
The Response of Antioxidant System of Drought-Stressed Green Pea (Pisum sativum L.) Affected by Watering and Foliar Spray with Silica Nanoparticles

 ,
,  ,
,
Abstract
:
1. Introduction
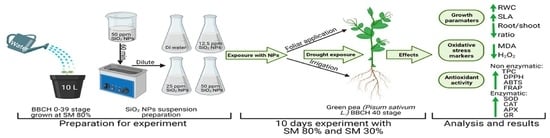
2. Materials and Methods
2.1. Study Site and Treatments
2.2. Relative Water Content, Specific Leaf Area, and Root/Shoot Ratio
2.3. Antioxidant Properties and Total Phenolic Compounds
2.4. Malondialdehyde and Hydrogen Peroxide
2.5. Antioxidant Enzymes Activities
2.6. Statistical Analysis
3. Results
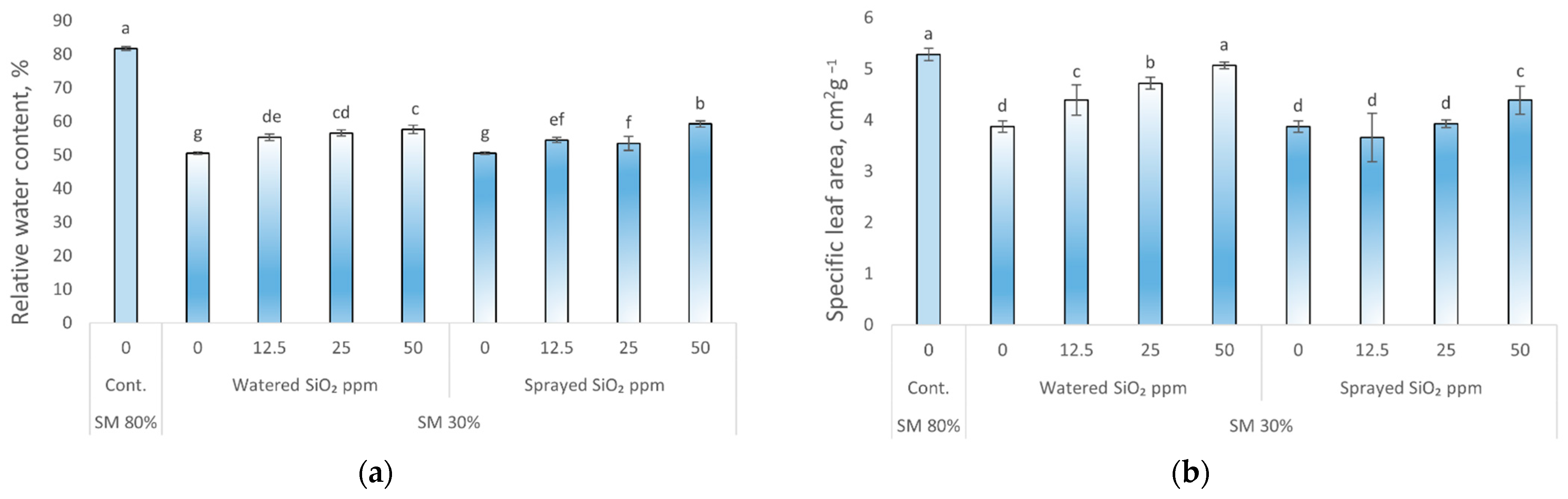
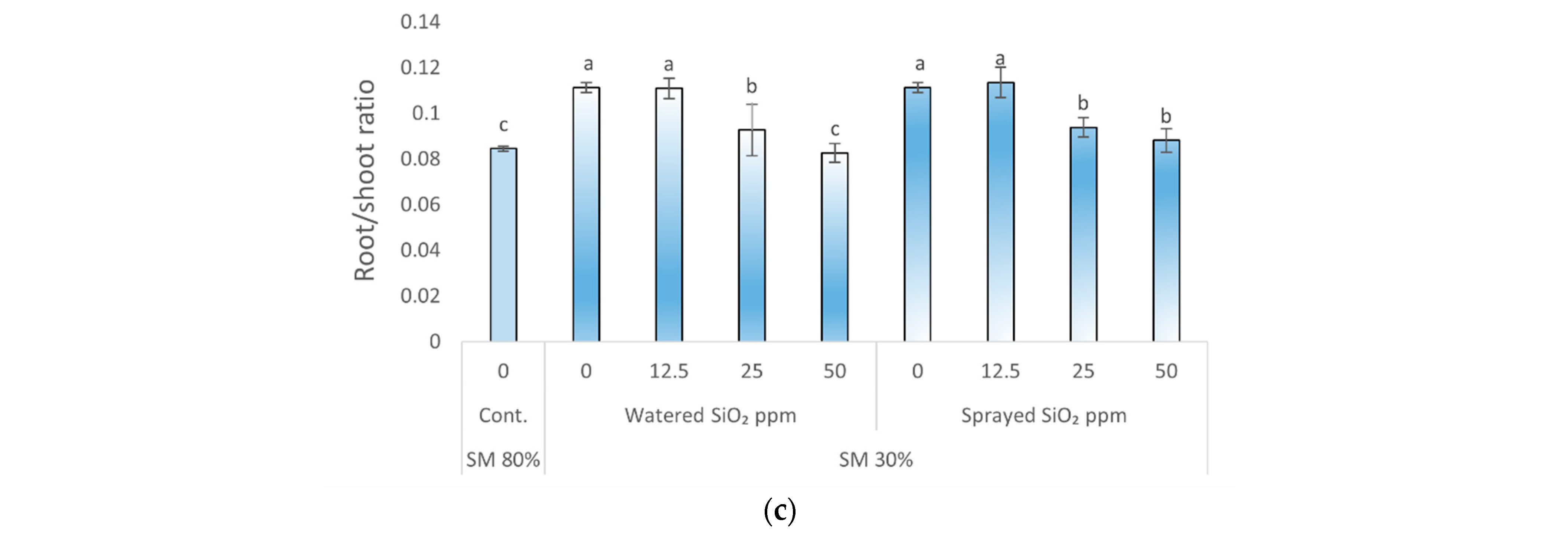
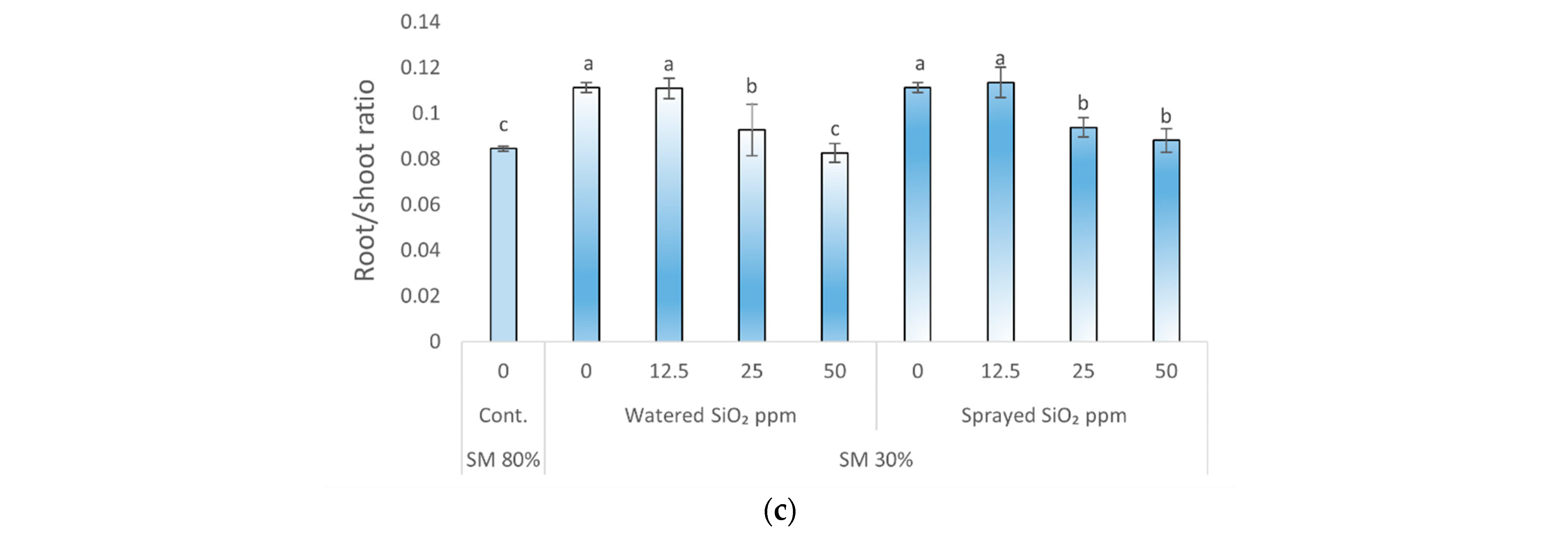
3.1. Effects of Drought Stress and SiO2 NPs on Plants Specific Leaf Area, Relative Water Content, and Root/Shoot Ratio
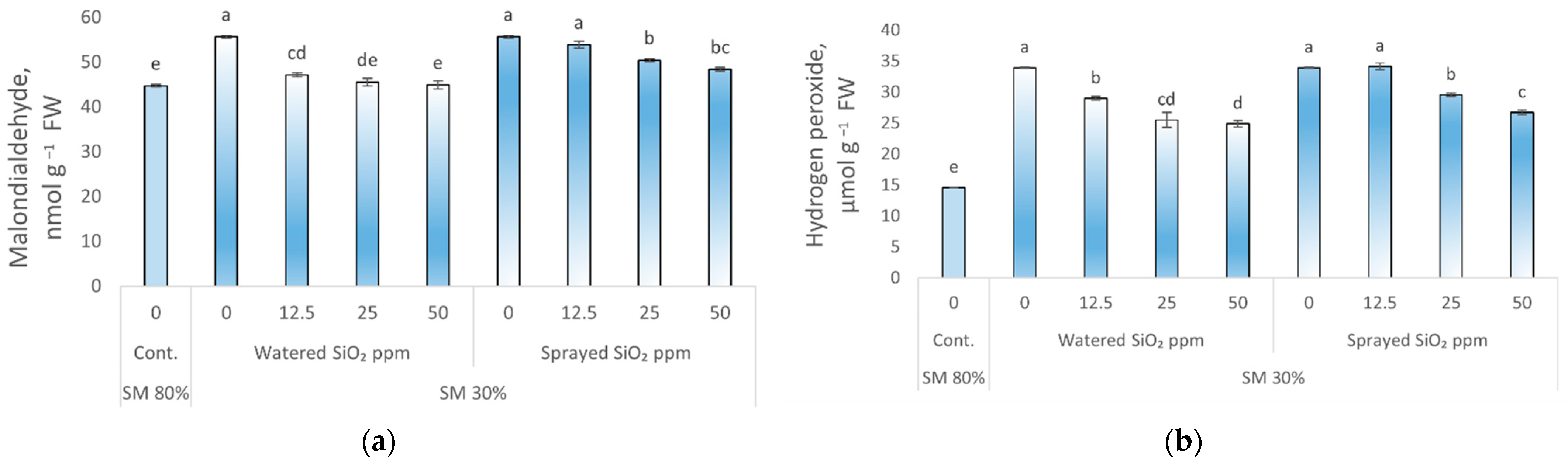
3.2. Influence of Drought Stress and SiO2 NPs on Lipid Peroxidation and Hydrogen Peroxide
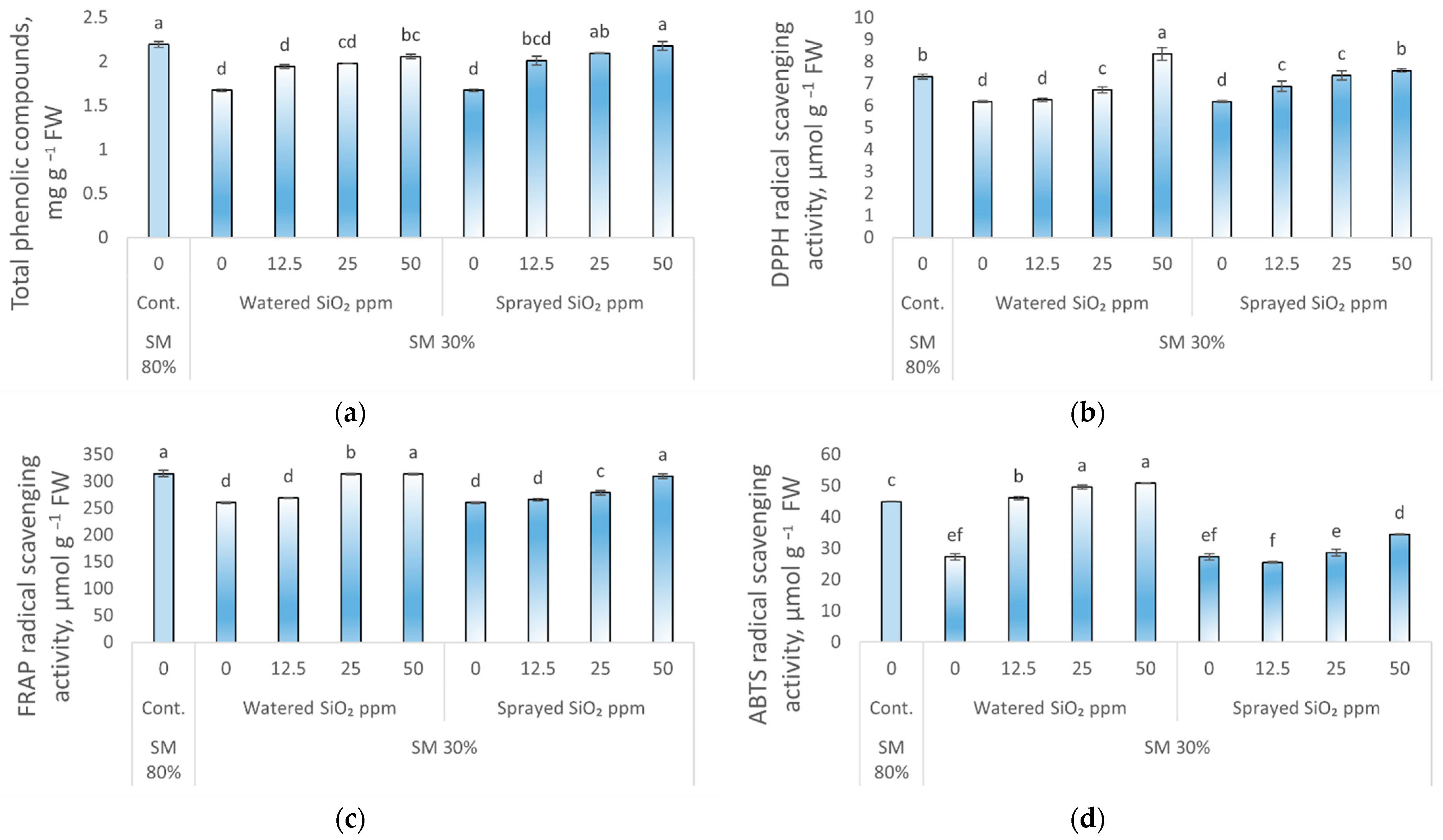
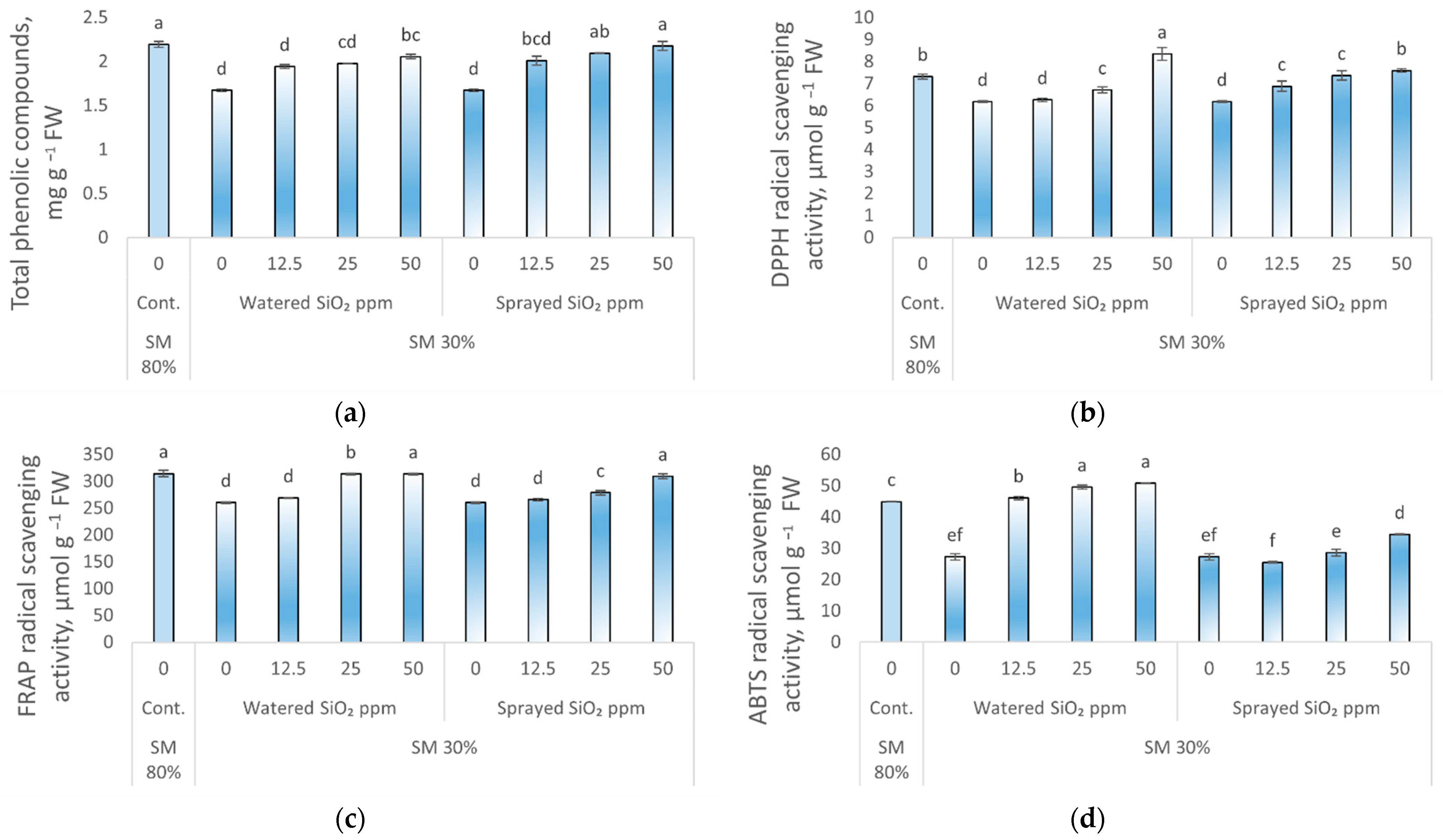
3.3. Effect of Drought Stress and SiO2 NPs on Antioxidant Activity
3.4. Effect of Drought Stress and SiO2 NPs on Antioxidant Enzymes Activities
4. Discussion
4.1. Effects of Different Applications and Concentration of NPs
4.2. Influence of Drought Stress and SiO2 NPs on Oxidative Stress Markers and Antioxidant Enzymes Activities in Peas
4.3. Effects of Drought Stress and SiO2 NPs on Peas Specific Leaf Area, Relative Water Content, and Root/Shoot Ratio
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spinoni, J.; Barbosa, P.; Bucchignani, E.; Cassano, J.; Cavazos, T.; Christensen, H.J.; Christensen, B.O.; Coppola, E.; Evans, J.; Geyer, B.; et al. Future global meteorological drought hot spots: A study based on CORDEX data. J. Clim. 2020, 33, 3635–3661. [Google Scholar] [CrossRef]
