
Embryo Culture, In Vitro Propagation, and Molecular Identification for Advanced Olive Breeding Programs

 ,
,  ,
,  ,
,
Abstract
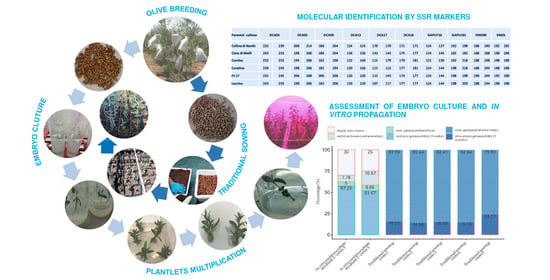
:
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction and Microsatellite Analysis
2.3. Controlled Crossings
2.4. Traditional Sowing
2.5. In Vitro Embryo Culture
2.6. Media Composition
2.7. Statistical Analysis
3. Results
3.1. SSR Markers Analysis
3.2. Traditional Sowing
3.3. In Vitro Embryo Culture
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Rienzo, V.; Sion, S.; Taranto, F.; D’Agostino, N.; Montemurro, C.; Fanelli, V.; Sabetta, W.; Boucheffa, S.; Tamendjari, A.; Pasqualone, A.; et al. Genetic flow among olive populations within the Mediterranean basin. PeerJ 2018, 6, e5260. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, G.; Prevost, G.; Messeri, C.; Carignani, G. Olive Germplasm: Cultivars and World-Wide Collections; FAO: Roma, Italy, 1998. [Google Scholar]
- Loreti, F.; Guerriero, R.; Triolo, E.; Vitagliano, C.; Anna Pisa, S. Proposal of a method for clonal and healt selection in olive cultivation. Acta Hortic. 1994, 356, 82–86. [Google Scholar] [CrossRef]
- Gregoriou, C. Assessment of Variation of Landraces of Olive Tree in Cyprus. Euphytica 1996, 87, 173–176. [Google Scholar] [CrossRef]
- Saponari, M.; Nigro, F.; Vovlas, N.; Cariddi, C.; Grieco, F.; Trisciuzzi, N.; Savino, V.; Martelli, G.P. Punti critici dell’olivo. In Proceedings of the Incontro Divulgativo POM A32-Risultati di due Anni di Attività, Termoli-Campobassol, Italy, 1–2 March 2001; pp. 311–324. [Google Scholar]
- Rugini, E.; Gutiérrez-Pesce, P. Genetic improvement of olive. Pomol. Croat. 2006, 12, 43–72. [Google Scholar]
- Fontanazza, G.; Baldoni, L. Proposed programme for the genetic improvement of the olive. Olivae 1990, 34, 32–40. [Google Scholar]
- Baù, A.; Delbianco, A.; Stancanelli, G.; Tramontini, S. Susceptibility of Olea europaea L. varieties to Xylella fastidiosa subsp. pauca ST53: Systematic literature search up to 24 March 2017. EFSA J. 2017, 15, e04772. [Google Scholar]
- Boscia, D.; Altamura, G.; Ciniero, A.; di Carolo, M.; Dongiovanni, C.; Fumarola, G.; Giampetruzzi, A.; Greco, P.; la Notte, P.; Loconsole, G. Resistance to Xylella fastidiosa in different olive cultivars. Inf. Agrar. 2017, 73, 59–63. [Google Scholar]
- Pavan, S.; Vergine, M.; Nicolì, F.; Sabella, E.; Aprile, A.; Negro, C.; Fanelli, V. Screening of Olive Biodiversity Defines Genotypes Potentially Resistant to Xylella fastidiosa. Front. Plant Sci. 2021, 12, 1734. [Google Scholar] [CrossRef] [PubMed]
