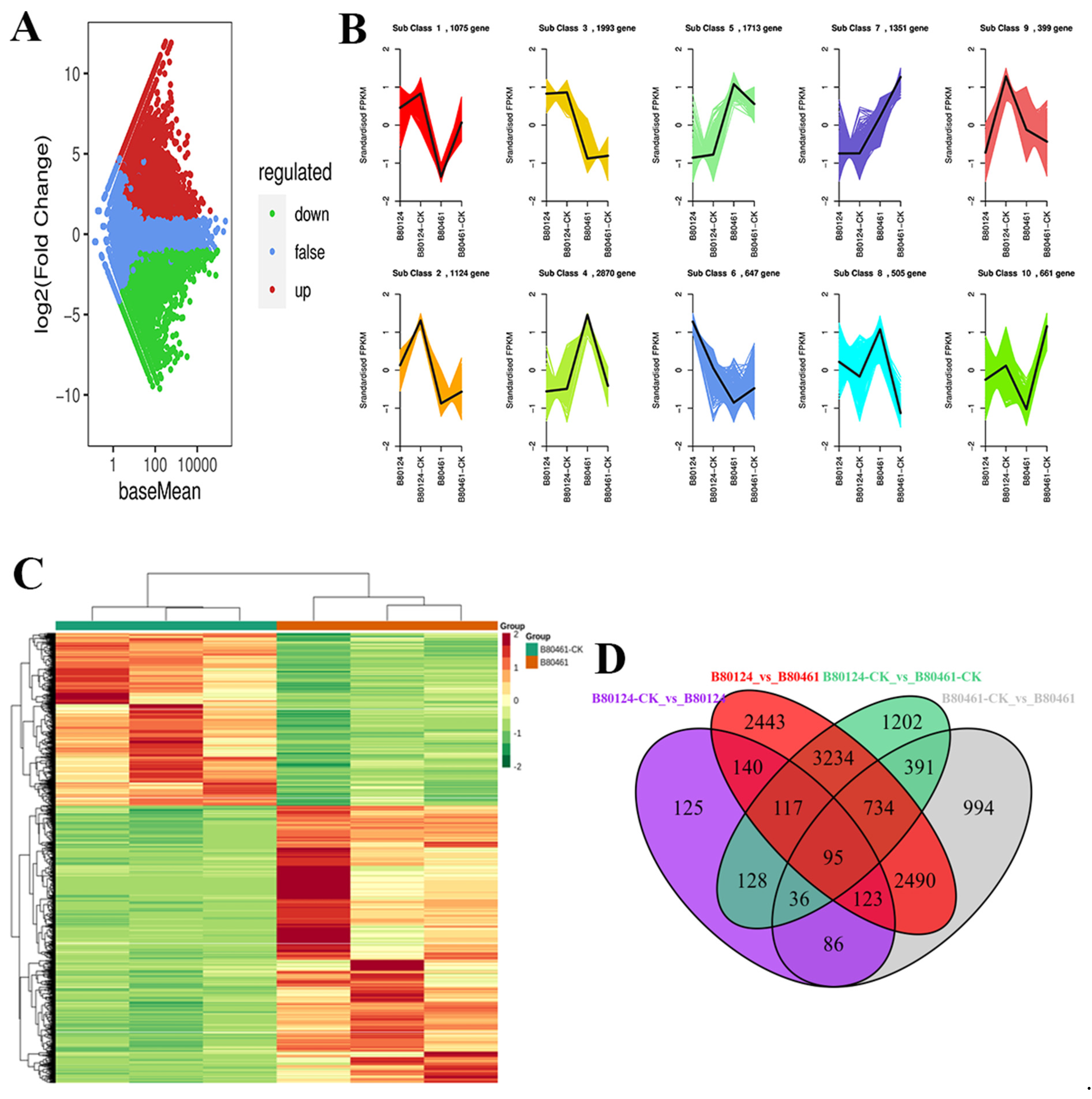
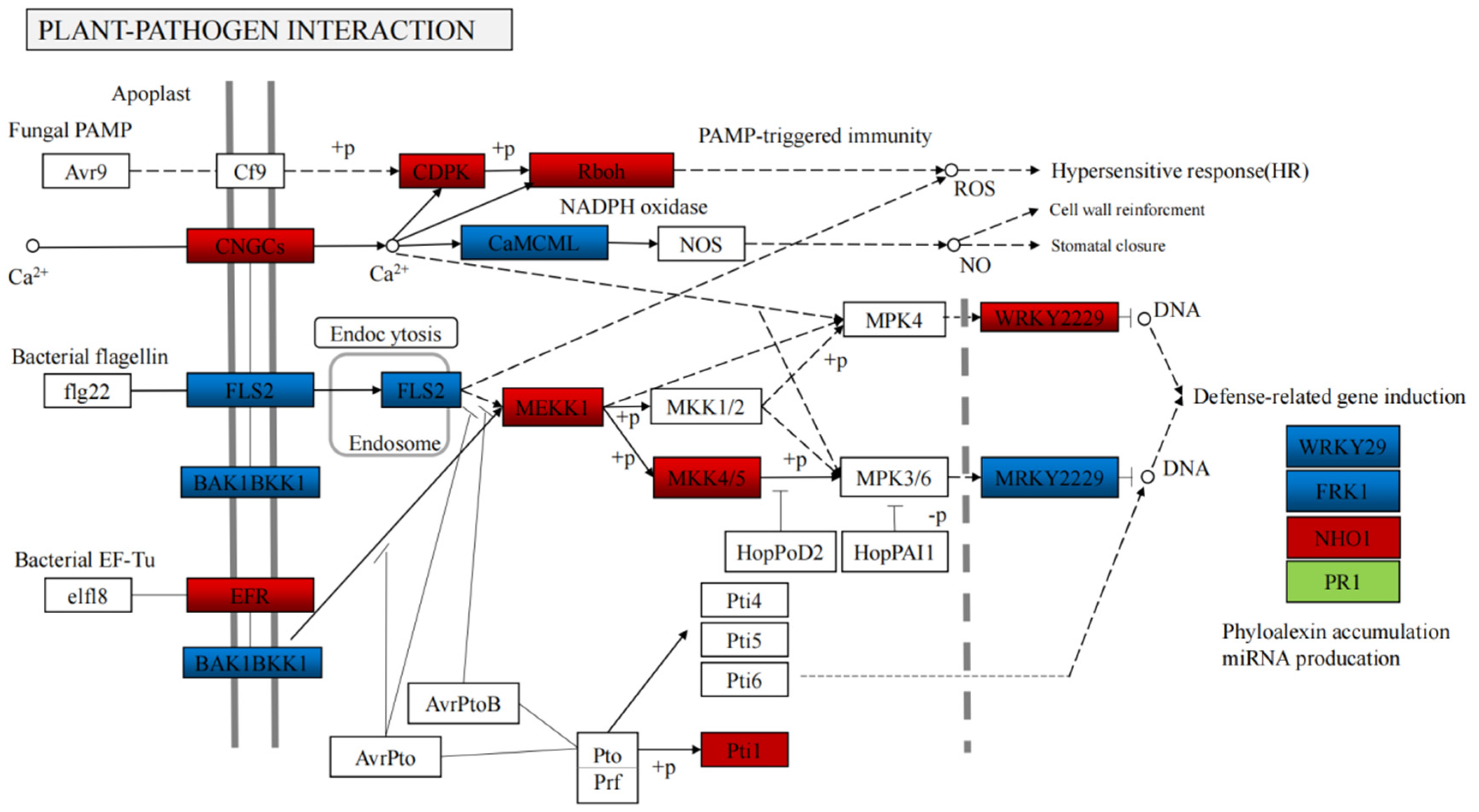
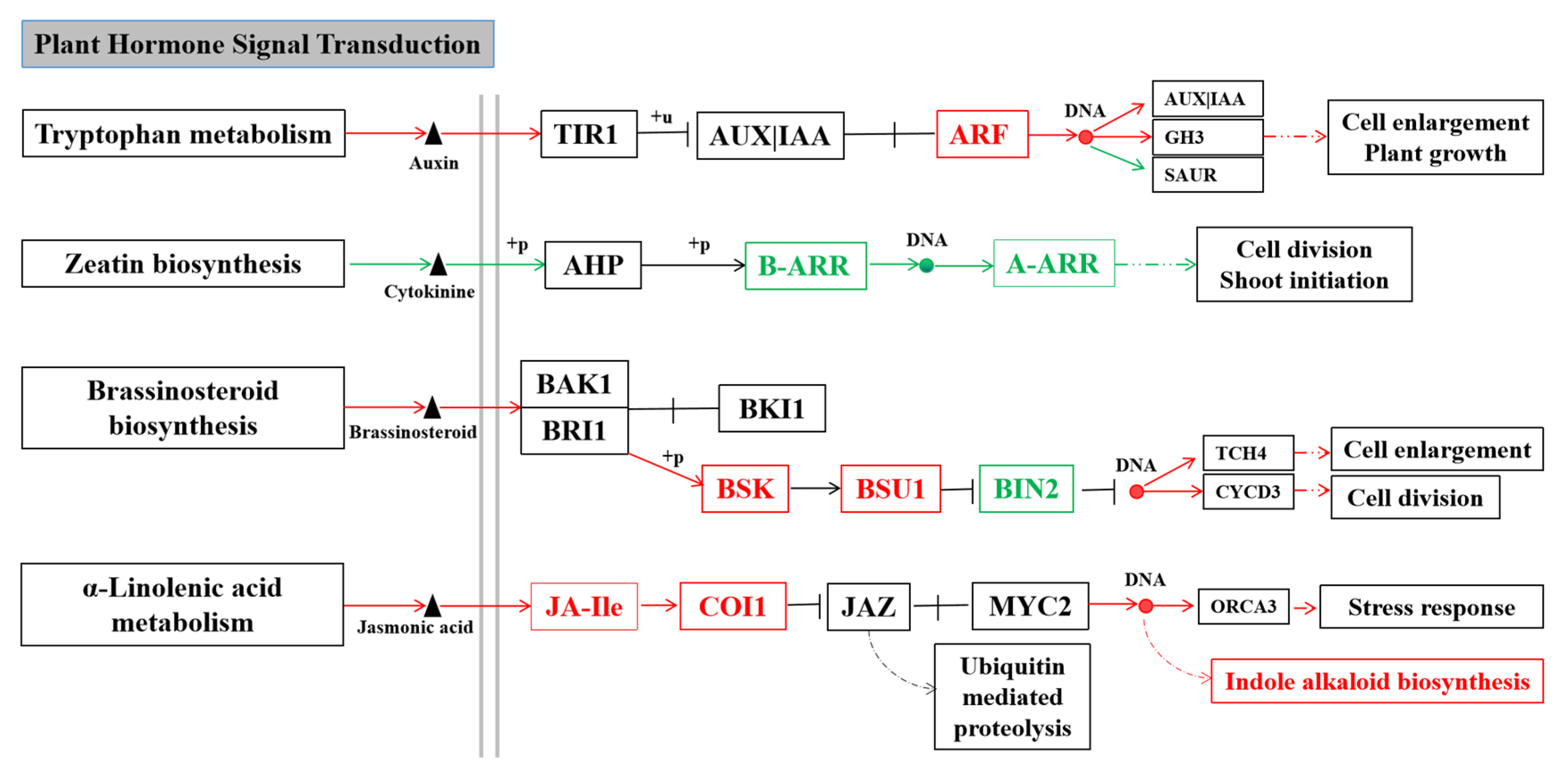
3.4. Integrated Volatile Metabolome and Transcriptome Analysis of VOC Accumulation in B. rapa
4. Discussion
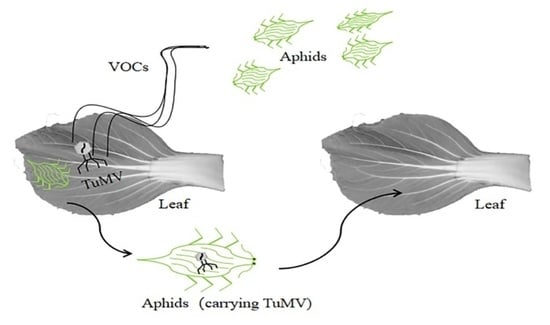
4.1. VOCs Changed Greatly between B80124 and B80461 Inoculated with TuMV
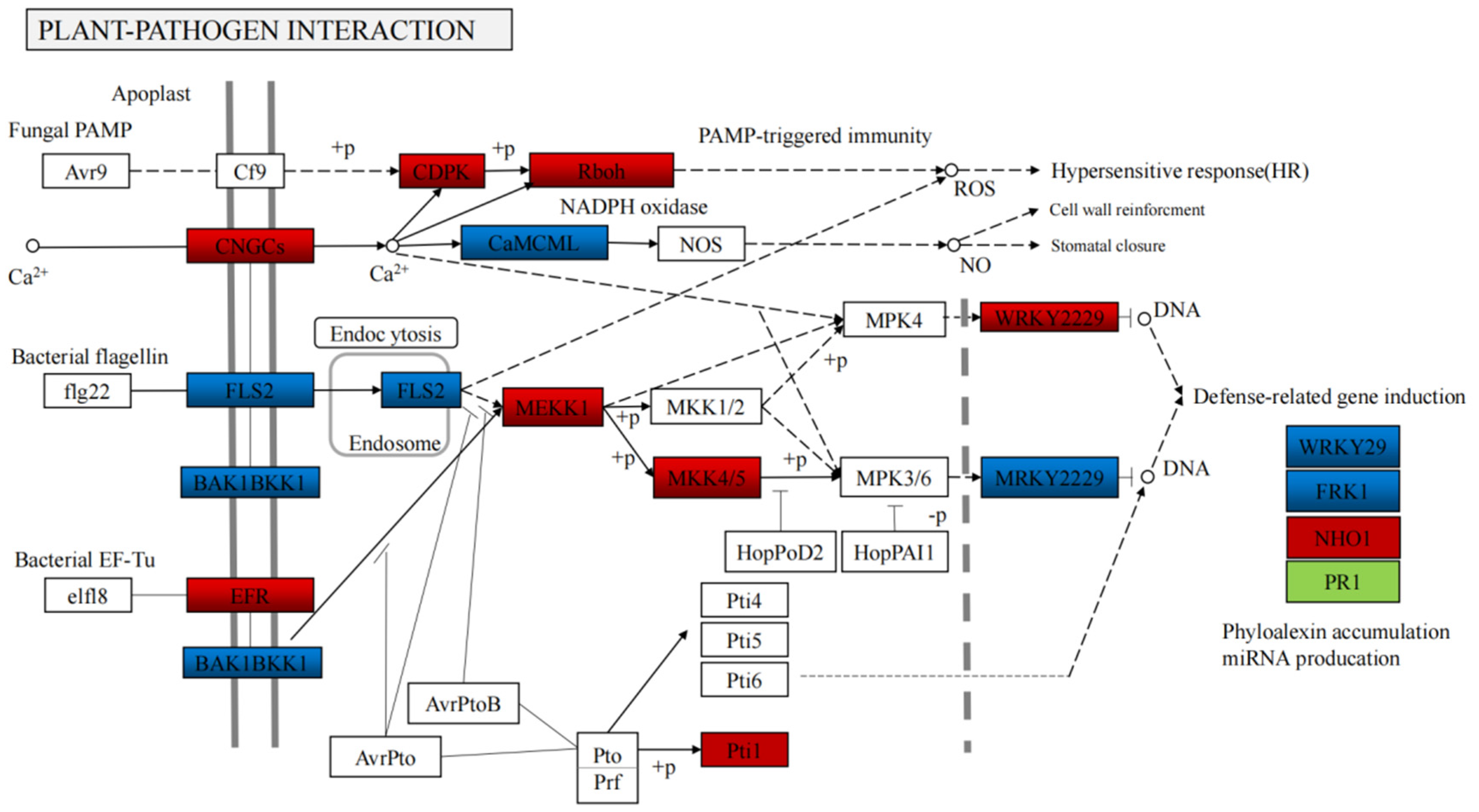
4.2. Integrated Volatile Metabolome and Transcriptome analysis of VOC accumulation in B. rapa
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tomlinson, J.A. Epidemiology and control of virus diseases of vegetables. Ann. Appl. Biol. 1987, 110, 661–681. [Google Scholar] [CrossRef]
- Walkey, D.G.A.; Pink, D.A.C. Reactions of white cabbage (Brassica oleracea var. capitata) to four different strains of turnip mosaic virus. Ann. Appl. Biol. 1988, 112, 273–284. [Google Scholar] [CrossRef]
- Walsh, J.A.; Jenner, C.E. Turnip mosaic virus and the quest for durable resistance. Mol. Plant Pathol. 2002, 3, 289–300. [Google Scholar] [CrossRef]
- Walsh, J.A. Turnip mosaic virus. In Data Sheet for Commonwealth Agriculture Bureau International Global Crop Protection Compendium; CAB International: Wallingford, UK, 1997. [Google Scholar]
- Shattuck, V.I.; Stobbs, L.W. Evaluation of rutabaga cultivars for turnip mosaic virus resistance and the inheritance of resistance. Hortscience. 1987, 22, 935–937. [Google Scholar]
