
Screening of Suitable Plant Regeneration Protocols for Several Capsicum spp. through Direct Organogenesis


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Growth Conditions of the Starting Material
2.3. In Vitro Culture of Explants
2.4. Response of Explants and Their Characterization
2.5. Elongation, Rooting and Acclimatization
2.6. Statistical Analysis
3. Results
3.1. Differences in Organogenic Response According to Explant and Medium
3.2. Differences in Shoot Formation among the Capsicum spp.
3.3. Elongation, Rooting and Acclimatization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT (Food and Agriculture of Organization of the United Nations Statistics Database). Available online: http://www.fao.org/faostat/es/#data/QC (accessed on 16 June 2021).
- Tripodi, P.; Kumar, S. The Capsicum Crop: An Introduction. In The Capsicum Genome; Ramchiary, N., Kole, C., Eds.; Compendium of Plant Genomes Springer Nature: Cham, Switzerland, 2019; pp. 1–8. [Google Scholar] [CrossRef]
- Monteiro do Rêgo, M.; Ramalho do Rêgo, E.; Barroso, P.A. Tissue Culture of Capsicum spp. In Production and Breeding of Chilli Peppers (Capsicum spp.); Springer International Publishing: Cham, Switzerland, 2016; pp. 97–127. [Google Scholar] [CrossRef]
- Park, S.; Kim, H.-B.; Jeon, H.-J.; Kim, H. Agrobacterium-mediated Capsicum annuum gene editing in two cultivars, hot pepper CM334 and bell pepper Dempsey. Int. J. Mol. Sci. 2021, 22, 3921. [Google Scholar] [CrossRef]
- Gunay, A.; Rao, P. In Vitro plant regeneration from hypocotyl and cotyledon explants of red pepper (Capsicum). Plant Sci. Lett. 1978, 11, 365–372. [Google Scholar] [CrossRef]
- Kothari, S.; Joshi, A.; Kachhwaha, S.; Ochoa-Alejo, N. Chilli peppers—A review on tissue culture and transgenesis. Biotechnol. Adv. 2010, 28, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Sanatombi, K.; Sharma, G. In Vitro plant regeneration in six cultivars of Capsicum spp. using different explants. Biol. Plant. 2008, 52, 141–145. [Google Scholar] [CrossRef]