- Chhipa, H.; Joshi, P. Nanofertilisers, Nanopesticides and Nanosensors in Agriculture. Sustain. Agric. Rev. 2016, 20, 247–282. [Google Scholar]
- Zulfigar, F.; Navarro, M.; Ashraf, M.; Akram, A.N.; Munne-Bosch, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef]
- Gomez, A.; Narayan, M.; Zhao, L.; Jia, X.; Bernal, R.; Lopez-Moreno, L.M.; Peralta-Videa, J. Effects of nano-enabled agricultural strategies on food quality: Current knowledge and future research needs. J. Hazard. Mater. 2020, 401, 12391–12398. [Google Scholar] [CrossRef]
- Rui, Y.; Gui, X.; Li, X.; Liu, S.; Han, Y. Uptake, transport, distribution and bio-effects of SiO2 nanoparticles in Bt-transgenic cotton. J. Nanobiotechnol. 2014, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tarafdar, J.C.; Xiong, Y.; Wang, W.N.; Quinl, D.; Biswas, P. Standardization of size, shape and concentration of nanoparticle for plant application. Appl. Biol. Res. 2012, 14, 138–144. [Google Scholar]
- Mittal, D.; Kaur, G.; Singh, P.; Yadav, K.; Ali, S.A. Nanoparticle-based sustainable agriculture and food science: Recent advances and future outlook. Front. Nanotechnol. 2020, 2, 10. [Google Scholar] [CrossRef]
- Nadmint, P.P.; Dong, Y.D.; Sayer, C.; Hay, P.; Rookes, J.E.; Boyd, B.J.; Cahill, D.M. Nanostructured liquid crystalline particles as an alternative delivery vehicle for plant agrochemicals. ACS Appl. Mater. Interfaces 2013, 5, 1818–1826. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.; Pagay, V.; Stroock, D.A. Impact of electroviscosity on the hydraulic conductance of the bordered pit membrane: A theoretical investigation. Plant Physiol. 2013, 163, 999–1011. [Google Scholar] [CrossRef] [Green Version]
- Meychik, R.N.; Nikolaeva, I.J.; Yermakov, P.I. Ion exchange properties of the root cell walls isolated from the halophyte plants (Suaeda altissima L.) grown under conditions of different salinity. Plant Soil. 2005, 277, 163–174. [Google Scholar] [CrossRef]