- Hardwick, N.V.; Davies, J.M.L.; Wright, D.M. The incidence of three virus diseases of winter oilseed rape in England and Wales in the 1991/92 and 1992/93 growing seasons. Plant Pathol. 1994, 43, 1045–1049. [Google Scholar] [CrossRef]
- Spence, N.J.; Phiri, N.A.; Hughes, S.L.; Mwaniki, A.; Simons, S.; Oduor, G.; Chacha, D.; Kuria, A.; Ndirangu, S.; Kibata, G.N.; et al. Economic impact of Turnip mosaic virus, Cauliflower mosaic virus and Beet mosaic virus in three Kenyan vegetables. Plant Pathol. 2007, 56, 317–323. [Google Scholar] [CrossRef]
- Casteel, C.L.; Hansen, A.K. Evaluating Insect-Microbiomes at the Plant-Insect Interface. J. Chem. Ecol. 2014, 40, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Antolinez, C.A.; Fereres, A.; Moreno, A. Risk assessment of ‘Candidatus Liberibacter solanacearum’ transmission by the psyllids Bactericera trigonica and B-tremblayi from Apiaceae crops to potato. Sci Rep. 2017, 7, 1–10. [Google Scholar]
- Webster, B. The role of olfaction in aphid host location. Physiol. Entomol. 2012, 37, 10–18. [Google Scholar] [CrossRef]
- Davis, T.S.; Wu, Y.; Eigenbrode, S.D. The effects of bean leafroll virus on life history traits and host selection behavior of specialized pea aphid (Acyrthosiphon pisum, Hemiptera: Aphididae) genotypes. Environ. Entomol. 2017, 46, 68–74. [Google Scholar] [PubMed]
- Fereres, A.; Penaflor, M.F.G.V.; Favaro, C.F.; Azevedo, K.E.X.; Landi, C.H.; Maluta, N.K.P.; Bento, J.M.S.; Lopes, J.R.S. Tomato infection by whitefly-transmitted circulative and non-circulative viruses induce contrasting changes in plant volatiles and vector behaviour. Viruses-Basel 2016, 8, 225. [Google Scholar] [CrossRef] [Green Version]