- Kumar, A.; Rupavathi, O.; Tata, S. Adventitious shoot bud induction in chili pepper (Capsicum annuum L. CV. x-235). Int. J. Sci. Nat. 2012, 3, 192–196. [Google Scholar]
- Orlińska, M.; Nowaczyk, P. In Vitro plant regeneration of 4 Capsicum spp. Genotypes using different explant types. Turk. J. Biol. 2015, 39, 60–68. [Google Scholar] [CrossRef]
- Gammoudi, N.; Pedro, T.; Ferchichi, A.; Gisbert, C. Improvement of regeneration in pepper: A recalcitrant species. Plant. Vitr. Cell. Dev. Biol. Plant 2018, 54, 145–153. [Google Scholar] [CrossRef]
- Bora, G.; Gogoi, H.; Handique, P. Influence of silver nitrate and glutamine on in vitro organogenesis of lota bhot (Capsicum chinense Jacq.), an indigenous pungent pepper variety of Assam. J. Appl. Biol. Biotechnol. 2019, 7, 21–28. [Google Scholar] [CrossRef]
- Ashwani, S.; Ravishankar, G.A.; Giridhar, P. Silver nitrate and 2-(N-morpholine) ethane sulphonic acid in culture medium promotes rapid shoot regeneration from the proximal zone of the leaf of Capsicum frutescens Mill. Plant. Cell Tissue Organ. Cult. 2017, 129, 175–180. [Google Scholar] [CrossRef]
- Joshi, A.; Kothari, S. High copper levels in the medium improves shoot bud differentiation and elongation from the cultured cotyledons of Capsicum annuum L. Plant. Cell Tissue Organ. Cult. 2007, 88, 127–133. [Google Scholar] [CrossRef]
- Valadez-Bustos, M.; Aguado-Santacruz, G.; Carrillo-Castañeda, G.; Aguilar-Rincón, V.; Espitia-RangeL, E.; Montes-Hernández, S.; Robledo-Paz, A. In vitro propagation and agronomic performance of regenerated chili pepper (Capsicum spp.) plants from commercially important genotypes. Vitr. Cell. Dev. Biol. Plant 2009, 45, 650–658. [Google Scholar] [CrossRef]
- Dabauza, M.; Peña, L. High efficiency organogenesis in sweet pepper (Capsicum annuum L.) Tissues from different seedling explants. Plant. Growth Regul. 2001, 33, 221–229. [Google Scholar] [CrossRef]