- Sun, D.; Hussain, H.I.; Yi, Z.; Siegele, R.; Cresswell, T.; Kong, L.; Cahill, D.M. Uptake and cellular distribution, in four plant species, of fluorescently labeled mesoporous silica nanoparticles. Plant Cell Rep. 2014, 33, 1389–1402. [Google Scholar] [CrossRef]
- Luyckx, M.; Hausman, J.F.; Guerriero, G.; Lutts, S. Silicon and plants: Current knowledge and technological perspectives. Front. Plant Sci. 2017, 8, 411. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon nanoparticles more efficiently alleviate arsenate toxicity than silicon in maize cultivar and hybrid differing in arsenate tolerance. Front. Environ. Sci. 2016, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 17672. [Google Scholar] [CrossRef]
- Shabbaj, I.I.; Madany, M.M.Y.; Tammar, A.; Balkhyour, M.A.; AbdElgawad, H. Silicon Dioxide Nanoparticles Orchestrate Carbon and Nitrogen Metabolism in Pea Seedlings to Cope with Broomrape Infection. Environ. Sci. Nano 2021, 8, 1960–1977. [Google Scholar] [CrossRef]
- Tantawy, S.A.; Salama, M.Y.A.; El-Nemr, A.M.; Abdel-Mawgoud, R.A.M. Nano silicon application improves salinity tolerance of sweet pepper plants. Int. J. Chemtech Res. 2015, 8, 11–17. [Google Scholar]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Rehman, Z.M.; Malik, S.; Adrees, M.; Qayyum, F.M.; Alamri, A.S.; Alyemeni, N.M.; Ahmad, P. Effect of foliar applications of silicon and titanium dioxide nanoparticles on growth, oxidative stress, and cadmium accumulation by rice (Oryza sativa). Acta Physiol. Plant 2019, 41, 1–12. [Google Scholar] [CrossRef]
- Gowayed, H.M.; Al-Zahrani, S.H.; Metwali, M.E. Improving the salinity tolerance in potato (Solanum tuberosum) by exogenous application of silicon dioxide nanoparticles. Int. J. Agric. Biol. 2017, 19, 183–192. [Google Scholar]