- Read, D.P.; Feeny, P.P.; Root, R.B. Habitat selection by the aphid parasite Diaeretiella rapae (Hymenoptera: Braconidae) and hyperparasite Charips brassicae (Hymenoptera: Cynipidae). Can. Entomol. 1970, 102, 1567–1578. [Google Scholar] [CrossRef]
- Li, R.; Weldegergis, B.T.; Li, J.; Jung, C.; Qu, J.; Sun, Y.; Qian, H.; Tee, C.; Van Loon, J.J.A.; Dicke, M.; et al. Virulence factors of geminivirus interact with MYC2 to subvert plant resistance and promote vector performance. Plant Cell. 2014, 26, 4991–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, J.A.; Sharpe, A.G.; Jenner, C.E.; Lydiate, D.J. Characterisation of resistance to turnip mosaic virus in oilseed rape (Brassica napus) and genetic mapping of TuRB01. Theor. Appl. Genet. 1999, 99, 1149–1154. [Google Scholar] [CrossRef]
- Li, G.L.; Lv, H.H.; Zhang, S.J.; Zhang, S.F.; Li, F.; Zhang, H.; Qian, W.; Fang, Z.Y.; Sun, R. TuMV management for brassica crops through host resistance: Retrospect and prospects. Plant Pathol. 2019, 68, 1035–1044. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Committee. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press: Beijin, China, 2015; p. 105. (In Chinese)