- Hailu, T.; Abera, B.; Shekata, J. In Vitro Direct Organogenesis Protocol for Mass Propagation of an Elite Ethiopian Hot Pepper (Capsicum annuum L.) Cultivar: Mareko Fana. Am. J. Plant Sci. 2015, 6, 1435–1443. [Google Scholar] [CrossRef]
- Bora, G.; Gogoi, H.K.; Jyoti Handique, P. Micropropagation of Capsicum chinense Jacq. cv. Lota Bhot via indirect organogenesis molecular mechanisms of smut formation in economically important crops. Int. J. Agric. Sci. 2014, 6, 384–387. [Google Scholar] [CrossRef]
- Golegaonkar, P.; Kantharajah, A. High-frequency adventitious shoot bud induction and shoot elongation of chile pepper (Capsicum annuum L.). Vitr. Cell. Dev. Biol. Plant 2006, 42, 341–344. [Google Scholar] [CrossRef]
- Manzur, J.P.; Fita, A.; Prohens, J.; Rodríguez-Burruezo, A. Successful hybridization and introgression breeding in a diverse set of common peppers (Capsicum annuum) using different cultivated ají (C. baccatum) accessions as donor parents. PLoS ONE 2015, 10, e0144142. [Google Scholar] [CrossRef]
- García-Fortea, E.; Lluch-Ruiz, A.; Pineda-Chaza, B.; García-Pérez, A.; Bracho-Gil, J.; Plazas, M.; Gramazio, P.; Vilanova, S.; Moreno, V.; Prohens, J. A highly efficient organogenesis protocol based on zeatin riboside for in vitro regeneration of eggplant. BMC Plant. Biol. 2020, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Chee, M.; Lycett, G.; Chin, C. Development of a direct transformation method by GFP screening and in vitro whole plant regeneration of Capsicum frutescens L. Electron. J. Biotechnol. 2018, 34, 51–58. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ihaka, R.; Gentleman, R. A Language for Data Analysis and Graphics. J. Comput. Graph. Stat. 2012, 5, 299–314. [Google Scholar] [CrossRef]
- Zemene, A.; Worku, A. Protocol optimization for micro-propagation of Green pepper (Capsicum annuum L.) cultivated in Ethiopia. J. Med. Plants Stud. 2018, 6, 229–234. [Google Scholar]
