
A Reliable Regeneration Method in Genome-Editable Bell Pepper ‘Dempsey’

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Pepper Sowing
2.2. Shooting and Shoot Elongation
2.3. Rooting and Acclimation
2.4. Cytogenetic Analysis
2.5. Statistical Analysis
3. Results
3.1. Regeneration Procedures of Bell Pepper ‘Dempsey’
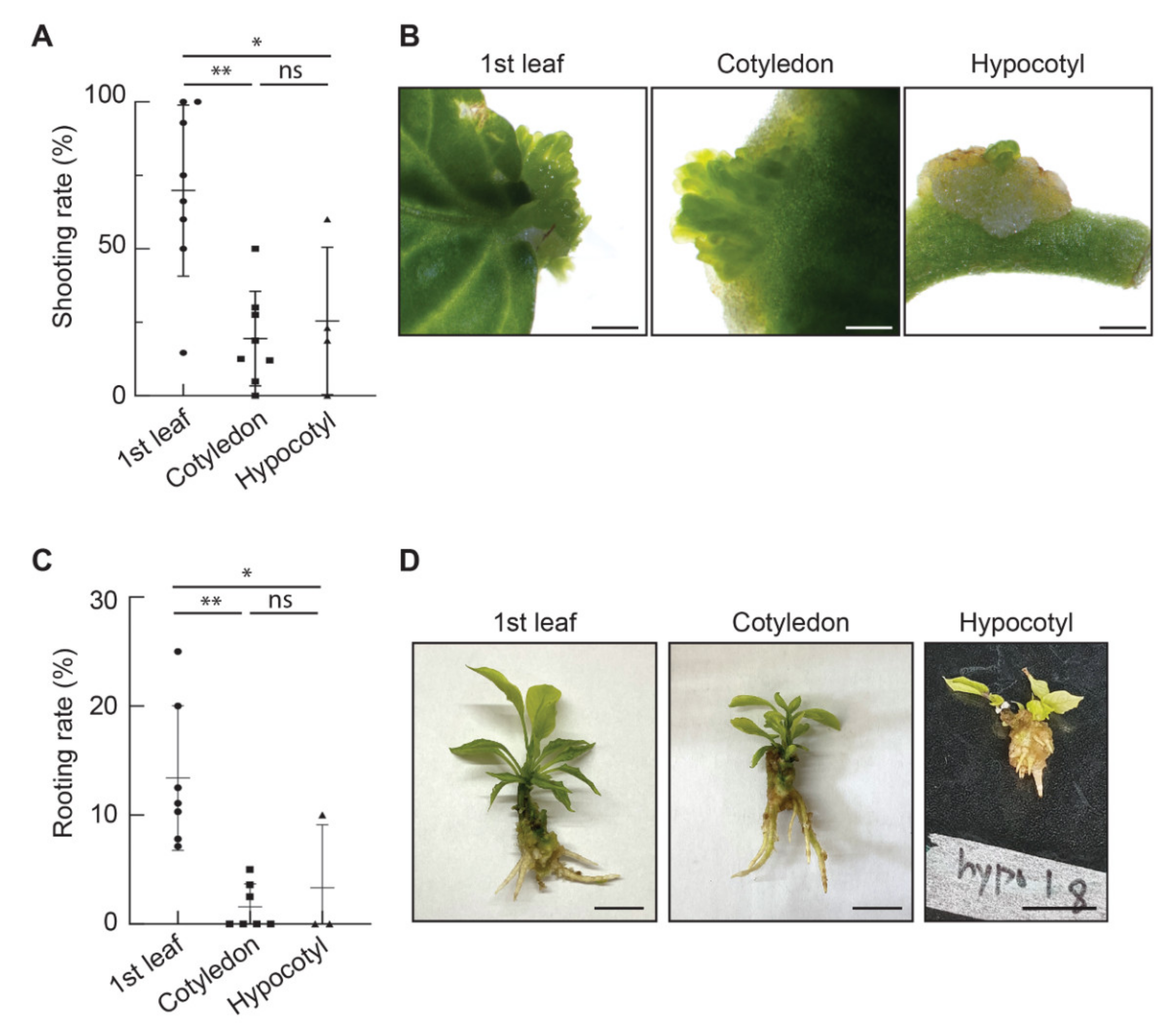
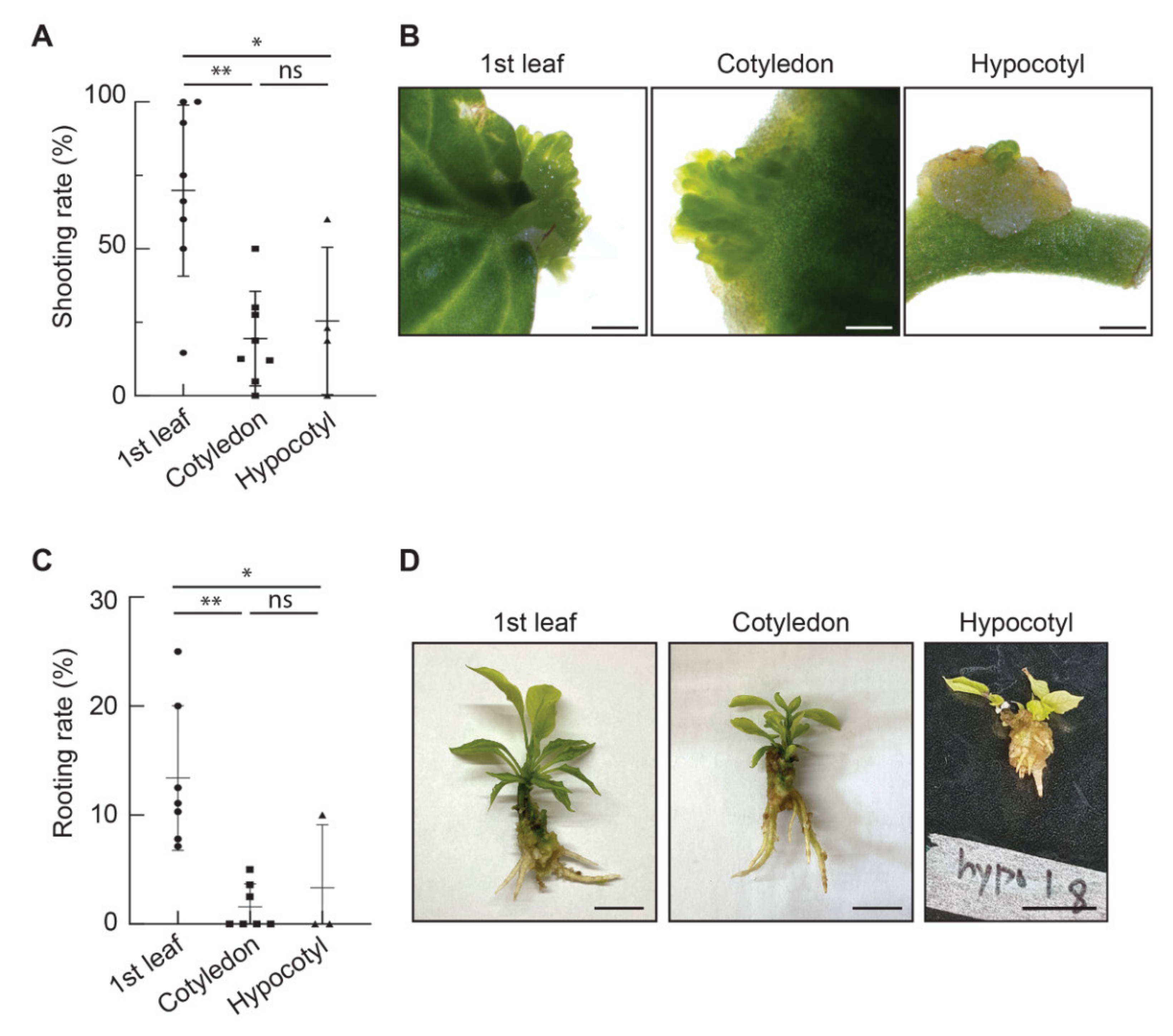
3.2. Comparison of Organogenesis Efficiencies among the Three Explant Types
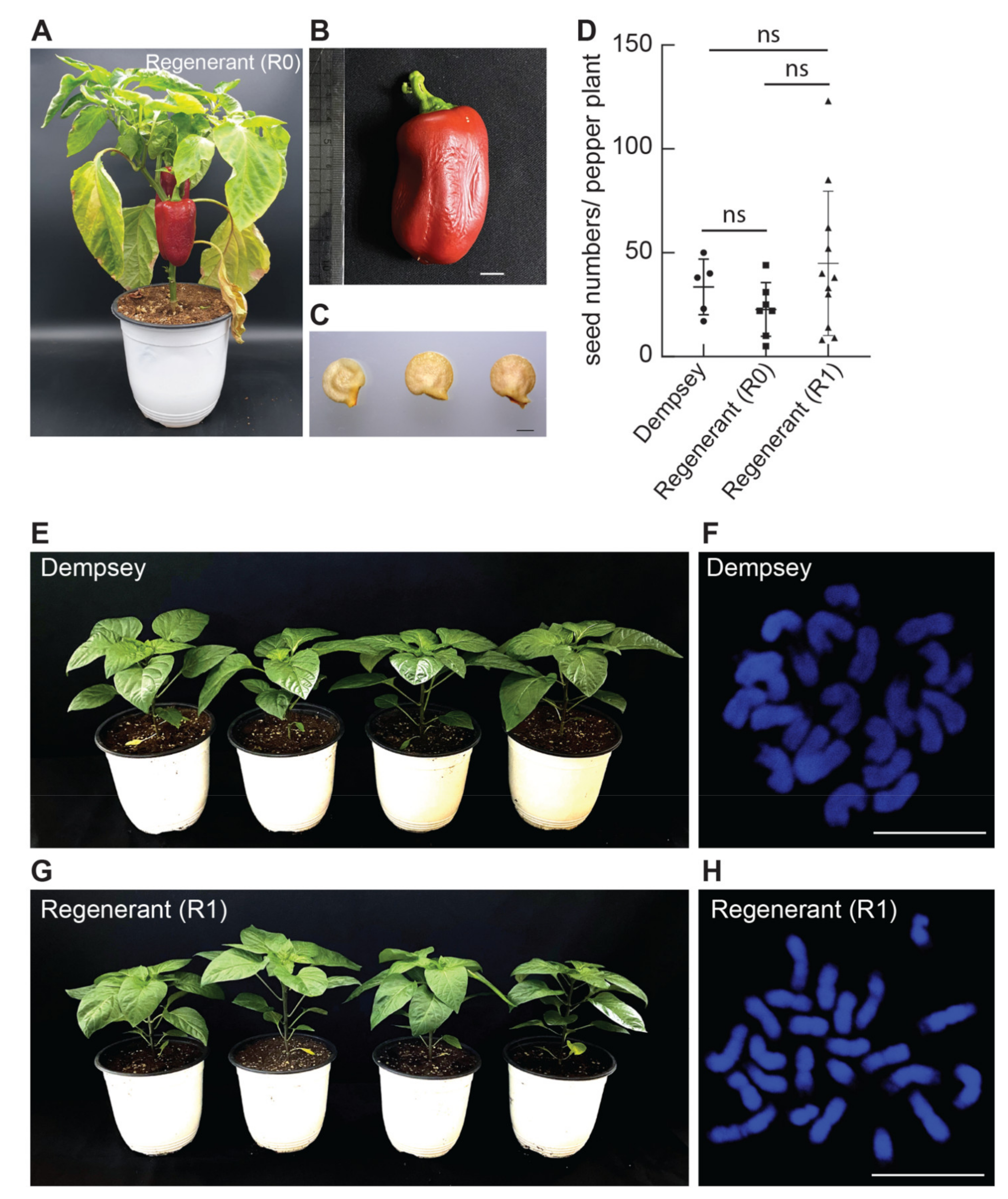
3.3. Phenotypic and Cytogenetic Analysis of ‘Dempsey’ Regenerants and Their Progenies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pickersgill, B. Genetic resources and breeding of Capsicum spp. Euphytica 1997, 96, 129–133. [Google Scholar] [CrossRef]
- Kang, J.H.; Yang, H.B.; Jeong, H.S.; Choe, P.; Kwon, J.K.; Kang, B.C. Single nucleotide polymorphism marker discovery from transcriptome sequencing for marker-assisted backcrossing in Capsicum. Korean J. Hortic. Sci. Technol. 2014, 32, 535–543. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, K.; Jang, S.; Lee, J.-H.; Lee, D.-G.; Kwon, J.-K.; Kang, B.-C. A MYB transcription factor is a candidate to control pungency in Capsicum annuum. Theor. Appl. Genet. 2019, 132, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Blum, E.; Mazourek, M.; O’Connell, M.; Curry, J.; Thorup, T.; Liu, K.; Jahn, M.; Paran, I. Molecular mapping of capsaicinoid biosynthesis genes and quantitative trait loci analysis for capsaicinoid content in Capsicum. Theor. Appl. Genet. 2003, 108, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.P.; McCarter, S.M.; Kuhn, C.W.; Deom, C.M. “Dempsey”, a virus- and bacterial spot-resistant bell pepper. HortScience 1997, 32, 333–334. [Google Scholar] [CrossRef] [Green Version]
- Hill, T.A.; Ashrafi, H.; Reyes-Chin-Wo, S.; Yao, J.; Stoffel, K.; Truco, M.-J.; Kozik, A.; Michelmore, R.W.; Van Deynze, A. Characterization of Capsicum annuum genetic diversity and population structure based on parallel polymorphism discovery with a 30K unigene Pepper GeneChip. PLoS ONE 2013, 8, e56200. [Google Scholar] [CrossRef] [Green Version]