- Sotomayor-León, E.M.; Caballero, J.M. An easy method of breaking olive stones to remove mechanical dormancy. Acta Hortic. 1990, 286, 113–116. [Google Scholar] [CrossRef]
- Bellini, E. Genetic Improvement of Olive from Clonal Selection to Cross-Breeding Programs. Adv. Hortic. Sci. 2008, 22, 1000–1014. [Google Scholar] [CrossRef]
- Rallo, L.; El Riachy, M.; Rallo, P. The Time and Place for Fruit Quality in Olive Breeding. In Breeding for Fruit Quality; Jenks, M.A., Bebeli, P.J., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011; pp. 323–347. [Google Scholar] [CrossRef]
- Scaramuzzi, F. Influenza dell’epoca racolta dei frutti sulla germinabilità dei semi di olivo. Agric. Ital. 1958, 58, 387–402. [Google Scholar]
- Crisosto, C.; Sutter, E.G. Improving Manzanillö olive seed germination. Improv. Manzanillö Olive Seed Germination 1985, 20, 100–102. [Google Scholar]
- Cañas, L.A.; Carramolino, L.; Vicente, M. Vegetative Propagation of the Olive Tree from in Vitro Cultured Embryos. Plant Sci. 1987, 50, 85–90. [Google Scholar] [CrossRef]
- Rinaldi, L.M.R.; Lambardi, M. In vitro germinability and ethylene biosynthesis in cytokinin-treated olive seeds (Olea europaea L.). Adv. Hortic. Sci. 1998, 12, 59–62. [Google Scholar]
- Acebedo, M.M.; Lavee, S.; Liñán, J.; Troncoso, A. In Vitro Germination of Embryos for Speeding up Seedling Development in Olive Breeding Programmes. Sci. Hortic. 1997, 69, 207–215. [Google Scholar] [CrossRef]
- García, J.L.; Troncoso, J.; Sarmiento, R.; Troncoso, A. Influence of Carbon Source and Concentration on the in Vitro Development of Olive Zygotic Embryos and Explants Raised from Them. Plant Cell Tissue Organ. Cult. 2002, 69, 95–100. [Google Scholar] [CrossRef]
- Sharma, D.R.; Kaur, R.; Kumar, K. Embryo Rescue in Plants—A Review. Euphytica 1996, 89, 325–337. [Google Scholar] [CrossRef]
- Zulkarnain, Z.; Tapingkae, T.; Taji, A. Applications of In Vitro Techniques in Plant Breeding. In Advances in Plant Breeding Strategies: Breeding, Biotechnology and Molecular Tools; Springer: Cham, Switzerland, 2015; pp. 293–328. [Google Scholar] [CrossRef]
- Raghavan, V. One Hundred Years of Zygotic Embryo Culture Investigations. Vitro Cell. Dev. Biol. Plant 2003, 39, 437–442. [Google Scholar] [CrossRef]
- Rai, M.K.; Kalia, R.K.; Singh, R.; Gangola, M.P.; Dhawan, A.K. Developing Stress Tolerant Plants through in Vitro Selection—An Overview of the Recent Progress. Environ. Exp. Bot. 2011, 71, 89–98. [Google Scholar] [CrossRef]