- Guo, H.J.; Gu, L.; Liu, F.; Chen, F.; Ge, F.; Sun, Y. Aphid-borne viral spread is enhanced by virus-induced accumulation of plant reactive oxygen species. Plant Physiol. 2019, 179, 143–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alazem, M.; Lin, N.-S. Roles of plant hormones in the regulation of host-virus interactions. Mol. Plant Pathol. 2015, 16, 529–540. [Google Scholar] [CrossRef]
- Collum, T.D.; Culver, J.N. The impact of phytohormones on virus infection and disease. Curr. Opin. Virol. 2016, 17, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone crosstalk in plant disease and defense: More than just JASMONATE-SALICYLATE antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef]
- Derksen, H.; Rampitsch, C.; Daayf, F. Signaling cross-talk in plant disease resistance. Plant Sci. 2013, 207, 79–87. [Google Scholar] [CrossRef]
- Zarate, S.I.; Kempema, L.A.; Walling, L.L. Silverleaf whitefly induces salicylic acid defenses and suppresses effectual jasmonic acid defenses. Plant Physiol. 2007, 143, 866–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broin, M.; Cuine, S.; Eymery, F.; Rey, P. The plastidic 2-cysteine peroxiredoxin is a target for a thioredoxin involved in the protection of the photosynthetic apparatus against oxidative damage. Plant Cell. 2002, 14, 1417–1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, E.S. A transmissible mosaic disease of Chinese cabbage, mustard and Turnip. Afr. J. Agric. Res. 1921, 22, 173–177. [Google Scholar]
- Smith, K.M. A virus disease of cultivated Crucifers. Ann. Appl. Biol. 1935, 22, 239–242. [Google Scholar] [CrossRef]