- Tripathi, D.K.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon nanoparticles (SiNp) alleviate chromium (VI) phytotoxicity in Pisum sativum (L.) seedlings. Plant Physiol. Biochem. 2015, 96, 189–198. [Google Scholar] [CrossRef]
- Attia, A.E.; Elhawat, N. Combined foliar and soil application of silica nanoparticles enhances the growth, flowering period and flower characteristics of marigold (Tagetes erecta L.). Sci. Hortic. 2021, 282, 110015. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants: BBCH-Monograph Entwicklungsstadien Mono- und Dikotyler Pflanzen; Biologische Bundesanstalt für Land- und Forstwirtschaft in Berlin und Braunschweig, Ed.; Blackwell Wissenschafts-Verlag: Berlin, Germany; Wien, Austria, 1997; ISBN 9783826331527. [Google Scholar]
- Baris, D.H.; Weatherley, E.P. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 24, 519–570. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of Total Phenolic Content and Other Oxidation Substrates in Plant Tissues Using Folin–Ciocalteu Reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.R.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Sharma, P.O.; Bhat, K.T. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a aeasure of “Antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant system in acid rain treated bean plants: Protective role of exogenous polyammines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dhindsa, S.R.; Plumb-Dhindsa, P.; Thorpe, A.T. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Meth. Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Sofo, A.; Tuzio, C.A.; Dichio, B.; Xiloyannis, C. Influence of water deficit and rewatering on the components of the ascorbate–glutathione cycle in four interspecific Prunus hybrids. Plant Sci. 2005, 169, 403–412. [Google Scholar] [CrossRef]