- Pasqualone, A.; Montemurro, C.; Di Rienzo, V.; Summo, C.; Paradiso, V.M.; Caponio, F. Evolution and perspectives of cultivar identification and traceability from tree to oil and table olives by means of DNA markers. J. Sci. Food Agric. 2016, 96, 3642–3657. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, V.; Miazzi, M.M.; Fanelli, V.; Sabetta, W.; Montemurro, C. The preservation and characterization of Apulian olive germplasm biodiversity. Acta Hortic. 2018, 1199, 1–6. [Google Scholar] [CrossRef]
- Spadoni, A.; Sion, S.; Gadaleta, S.; Savoia, M.; Piarulli, L.; Fanelli, V.; Di Rienzo, V.; Taranto, F.; Miazzi, M.M.; Montemurro, C. A simple and rapid method for genomic DNA extraction and microsatellite analysis in tree plants. J. Agric. Sci. Technol. 2019, 21, 1215–1226. [Google Scholar]
- Alba, V.; Sabetta, W.; Blanco, A.; Pasqualone, A.; Montemurro, C. Microsatellite markers to identify specific alleles in DNA extracted from monovarietal virgin olive oils. Eur. Food Res. Technol. 2009, 229, 375–382. [Google Scholar] [CrossRef]
- Miazzi, M.M.; di Rienzo, V.; Mascio, I.; Montemurro, C.; Sion, S.; Sabetta, W.; Vivaldi, G.A.; Camposeo, S.; Caponio, F.; Squeo, G.; et al. Re.Ger.O.P.: An Integrated Project for the Recovery of Ancient and Rare Olive Germplasm. Front. Plant Sci. 2020, 11, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montemurro, C.; Dambruoso, G.; Bottalico, G.; Sabetta, W. Self-Incompatibility Assessment of Some Italian Olive Geno-types (Olea europaea L.) and Cross-Derived Seedling Selection by SSR Markers on Seed Endosperms. Front. Plant Sci. 2019, 10, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voyiatzis, D.G.; Pritsa, T. The onset and disappearance of relative dormancy of olive embryos as affected by age. Acta Hortic. 1994, 356, 148–151. [Google Scholar] [CrossRef]
- Liñán, J.; Troncoso, A.; Rapoport, H.F. Olive Embryo development stage and the possibility of obtaining viable seedlings. Acta Hortic. 1999, 474, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Kiani, M.; Zamani, Z.; Ebadi, A. In vitro germination of three olive cultivars. Acta Hortic. 2006, 725, 333–336. [Google Scholar] [CrossRef]
- Germanà, M.A.; Chiancone, B.; Hammami, S.B.M.; Rapoport, H.F. Olive Embryo in Vitro Germination Potential: Role of Explant Configuration and Embryo Structure among Cultivars. Plant Cell Tissue Organ. Cult. 2014, 118, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Rugini, E. In Vitro Propagation of Some Olive (Olea Europaea Sativa, L.) Cultivars with Different Root-Ability, and Medium Development Using Analytical Data from Developing Shoots and Embryos. Sci. Hortic. 1984, 24, 123–134. [Google Scholar] [CrossRef]