- Abdalmajid, N.; Mohd, R.; Mihdzar, A.; Halimi, M. In vitro performances of hypocotyl and cotyledon explants of tomato cultivars under sodium chloride stress. Afr. J. Biotechnol. 2011, 10, 8757–8764. [Google Scholar] [CrossRef]
- Souza, F.; Garcia-Sogo, B.; Souza, A.; San-Juán, A.; Moreno, V. Morphogenetic response of cotyledon and leaf explants of melon (Cucumis melo L.) cv. Amarillo Oro. Braz. Arch. Biol. Technol. 2006, 49, 21–27. [Google Scholar] [CrossRef]
- Christopher, T.; Rajam, M. Effect of genotype, explant and medium on in vitro regeneration of red pepper. Plant. Cell Tissue Organ. Cult. 1996, 46, 245–250. [Google Scholar] [CrossRef]
- Trujillo-Moya, C.; Gisbert, C. The influence of ethylene and ethylene modulators on shoot organogenesis in tomato. Plant. Cell Tissue Organ. Cult. 2012, 111, 41–48. [Google Scholar] [CrossRef]
- Hussain, S.; Ali, S.; Ahmad Jan, S.; Ud-Din, J.; Ghulam, M.A. Assessment of silver nitrate on callus induction and in vitro shoot regeneration in tomato (Solanum lycopersicum mill.). Pak. J. Bot. 2014, 46, 2163–2174. [Google Scholar]
- Alva Ticona, S.; Oropeza, M. Effect of culture medium consistence and silver nitrate on micropropagation of two potato (Solanum tuberosum) cultivars. Rev. Colomb. Biotecnol. 2013, 15, 55–62. [Google Scholar] [CrossRef]
- Kaur, A.; Reddy, M.; Kumar, A. Efficient, one step and cultivar independent shoot organogenesis of potato. Physiol. Mol. Biol. Plants 2017, 23, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Papani, S.; Naidu, C.V. Antagonistic effect of silver nitrate and cobalt chloride against ethylene action to enhance in vitro regeneration potency of Solanum viarum Dunual. Int. J. Innov. Res. Adv. Stud. 2015, 2, 40–44. [Google Scholar]
- Geetha, G.; Harathi, K.; Naidu, C. Role of silver nitrate on in vitro flowering and shoot regeneration of Solanum nigrum (L.)—An Important Multipurpose Medicinal Plant. Am. J. Plant. Sci. 2016, 7, 1021–1032. [Google Scholar] [CrossRef][Green Version]