- Han, K.; Jeong, H.-J.; Yang, H.-B.; Kang, S.-M.; Kwon, J.-K.; Kim, S.; Choi, D.; Kang, B.-C. An ultra-high-density bin map facilitates high-throughput QTL mapping of horticultural traits in pepper (Capsicum annuum). DNA Res. 2016, 23, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Han, K.; Lee, H.-Y.; Ro, N.-Y.; Hur, O.-S.; Lee, J.-H.; Kwon, J.-K.; Kang, B.-C. QTL mapping and GWAS reveal candidate genes controlling capsaicinoid content in Capsicum. Plant Biotechnol. J. 2018, 16, 1546–1558. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.-L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef]
- Woo, J.W.; Kim, J.; Kwon, S.I.; Corvalan, C.; Cho, S.W.; Kim, H.; Kim, S.-G.; Kim, S.-T.; Choe, S.; Kim, J.-S. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.T.; Ryu, J.; Kang, B.C.; Kim, J.S.; Kim, S.G. CRISPR/Cpf1-mediated DNA-free plant genome editing. Nat. Commun. 2017, 8, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanta, T.K.; Bashir, T.; Hashem, A.; Abd Allah, E.F.; Bae, H. Genome editing tools in plants. Genes 2017, 19, 399. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Zhang, Z.; Unver, T.; Zhang, B. CRISPR/Cas: A powerful tool for gene function study and crop improvement. J. Adv. Res. 2020, 21, 207–221. [Google Scholar] [CrossRef]
- Barton, K.A.; Binns, A.N.; Matzke, A.J.; Chilton, M.D. Regeneration of intact tobacco plants containing full length copies of genetically engineered T-DNA, and transmission of T-DNA to R1 progeny. Cell 1983, 32, 1033–1043. [Google Scholar] [CrossRef]
- Fillatti, J.J.; Kiser, J.; Rose, R.; Comai, L. Efficient transfer of a glyphosate tolerance gene into tomato using a binary Agrobacterium tumefaciens Vector. Bio/Technology 1987, 5, 726–730. [Google Scholar] [CrossRef]
- Han, E.H.; Goo, Y.M.; Lee, M.K.; Lee, S.W. An efficient transformation method for a potato (Solanum tuberosum L. var. Atlantic). J. Plant Biotechnol. 2015, 42, 77–82. [Google Scholar] [CrossRef]