- Kumar, S.; Soukup, M.; Elbaum, R. Silicification in grasses: Variation between different cell types. Front. Plant Sci. 2017, 8, 438. [Google Scholar] [CrossRef] [Green Version]
- Maurel, C.; Boursiac, Y.; Luu, T.D.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef]
- Rao, G.B.; Susmitha, P. Silicon uptake, transportation and accumulation in rice. J. Pharmacogn. Phytochem. 2017, 6, 290–293. [Google Scholar]
- Guntzer, F.; Keller, C.; Meunier, D.J. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Sangster, A.G.; Hodson, M.J.; Tubb, H.J. Silicon deposition in higher plants. In Studies in Plant Science; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; Volume 8, pp. 85–113. [Google Scholar] [CrossRef]
- Roduner, E. Size matters: Why nanomaterials are different. Chem. Soc. Rev. 2006, 35, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.H.; Al-Whaibi, M.H. Role of nano-SiO2 in germination of tomato (Lycopersicum esculentum seeds Mill.). Saudi J. Biol. Sci. 2014, 21, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Palocci, C.; Valletta, A.; Chronopoulou, L.; Donati, L.; Bramosanti, M.; Brasili, E.; Baldan, B.; Pasqua, G. Endocytic pathways involved in PLGA nanoparticle uptake by grapevine cells and role of cell wall and membrane in size selection. Plant Cell Rep. 2017, 36, 1917–1928. [Google Scholar] [CrossRef]
- Zeng, Y.; Himmel, M.E.; Ding, S.Y. Visualizing chemical functionality in plant cell walls. Biotechnol. Biofuels 2017, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Darlington, T.K.; Neigh, M.A.; Spencer, T.M.; Guyen, T.O.; Oldenburg, S.J. Nanoparticle characteristics affecting environmental fate and transport through soil. Environ. Toxicol. Chem. 2009, 28, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Schrick, B.; Hydusky, W.B.; Blough, L.J.; Mallouk, E.T. Delivery vehicles for zerovalent metal nanoparticles in soil and groundwater. Chem. Mater. 2004, 16, 2187–2193. [Google Scholar] [CrossRef]