- Núñez-Pastrana, R.; Arcos-Ortega, G.F.; Souza-Perera, R.A.; Sánchez-Borges, C.A.; Nakazawa-Ueji, Y.E.; García-Villalobos, F.J.; Guzmán-Antonio, A.A.; Zúñiga-Aguilar, J.J. Ethylene, but not salicylic acid or methyl jasmonate, induces a resistance response against Phytophthora capsici in Habanero pepper. Eur. J. Plant. Pathol. 2011, 131, 669. [Google Scholar] [CrossRef]
- Grozeva, S. Effect of copper levels in the culture medium on shoot regeneration in pepper. Banat. J. Biotechnol. 2015, 6, 86. [Google Scholar] [CrossRef]
- Ciardi, J.; Klee, H. Regulation of ethylene-mediated responses at the level of the receptor. Ann. Bot. 2001, 88, 813–822. [Google Scholar] [CrossRef]
- İzgü, T.; İlbi, H.; Mendi, Y.Y. Optimization of plant regeneration in different pepper (Capsicum annuum L.) lines. Turk. J. Agric. Food Sci. Tech. 2020, 8, 471. [Google Scholar] [CrossRef]
- Robledo, P.; Carrillo, G. Regeneración in vitro de plantas de chile (Capsicum annuum L.) mediante cultivo de cotiledones e hipocótilos. Rev. Fitotec. Mex. 2004, 27, 121–126. [Google Scholar]






{kind=link}
{kind=link}
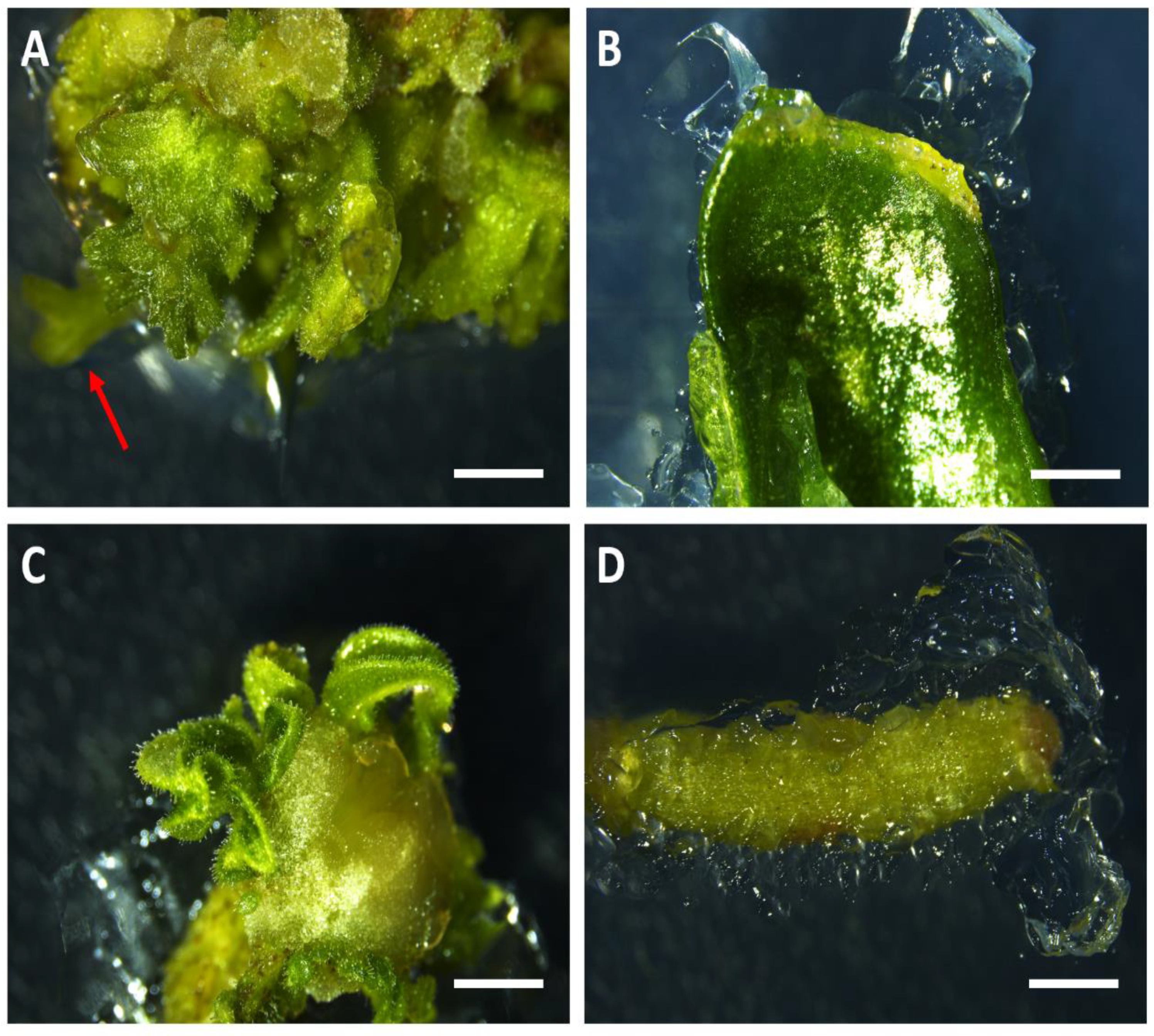{kind=link}
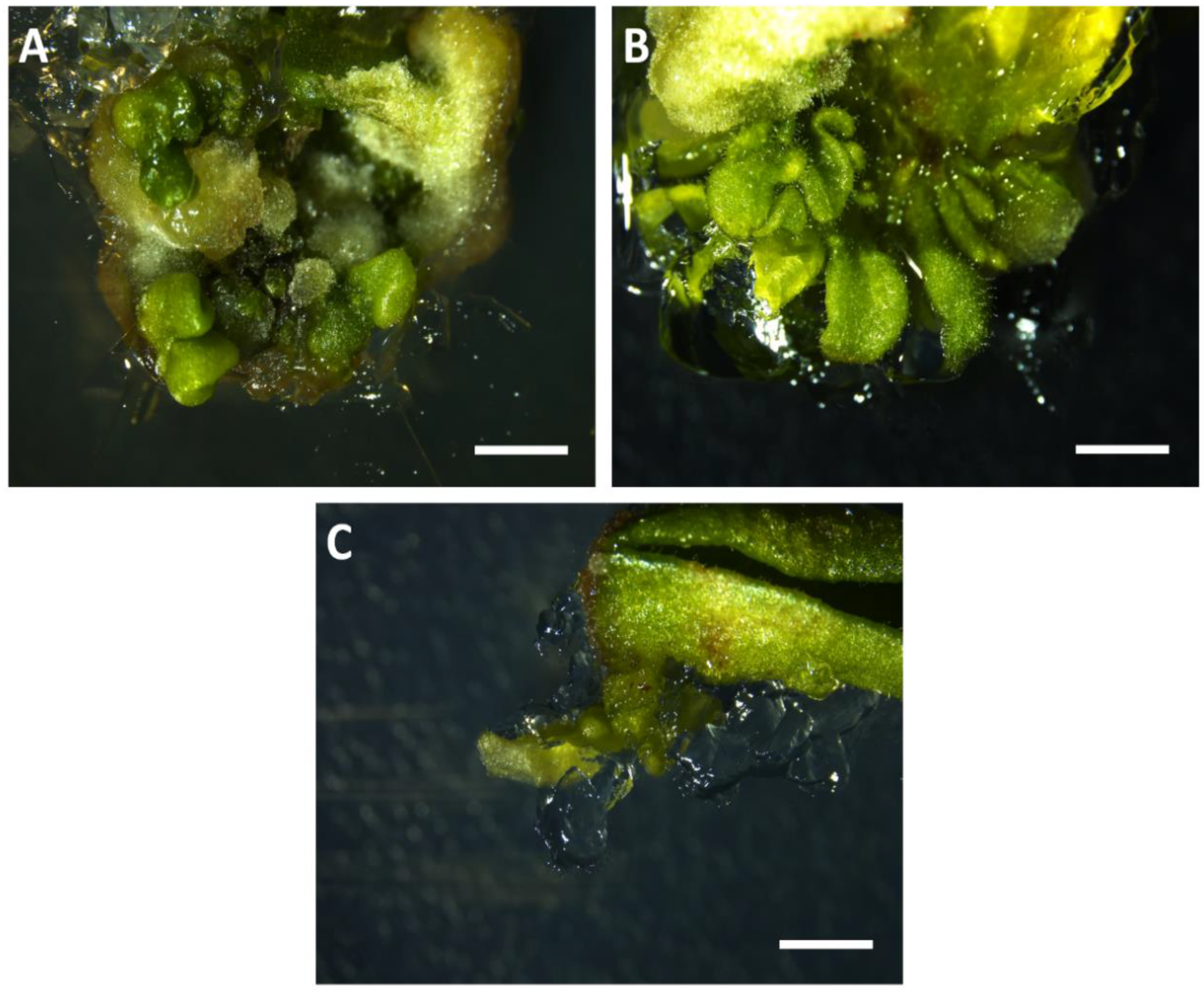{kind=link}
{kind=link}
| Medium | MS (g/L) | Sucrose (g/L) | Gelrite (g/L) | BAP (mg/L) | IAA (mg/L) | NAA (mg/L) | ZR (mg/L) | PAA (mg/L) | CuSO4 (mg/L) | AgNO3 (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 4.4 | 30 | 7 | - | - | - | - | - | - | - |
| Pep1 a | 4.4 | 30 | 7 | 5 | 0.5 | - | - | - | 0.47 | - |
| Pep2 | 4.4 | 30 | 7 | 10 | - | 0.1 | - | - | - | - |
| Pep3 b | 4.4 | 30 | 7 | 5 | 1 | - | - | - | - | - |
| Pep4 | 4.4 | 30 | 7 | 5 | - | - | - | 3 | 0.47 | - |
| Pep5 | 2.2 | 15 | 7 | - | - | - | 2 | - | - | - |
| Medium | MS (g/L) | Sucrose (g/L) | Gelrite (g/L) | Citric Acid (mg/L) | GA3 (mg/L) | IAA (mg/L) | IBA (mg/L) |
|---|---|---|---|---|---|---|---|
| E0 | 2.6 | 15 | 7 | - | - | - | - |
| ELG | 4.4 | 30 | 7 | 1 | 1 | 1 | - |
| R1 | 2.2 | 15 | 7 | - | - | - | 0.5 |
| R2 | 2.2 | 15 | 7 | - | - | - | 1 |