- Muktadir, M.A.; Habib, M.A.; Khaleque Mian, M.A.; Yousuf Akhond, M.A. Regeneration efficiency based on genotype, culture condition and growth regulators of eggplant (Solanum melongena L.). Agric. Nat. Resour. 2016, 50, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.H.; Kim, H.S.; Kim, J.Y.; Jung, M.; Park, Y.S.; Lee, J.S.; Choi, S.H.; Her, N.H.; Lee, J.H.; Hyung, N.I.; et al. A new selection method for pepper transformation: Callus-mediated shoot formation. Plant Cell Rep. 2004, 23, 50–58. [Google Scholar] [CrossRef]
- Moon, K.K.; Soh, H.; Kim, K.M.; Young, S.K.; Im, K. Stable production of transgenic pepper plants mediated by Agrobacterium tumefaciens. HortScience 2007, 42, 1425–1430. [Google Scholar] [CrossRef]
- Maligeppagol, M.; Manjula, R.; Navale, P.M.; Prasad Babu, K.; Kumbar, B.M.; Laxman, R.H. Genetic transformation of chilli (Capsicum annuum L.) with Dreb1A transcription factor known to impart drought tolerance. Indian J. Biotechnol. 2016, 15, 17–24. [Google Scholar]
- Mahto, B.K.; Sharma, P.; Rajam, M.V.; Reddy, P.M.; Dhar-Ray, S. An efficient method for Agrobacterium-mediated genetic transformation of chilli pepper (Capsicum annuum L.). Indian J. Plant Physiol. 2018, 23, 573–581. [Google Scholar] [CrossRef]
- Bagga, S.; Lucero, Y.; Apodaca, K.; Rajapakse, W.; Lujan, P.; Ortega, J.L.; Sengupta-Gopalan, C. Chile (Capsicum annuum) plants transformed with the RB gene from Solanum bulbocastanum are resistant to Phytophthora capsici. PLoS ONE 2019, 14, e0223213. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Siddique, I.; Anis, M. Improved plant regeneration in Capsicum annuum L. from nodal segments. Biol. Plant. 2006, 50, 701–704. [Google Scholar] [CrossRef]
- Kumar, R.V.; Sharma, V.K.; Chattopadhyay, B.; Chakraborty, S. An improved plant regeneration and Agrobacterium—Mediated transformation of red pepper (Capsicum annuum L.). Physiol. Mol. Biol. Plants 2012, 18, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Mahto, B.K.; Singh, A.; Pareek, M.; Rajam, M.V.; Dhar-Ray, S.; Reddy, P.M. Host-induced silencing of the Colletotrichum gloeosporioides conidial morphology 1 gene (CgCOM1) confers resistance against Anthracnose disease in chilli and tomato. Plant Mol. Biol. 2020, 104, 381–395. [Google Scholar] [CrossRef]