- Kumar, A.; Dixit, K.C. Methods for Characterization of Nanoparticles. In Advances in Nanomedicine for the Delivery of Therapeutic Nucleic Acids; Nimesh, S., Chandra, R., Gupta, N., Eds.; Woodhead Publishing: Cambridge, UK, 2017; pp. 43–58. [Google Scholar] [CrossRef]
- Clogston, J.D.; Patri, A.K. Zeta potential measurement. Methods Mol. Biol. 2011, 697, 63–70. [Google Scholar] [CrossRef]
- Kinraide, T.B.; Wang, P. The surface charge density of plant cell membranes (σ): An attempt to resolve conflicting values for intrinsic σ. J. Exp. Bot. 2010, 61, 2507–2518. [Google Scholar] [CrossRef]
- El-Naggar, M.E.; Abdelsalam, N.R.; Fouda, M.M.; Mackled, M.I.; Al-Jaddadi, M.A.; Ali, H.M.; Siddiqui, M.H.; Kandil, E.E. Soil application of nano silica on maize yield and its insecticidal activity against some stored insects after the post-harvest. Nanomaterials 2020, 10, 739. [Google Scholar] [CrossRef]
- Zhu, J.Z.; Wang, H.; Yan, B.; Zheng, H.; Jiang, Y.; Mirand, R.O.; Rotello, M.V.; Xing, B.; Vachet, W.R. Effect of surface charge on the uptake and distribution of gold nanoparticles in four plant species. Environ. Sci. Technol. 2012, 46, 12391–12398. [Google Scholar] [CrossRef]
- Alsaeedi, A.; El-Ramady, H.; Alshaal, A.T.; Almohsen, M. Enhancing seed germination and seedlings development of common bean (Phaseolus vulgaris) by SiO2 nanoparticles. Egypt. J. Soil Sci. 2017, 54, 407–415. [Google Scholar]