- Murashige, T.; Folke, S. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Nitsch, J.P.; Nitsch, C. Haploid plants from pollen grains. Science 1969, 163, 85–87. [Google Scholar] [CrossRef] [PubMed]
- Leva, A. Innovative Protocol for “Ex Vitro Rooting” on Olive Micropropagation. Open Life Sci. 2011, 6, 352–358. [Google Scholar] [CrossRef]
- Zienkiewicz, A.; Jiménez-López, J.C.; Zienkiewicz, K.; de Dios Alché, J.; Rodríguez-García, M.I. Development of the Cotyledon Cells during Olive (Olea Europaea L.) in Vitro Seed Germination and Seedling Growth. Protoplasma 2011, 248, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Brhadda, N.; Walali Loudyi, D.M.; Abousalim, A.; Benali, D. Effect of temperature and endosperm on dormancy and germination of olive embryos (Moroccan Picholine variety). Agronomie 2000, 20, 643–653. [Google Scholar] [CrossRef] [Green Version]
- Rugini, E.; Gutierrez-Pesce, P.; Spampinato, P.L.; Ciarmiello, A.; D’Ambrosio, C. New perspective for biotechnologies in olive breeding: Morphogenesis, in vitro selection and gene transformation. Acta Hortic. 1999, 474, 107–110. [Google Scholar] [CrossRef]
- Lambardi, M.; Rugini, E. Micropropagation of olive (Olea europaea L.). In Micropropagation of Woody Trees and Fruits; Springer: Dordrecht, The Netherlands, 2003; pp. 621–646. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parental Cultivar | DCA03 | DCA05 | DCA09 | DCA13 | DCA17 | DCA18 | GAPU71b | GAPU101 | EMO90 | EMOL | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cellina di Nardò | 232 | 239 | 206 | 214 | 186 | 204 | 124 | 124 | 179 | 179 | 171 | 171 | 124 | 127 | 192 | 198 | 188 | 190 | 192 | 192 |
| Cima di Melfi | 243 | 253 | 198 | 206 | 186 | 204 | 156 | 156 | 143 | 143 | 175 | 177 | 124 | 144 | 182 | 182 | 188 | 188 | 198 | 198 |
| Carolea | 232 | 253 | 194 | 206 | 162 | 198 | 122 | 140 | 115 | 179 | 179 | 181 | 121 | 130 | 192 | 218 | 188 | 198 | 198 | 198 |
| Coratina | 239 | 243 | 198 | 206 | 182 | 194 | 120 | 156 | 115 | 115 | 177 | 181 | 124 | 144 | 198 | 218 | 188 | 194 | 198 | 198 |
| FS 17 | 232 | 245 | 206 | 208 | 206 | 206 | 120 | 120 | 115 | 143 | 173 | 177 | 124 | 144 | 198 | 198 | 190 | 194 | 192 | 198 |
| Leccino | 243 | 253 | 198 | 206 | 162 | 206 | 120 | 120 | 107 | 117 | 177 | 177 | 124 | 144 | 198 | 200 | 188 | 194 | 198 | 198 |
| Crossbreed Combination | No. of Stones Directly Sown | No. of Seedlings Emerged after 5 Months | Germinability (%) |
|---|---|---|---|
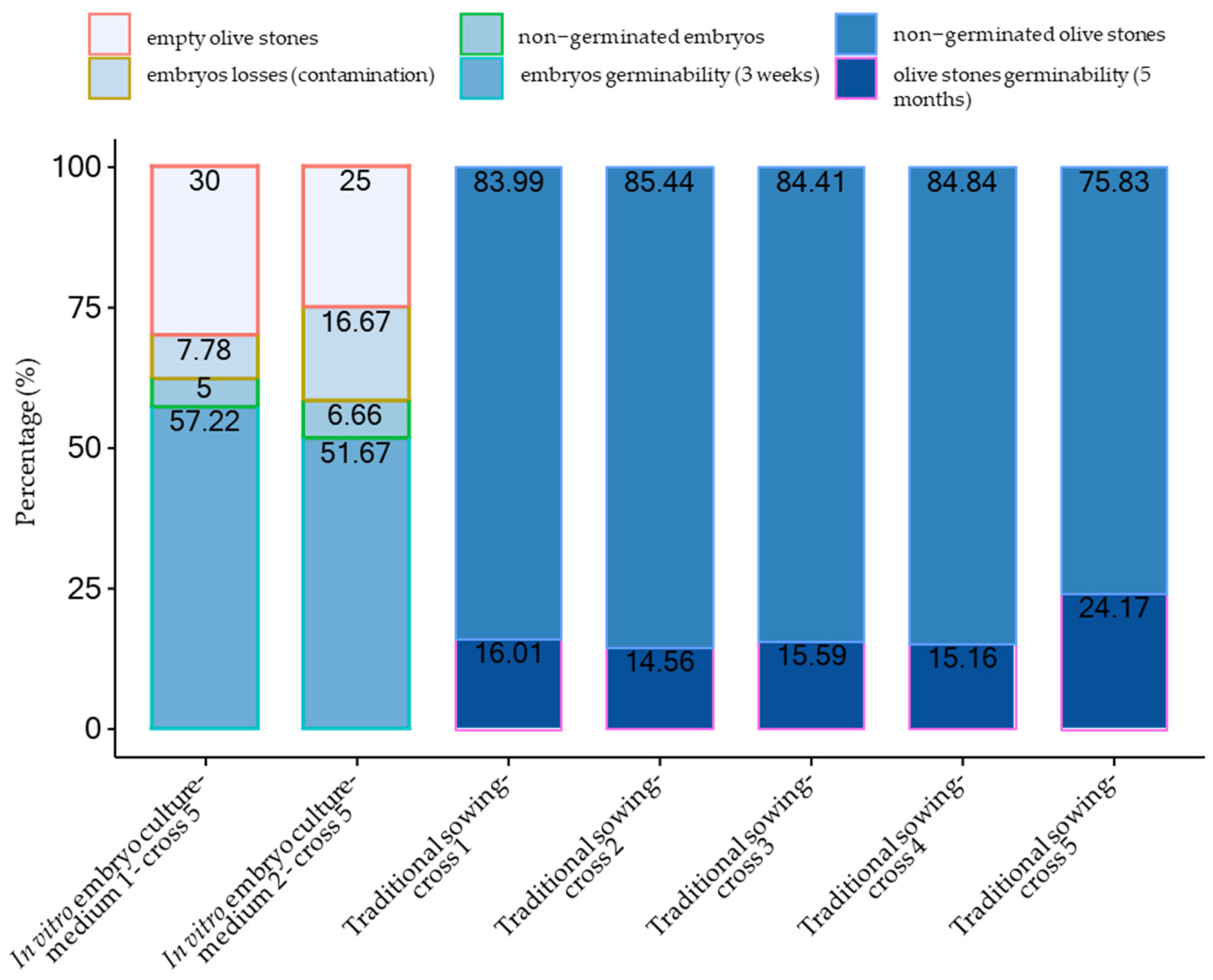
| Combination 1—Cima di Melfi × FS 17 | 818 | 131 | 16.01 |
| Combination 2—Leccino × Cima di Melfi | 364 | 53 | 14.56 |
| Combination 3—Leccino free pollinated | 571 | 89 | 15.59 |