- Prakash, A.H.; Sankara Rao, K.; Kumar, M.U. Plant regeneration from protoplasts of Capsicum annuum L. cv. California Wonder. J. Biosci. 1997, 22, 339–344. [Google Scholar] [CrossRef]
- Franck-Duchenne, M.; Wang, Y.; Ben Tahar, S.; Beachy, R.N. In vitro stem elongation of sweet pepper in media containing 24-epi-brassinolide. Plant Cell Tissue Organ Cult. 1998, 53, 79–84. [Google Scholar] [CrossRef]
- Dabauza, M.; Peña, L. High Efficiency Organogenesis in Sweet Pepper (Capsicum annuum L.) tissues from different seedling explants. Plant Growth Regul. 2001, 33, 221–229. [Google Scholar] [CrossRef]
- Gammoudi, N.; Pedro, T.S.; Ferchichi, A.; Gisbert, C. Improvement of regeneration in pepper: A recalcitrant species. In Vitro Cell. Dev. Biol. Plant 2018, 54, 145–153. [Google Scholar] [CrossRef]
- Kim, H.; Choi, J.; Won, K.-H. A stable DNA-free screening system for CRISPR/RNPs-mediated gene editing in hot and sweet cultivars of Capsicum annuum. BMC Plant Biol. 2020, 20, 449. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-I.; Kim, H.-B.; Jeon, H.-J.; Kim, H. Agrobacterium-mediated Capsicum annuum gene editing in two cultivars, hot pepper CM334 and bell pepper Dempsey. Int. J. Mol. Sci. 2021, 22, 3921. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Lamb, J.C.; Birchler, J.A. Chromosome painting using repetitive DNA sequences as probes for somatic chromosome identification in maize. Proc. Natl. Acad. Sci. USA 2004, 101, 13554–13559. [Google Scholar] [CrossRef] [Green Version]
- Waminal, N.E.; Pellerin, R.J.; Kim, N.S.; Jayakodi, M.; Park, J.Y.; Yang, T.J.; Kim, H.H. Rapid and efficient FISH using pre-labeled oligomer probes. Sci. Rep. 2018, 8, 8224. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.C.; Waminal, N.E.; Kim, H.H. In silico mining and FISH mapping of a chromosome-specific satellite DNA in Capsicum annuum L. Genes Genom. 2019, 41, 1001–1006. [Google Scholar] [CrossRef]