- Alsaeedi, A.; El-Ramady, H.; Alshaal, T.; El-Garawany, M.; Elhawat, N.; Al-Otaibi, A. Silica nanoparticles boost growth and productivity of cucumber under water deficit and salinity stresses by balancing nutrients uptake. Plant Physiol. Biochem. 2019, 139, 1–10. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Miller, J.N.; Rice-Evans, A.C. Factors influencing the antioxidant activity determined by the ABTS•+ radical cation assay. Free Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef]
- Behboudi, F.; Tahmasebi Sarvestani, S.; Kassaee, M.Z.; Modares Sanavi, S.A.M.; Sorooshzadeh, A. Improving growth and yield of wheat under drought stress via application of SiO2 nanoparticles. J. Agric. Sci. Technol. 2018, 20, 1479–1492. [Google Scholar]
- Noctor, G.; Veljovic-Jovanovic, S.; Driscoll, S.; Novitskaya, L.; Foyer, H.C. Drought and oxidative load in the leaves of C3 plants: A predominant role for photorespiration? Ann. Bot. 2002, 89, 841–850. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, F.M.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Grela, R.E.; Samolinska, W.; Kiczorowska, B.; Klebaniuk, R.; Kiczorowski, P. Content of minerals and fatty acids and their correlation with phytochemical compounds and antioxidant activity of Leguminous seeds. Biol. Trace Elem. Res. 2017, 180, 338–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, B.; Lin, Q.; Hamid, Y.; Sanaullah, M.; Di, L.; Khan, B.M.; He, Z.; Yang, X. Foliage application of selenium and silicon nanoparticles alleviates Cd and Pb toxicity in rice (Oryza sativa L.). Sci. Total Environ. 2020, 712, 136497. [Google Scholar] [CrossRef]
- Rehman, U.S.; Bilal, M.R.; Rana, M.R.; Tahir, N.M.; Shah, N.M.K.; Ayalew, H.; Yan, G. Cell membrane stability and chlorophyll content variation in wheat (Triticum aestivum) genotypes under conditions of heat and drought. Crop Pasture Sci. 2016, 67, 712–718. [Google Scholar] [CrossRef]