- Ling, L.; Yang, J.Y. A mosaic disease of Rape and other crucifera in China. Phytopathology. 1940, 30, 338–342. [Google Scholar]
- Walsh, J.A.; Rusholme, R.L.; Hughes, S.L.; Jenner, C.E.; Bambridge, J.M.; Lydiate, D.J.; Green, S.K. Different classes of resistance to turnip mosaic virus in Brassica rapa. Eur. J. Plant Pathol. 2002, 108, 15–20. [Google Scholar] [CrossRef]
- D’Alessandro, M.; Turlings, T.C.J. Advances and challenges in the identification of volatiles that mediate interactions among plants and arthropods. Analyst 2006, 131, 24–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meiners, T.; Hilker, M. Induction of plant synomones by oviposition of a phytophagous insect. J. Chem. Ecol. 2000, 26, 221–232. [Google Scholar] [CrossRef]
- Bleeker, P.M.; Diergaarde, P.J.; Ament, K.; Guerra, J.; Weidner, M.; Schuetz, S.; de Both, M.T.J.; Haring, M.A.; Schuurink, R.C. The role of specific tomato volatiles in tomato-whitefly interaction. Plant Physiol. 2009, 151, 925–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, J.-B.; Yao, D.-M.; Zhang, T.; Walling, L.L.; Yang, M.; Wang, Y.-J.; Liu, S.-S. Suppression of terpenoid synthesis in plants by a virus promotes its mutualism with vectors. Ecol. Lett. 2013, 16, 390–398. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; Bosque-Perez, N.A.; Davis, T.S. Insect-borne plant pathogens and their vectors: Ecology, evolution, and complex interactions. Annu. Rev. Entomol. 2018, 63, 169–191. [Google Scholar] [CrossRef]