- Abd El-Twab, M.H.; Kondo, K. FISH physical mapping of 5S, 45S and Arabidopsis-type telomere sequence repeats in Chrysanthemum zawadskii showing intra-chromosomal variation and complexity in nature. Chromosome Bot. 2006, 1, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Lim, J. Leaf-induced callus formation in two cultivars: Hot pepper “CM334” and bell pepper “Dempsey”. Plant Signal. Behav. 2019, 14, 1604016. [Google Scholar] [CrossRef]
- Verma, S.; Dhiman, K.; Srivastava, D.K. Efficient in vitro regeneration from cotyledon explants in bell pepper (Capsicum annuum L. cv. California Wonder). Int. J. Adv. Biotechnol. Res. 2013, 4, 391–396. [Google Scholar]
- Akther, S.; Banu, T.; Khan, S.; Akter, S.; Habib, A.; Islam, M.; Goswami, B.; Sarkar, B. Micropropagation of two varieties of bell pepper (Capsicum annuum L.). Plant Tissue Cult. Biotechnol. 2020, 30, 267–275. [Google Scholar] [CrossRef]
- Kothari, S.L.; Joshi, A.; Kachhwaha, S.; Ochoa-Alejo, N. Chilli peppers—A review on tissue culture and transgenesis. Biotechnol. Adv. 2010, 28, 35–48. [Google Scholar] [CrossRef]
- Valadez-Bustos, M.G.; Aguado-Santacruz, G.A.; Carrillo-Castañeda, G.; Aguilar-Rincón, V.H.; Espitia-Rangel, E.; Montes-Hernández, S.; Robledo-Paz, A. In vitro propagation and agronomic performance of regenerated chili pepper (Capsicum spp.) plants from commercially important genotypes. In Vitro Cell. Dev. Biol. Plant 2009, 45, 650. [Google Scholar] [CrossRef]
- İzgü, T.; İlbi, H.; Mendi, Y.Y. Optimization of plant regeneration in different Pepper (Capsicum annuum L.). Turk. J. Agric. Food Sci. Technol. 2020, 8, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Liao, M.; Verbelen, J.P.; Vissenberg, K. Direct creation of marker-free tobacco plants from agroinfiltrated leaf discs. Plant Cell Rep. 2007, 26, 1961–1965. [Google Scholar] [CrossRef]
- Zottini, M.; Barizza, E.; Costa, A.; Formentin, E.; Ruberti, C.; Carimi, F.; Lo Schiavo, F. Agroinfiltration of grapevine leaves for fast transient assays of gene expression and for long-term production of stable transformed cells. Plant Cell Rep. 2008, 27, 845–853. [Google Scholar] [CrossRef]