- Mateos-Naranjo, E.; Andrades-Moreno, L.; Davy, A.J. Silicon alleviates deleterious effects of high salinity on the halophytic grass Spartina densiflora. Plant Physiol. Biochem. 2013, 63, 115–121. [Google Scholar] [CrossRef]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant. 2015, 37, 9. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Suspension of SiO2 NPs | |
|---|---|
| Zeta potential (ζ; mV) | −20.64 ± 0.333 |
| Polydispersity index (PI) | 0.34 |
| NPs size 1–100 nm in suspension (%) | 70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutulienė, R.; Ragelienė, L.; Samuolienė, G.; Brazaitytė, A.; Urbutis, M.; Miliauskienė, J. The Response of Antioxidant System of Drought-Stressed Green Pea (Pisum sativum L.) Affected by Watering and Foliar Spray with Silica Nanoparticles. Horticulturae 2022, 8, 35. https://doi.org/10.3390/horticulturae8010035
Sutulienė R, Ragelienė L, Samuolienė G, Brazaitytė A, Urbutis M, Miliauskienė J. The Response of Antioxidant System of Drought-Stressed Green Pea (Pisum sativum L.) Affected by Watering and Foliar Spray with Silica Nanoparticles. Horticulturae. 2022; 8(1):35. https://doi.org/10.3390/horticulturae8010035
Chicago/Turabian StyleSutulienė, Rūta, Lina Ragelienė, Giedrė Samuolienė, Aušra Brazaitytė, Martynas Urbutis, and Jurga Miliauskienė. 2022. "The Response of Antioxidant System of Drought-Stressed Green Pea (Pisum sativum L.) Affected by Watering and Foliar Spray with Silica Nanoparticles" Horticulturae 8, no. 1: 35. https://doi.org/10.3390/horticulturae8010035
APA StyleSutulienė, R., Ragelienė, L., Samuolienė, G., Brazaitytė, A., Urbutis, M., & Miliauskienė, J. (2022). The Response of Antioxidant System of Drought-Stressed Green Pea (Pisum sativum L.) Affected by Watering and Foliar Spray with Silica Nanoparticles. Horticulturae, 8(1), 35. https://doi.org/10.3390/horticulturae8010035