- Mauck, K.E. Variation in virus effects on host plant phenotypes and insect vector behavior: What can it teach us about virus evolution? Curr. Opin. Virol. 2016, 21, 114–123. [Google Scholar] [CrossRef]
- Furlong, M.J.; Pell, J.K. Interactions between the fungal entomopathogen Zoophthora radicans Brefeld (Entomophthorales) and two hymenopteran parasitoids attacking the diamondback moth, Plutella xylostella L. J. Invertebr. Pathol. 1996, 68, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, U.; Loreto, F.; Reichstein, M. Physiological and physicochemical controls on foliar volatile organic compound emissions. Trends Plant Sci. 2004, 9, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Melotto, M.; Underwood, W.; He, S.Y. Role of stomata in plant innate immunity and foliar bacterial diseases. Annu. Rev. Phytopathol. 2008, 46, 101–122. [Google Scholar] [CrossRef] [Green Version]
- Ameye, M.; Allmann, S.; Verwaeren, J.; Smagghe, G.; Haesaert, G.; Schuurink, R.C.; Audenaert, K. Green leaf volatile production by plants: A meta-analysis. New Phytol. 2018, 220, 666–683. [Google Scholar] [CrossRef]
- Lin, P.A.; Chen, Y.; Ponce, G.; Acevedo, F.E.; Lynch, J.P.; Anderson, C.T.; Ali, J.G.; Felton, G.W. Stomata-mediated interactions between plants, herbivores, and the environment. Trends Plant Sci. 2021. [Google Scholar] [CrossRef]