- Faizal, A.; Geelen, D. Agroinfiltration of intact leaves as a method for the transient and stable transformation of saponin producing Maesa lanceolata. Plant Cell Rep. 2012, 31, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Hoshikawa, K.; Ezura, K.; Okazawa, R.; Fujita, S.; Takaoka, M.; Mason, H.S.; Ezura, H.; Miura, K. Improvement of the transient expression system for production of recombinant proteins in plants. Sci. Rep. 2018, 8, 4755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochoa-Alejo, N.; Ramirez-Malagon, R. In vitro chili pepper biotechnology. In Vitro Cell. Dev. Biol. Plant 2001, 37, 701–729, doiorg/101007/s11627. [Google Scholar] [CrossRef]
- Mezghani, N.; Jemmali, A.; Elloumi, N.; Gargouri-Bouzid, R.; Kintzios, S. Morpho-histological study on shoot bud regeneration in cotyledon cultures of pepper (Capsicum annuum). Biologia 2007, 62, 704–710. [Google Scholar] [CrossRef]
- Zimmerman, R.H. Rooting apple cultivars in vitro: Interactions among light, temperature, phloroglucinol and auxin. Plant Cell. Tissue Organ Cult. 1984, 3, 301–311. [Google Scholar] [CrossRef]
- Dumas, E.; Monteuuis, O. In vitro rooting of micropropagated shoots from juvenile and mature Pinus pinaster explants: Influence of activated charcoal. Plant Cell. Tissue Organ Cult. 1995, 40, 231–235. [Google Scholar] [CrossRef]
- Grozeva, S.; Todorova, V. In vitro regeneration in pepper (Capsicum annuum L.) and characterization of plant-regenerants. Electron. J. Biol. 2015, 11, 17–22. [Google Scholar]
- Barwale, U.B.; Widholm, J.M. Somaclonal variation in plants regenerated from cultures of soybean. Plant Cell Rep. 1987, 6, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Lewis-Smith, A.C.; Chamberlain, M.; Smith, S.M. Genetic and chromosomal variation in Petunia hybrida plants regenerated from protoplast and callus cultures. Biol. Plant. 1990, 32, 247–255. [Google Scholar] [CrossRef]
- Ezura, H.; Amagai, H.; Yoshioka, K.; Oosawa, K. Efficient production of tetraploid melon (Cucumis melo L.) by somatic embryogenesis. Jpn. J. Breed. 1992, 42, 137–144. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Media Name | Phytohormone | Solidifying Agent | Duration | |
|---|---|---|---|---|