| Combination 4—Leccino × Cellina di Nardò | 686 | 104 | 15.16 |
| Combination 5—Leccino × FS 17 | 1721 | 416 | 24.17 |
| Medium Type | No. of Stones | No. of Embryos Cultured | No. of Embryos Lost Due to Contamination | No. of Ungerminated Embryos | No. of Germinated Embryos | 1 Germinability (%) on the Total Olive stones |
|---|---|---|---|---|---|---|
| Medium 1 | 180 | 126 | 14 | 9 | 103 | 57.22 |
| Medium 2 | 120 | 90 | 20 | 8 | 62 | 51.67 |
| Statistical analysis of the germinability of isolated embryos between the two media tested | ||||||
| no. successfully stabilized embryos | no. of germinated embryos | 1,2 germinability (%) on the successfully stabilized embryos | ||||
| Medium 1 | 112 | 103 | 91.96% | |||
| Medium 2 | 70 | 62 | 88.57% | |||
| χ2 value | 0.25345 | |||||
| p-value | 0.6147 | |||||
| Crossbreed Combination | No. of Stones | No. of Embryos Cultured | No. of Ungerminated Embryos | No. of Germinated Embryos | No. of Embryos Lost Due to Contamination | Germinability (%) on the Total Olive Stones |
|---|---|---|---|---|---|---|
| Combination 6—FS 17 × Coratina | 200 | 143 | 13 | 109 | 21 | 54.5 |
| Combination 7—FS 17 × Carolea | 200 | 138 | 11 | 105 | 22 | 52.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montilon, V.; Susca, L.; Potere, O.; Roseti, V.; Campanale, A.; Saponari, A.; Montemurro, C.; Fanelli, V.; Venerito, P.; Bottalico, G. Embryo Culture, In Vitro Propagation, and Molecular Identification for Advanced Olive Breeding Programs. Horticulturae 2022, 8, 36. https://doi.org/10.3390/horticulturae8010036
Montilon V, Susca L, Potere O, Roseti V, Campanale A, Saponari A, Montemurro C, Fanelli V, Venerito P, Bottalico G. Embryo Culture, In Vitro Propagation, and Molecular Identification for Advanced Olive Breeding Programs. Horticulturae. 2022; 8(1):36. https://doi.org/10.3390/horticulturae8010036
Chicago/Turabian StyleMontilon, Vito, Leonardo Susca, Oriana Potere, Vincenzo Roseti, Antonia Campanale, Antonia Saponari, Cinzia Montemurro, Valentina Fanelli, Pasquale Venerito, and Giovanna Bottalico. 2022. "Embryo Culture, In Vitro Propagation, and Molecular Identification for Advanced Olive Breeding Programs" Horticulturae 8, no. 1: 36. https://doi.org/10.3390/horticulturae8010036
APA StyleMontilon, V., Susca, L., Potere, O., Roseti, V., Campanale, A., Saponari, A., Montemurro, C., Fanelli, V., Venerito, P., & Bottalico, G. (2022). Embryo Culture, In Vitro Propagation, and Molecular Identification for Advanced Olive Breeding Programs. Horticulturae, 8(1), 36. https://doi.org/10.3390/horticulturae8010036