- Seidl-Adams, I.; Richter, A.; Boomer, K.B.; Yoshinaga, N.; Degenhardt, J.; Tumlinson, J.H. Emission of herbivore elicitor-induced sesquiterpenes is regulated by stomatal aperture in maize (Zea mays) seedlings. Plant Cell Environ. 2015, 38, 23–34. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.; Zhang, L.; Huang, W.; Zhang, S.; Zhang, S.; Li, F.; Zhang, H.; Sun, R.; Zhao, J.; Li, G. Integrated Volatile Metabolomics and Transcriptomics Analyses Reveal the Influence of Infection TuMV to Volatile Organic Compounds in Brassica rapa. Horticulturae 2022, 8, 57. https://doi.org/10.3390/horticulturae8010057
Lu X, Zhang L, Huang W, Zhang S, Zhang S, Li F, Zhang H, Sun R, Zhao J, Li G. Integrated Volatile Metabolomics and Transcriptomics Analyses Reveal the Influence of Infection TuMV to Volatile Organic Compounds in Brassica rapa. Horticulturae. 2022; 8(1):57. https://doi.org/10.3390/horticulturae8010057
Chicago/Turabian StyleLu, Xinxin, Lei Zhang, Wenyue Huang, Shujiang Zhang, Shifan Zhang, Fei Li, Hui Zhang, Rifei Sun, Jianjun Zhao, and Guoliang Li. 2022. "Integrated Volatile Metabolomics and Transcriptomics Analyses Reveal the Influence of Infection TuMV to Volatile Organic Compounds in Brassica rapa" Horticulturae 8, no. 1: 57. https://doi.org/10.3390/horticulturae8010057
APA StyleLu, X., Zhang, L., Huang, W., Zhang, S., Zhang, S., Li, F., Zhang, H., Sun, R., Zhao, J., & Li, G. (2022). Integrated Volatile Metabolomics and Transcriptomics Analyses Reveal the Influence of Infection TuMV to Volatile Organic Compounds in Brassica rapa. Horticulturae, 8(1), 57. https://doi.org/10.3390/horticulturae8010057