| Shoot Induction Medium (SIM) | 8 mg/L BAP, 6 mg/L IAA | 0.8% agar A | 4 weeks | |
| Shoot Elongation Medium (SEM), 3 times subcultures | 2 mg/L Zeatin, 2 mg/L GA3 | 0.4% gelrite | 5 days | 6 weeks |
| 0.8% agar A | 9 days | |||
| Root Inducing Medium (RIM) | 0.1 mg/L NAA | 0.8% agar A | 3–4 weeks | |
| ‘Dempsey’ (WT) | Number of Fruit | Number of Seed | Total Seeds | Regenerant (R0) | Number of Fruit | Number of Seed | Total Seeds | Regenerant (R1) | Number of Fruit | Number of Seed | Total Seeds |
|---|---|---|---|---|---|---|---|---|---|---|---|
| #1 | 1 | 50 | 50 | #1 | 1 | 23 | 44 | #1 | 1 | 123 | 123 |
| #2 | 1 | 40 | 40 | 2 | 19 | #2 | 1 | 85 | 85 | ||
| #3 | 1 | 38 | 38 | 3 | 2 | #3 | 1 | 32 | 62 | ||
| #4 | 1 | 23 | 23 | #2 | 1 | 19 | 31 | 2 | 30 | ||
| #5 | 1 | 17 | 17 | 2 | 12 | #4 | 1 | 10 | 52 | ||
| #3 | 1 | 25 | 25 | 2 | 13 | ||||||
| #4 | 1 | 22 | 22 | 3 | 29 | ||||||
| #5 | 1 | 22 | 22 | #5 | 1 | 40 | 40 | ||||
| #6 | 1 | 10 | 10 | #6 | 1 | 38 | 38 | ||||
| #7 | 1 | 5 | 5 | #7 | 1 | 33 | 33 | ||||
| #8 | 1 | 30 | 30 | ||||||||
| #9 | 1 | 14 | 14 | ||||||||
| #10 | 1 | 9 | 9 | ||||||||
| #11 | 1 | 8 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Won, K.-H.; Park, S.-i.; Choi, J.; Kim, H.H.; Kang, B.-C.; Kim, H. A Reliable Regeneration Method in Genome-Editable Bell Pepper ‘Dempsey’. Horticulturae 2021, 7, 317. https://doi.org/10.3390/horticulturae7090317
Won K-H, Park S-i, Choi J, Kim HH, Kang B-C, Kim H. A Reliable Regeneration Method in Genome-Editable Bell Pepper ‘Dempsey’. Horticulturae. 2021; 7(9):317. https://doi.org/10.3390/horticulturae7090317
Chicago/Turabian StyleWon, Kang-Hee, Sung-il Park, Jisun Choi, Hyun Hee Kim, Byoung-Cheorl Kang, and Hyeran Kim. 2021. "A Reliable Regeneration Method in Genome-Editable Bell Pepper ‘Dempsey’" Horticulturae 7, no. 9: 317. https://doi.org/10.3390/horticulturae7090317
APA StyleWon, K.-H., Park, S.-i., Choi, J., Kim, H. H., Kang, B.-C., & Kim, H. (2021). A Reliable Regeneration Method in Genome-Editable Bell Pepper ‘Dempsey’. Horticulturae, 7(9), 317. https://doi.org/10.3390/horticulturae7090317