| Factors | Cut Edges with Calli | Tissue State | Number of Shoots Per Explant | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean a | Mode | Median | Range | Mean a | Mode | Median | Range | Mean a | Mode | Median | Range | |
| Explant | ||||||||||||
| Cotyledon | 1.50 a | 2 | 2 | 0–2 | 1.94 a | 1 | 2 | 1–3 | 1.44 a | 0 | 0 | 0–9 |
| Hypocotyl | 1.56 a | 2 | 2 | 0–2 | 1.54 b | 1 | 1 | 1–3 | 0.28 b | 0 | 0 | 0–8 |
| Medium | ||||||||||||
| Control | 0.50 c | 0 | 0.5 | 0–1 | 2.13 ab | 3 | 3 | 1–3 | 0.00 c | 0 | 0 | 0–0 |
| Pep1 + Pep1.2 | 1.80 ab | 2 | 2 | 1–2 | 1.07 d | 1 | 1 | 1–2 | 2.53 a | 0 | 2 | 0–8 |
| Pep2 | 1.67 b | 2 | 2 | 1–2 | 1.77 bc | 2 | 2 | 1–3 | 0.60 b | 0 | 0 | 0–8 |
| Pep3 + Pep3.2 | 1.73 b | 2 | 2 | 1–2 | 1.47 c | 1 | 1 | 1–2 | 2.00 a | 0 | 0 | 0–9 |
| Pep4 | 1.93 a | 2 | 2 | 1–2 | 2.23 a | 3 | 2.5 | 1–3 | 0.03 c | 0 | 0 | 0–1 |
| Pep5 | 1.53 b | 2 | 2 | 0–2 | 1.80 abc | 1 | 1 | 1–3 | 0.00 c | 0 | 0 | 0–0 |
| Factors | Cut Edges with Calli | Tissue State | Number of Shoots Per Explant | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean a | Mode | Median | Range | Mean a | Mode | Median | Range | Mean a | Mode | Median | Range | |
| Explant | ||||||||||||
| Cotyledon | 0.98 a | 1 | 1 | 0–2 | 1.81 a | 1 | 2 | 1–3 | 4.17 a | 0 | 3 | 0–19 |
| Hypocotyl | 1.19 a | 2 | 1 | 0–2 | 1.32 b | 1 | 1 | 1–3 | 3.20 a | 0 | 3 | 0–9 |
| Medium | ||||||||||||
| Control | 0.33 c | 0 | 0 | 0–2 | 2.13 a | 3 | 2.5 | 1–3 | 0.30 e | 0 | 0 | 0–4 |
| Pep1 + Pep1.2 | 1.00 b | 0 | 1 | 0–2 | 1.10 d | 1 | 1 | 1–2 | 7.87 a | 6 | 8 | 0–19 |
| Pep2 | 0.80 b | 0 | 1 | 0–2 | 1.47 bc | 1 | 1 | 1–2 | 1.57 d | 0 | 0.5 | 0–7 |
| Pep3 + Pep3.2 | 1.00 b | 1 | 1 | 0–2 | 1.60 bc | 1 | 1 | 1–3 | 5.67 b | 8 | 5 | 0–18 |
| Pep4 | 1.70 a | 2 | 2 | 0–2 | 1.43 c | 1 | 1 | 1–3 | 2.93 c | 0 | 2.5 | 0–7 |
| Pep5 | 1.67 a | 2 | 2 | 0–2 | 1.67 b | 2 | 2 | 1–2 | 3.77 bc | 3 | 3 | 0–9 |
| Factors | Cut Edges with Calli | Tissue State | Number of Shoots Per Explant | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean a | Mode | Median | Range | Mean a | Mode | Median | Range | Mean a | Mode | Median | Range | |
| Explant | ||||||||||||
| Cotyledon | 0.19 a | 0 | 0 | 0–2 | 2.12 a | 3 | 2 | 1–3 | 0.08 a | 0 | 0 | 0–4 |
| Hypocotyl | 0.31 a | 0 | 0 | 0–2 | 1.84 b | 1 | 2 | 1–3 | 0.00 a | 0 | 0 | 0–0 |
| Medium | ||||||||||||
| Control | 0.20 b | 0 | 0 | 0–1 | 1.96 abc | 1 | 1 | 1–3 | 0.00 a | 0 | 0 | 0–0 |
| Pep1 + Pep1.2 | 0.16 b | 0 | 0 | 0–1 | 1.64 c | 1 | 1 | 1–3 | 0.00 a | 0 | 0 | 0–0 |
| Pep2 | 0.08 bc | 0 | 0 | 0–1 | 2.08 b | 2 | 2 | 1–3 | 0.00 a | 0 | 0 | 0–0 |
| Pep3 + Pep3.2 | 0.00 c | 0 | 0 | 0–0 | 1.80 bc | 1 | 2 | 1–3 | 0.04 a | 0 | 0 | 0–1 |
| Pep4 | 0.00 c | 0 | 0 | 0–0 | 1.80 bc | 1 | 1 | 1–3 | 0.16 a | 0 | 0 | 0–4 |
| Pep5 | 1.12 a | 1 | 1 | 0–2 | 2.44 a | 2 | 2 | 2–3 | 0.00 a | 0 | 0 | 0–0 |
| Medium | Cotyledon | Hypocotyl | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean a | Mode | Median | Range | Mean a | Mode | Median | Range | |
| C. annuum | 1.45 AB | 0.28 A | ||||||
| Control | 0.00 c | 0 | 0 | 0–0 | 0.00 b | 0 | 0 | 0–0 |
| Pep1 + Pep1.2 | 4.40 a | 5 | 5 | 0–8 | 0.67 a | 0 | 0 | 0–4 |
| Pep2 | 1.20 b | 0 | 0 | 0–8 | 0.00 b | 0 | 0 | 0–0 |
| Pep3 + Pep3.2 | 3.07 ab | 0 | 2 | 0–9 | 0.93 a | 0 | 0 | 0–8 |
| Pep4 | 0.00 c | 0 | 0 | 0–0 | 0.07 ab | 0 | 0 | 0–1 |
| Pep5 | 0.00 c | 0 | 0 | 0–0 | 0.00 b | 0 | 0 | 0–0 |
| C. baccatum | 4.17 A | 3.20 B | ||||||
| Control | 0.00 c | 0 | 0 | 0–0 | 0.60 c | 0 | 0 | 0–4 |
| Pep1 + Pep1.2 | 8.87 a | 10 | 10 | 0–19 | 6.87 a | 6 | 6 | 4–9 |
| Pep2 | 3.07 b | 1 | 3 | 0–7 | 0.07 c | 0 | 0 | 0–1 |
| Pep3 + Pep3.2 | 7.33 a | 8 | 8 | 0–18 | 4.00 b | 5 | 4 | 0–8 |
| Pep4 | 2.60 b | 0 | 0 | 0–7 | 3.27 b | 2 | 3 | 1–7 |
| Pep5 | 3.13 b | 3 | 3 | 0–9 | 4.40 b | 6 | 6 | 0–7 |
| C. chinense | 0.08 B | 0.00 A | ||||||
| Control | 0.00 a | 0 | 0 | 0–0 | 0.00 a | 0 | 0 | 0–0 |
| Pep1 + Pep1.2 | 0.00 a | 0 | 0 | 0–0 | 0.00 a | 0 | 0 | 0–0 |
| Pep2 | 0.00 a | 0 | 0 | 0–0 | 0.00 a | 0 | 0 | 0–0 |
| Pep3 + Pep3.2 | 0.10 a | 0 | 0 | 0–1 | 0.00 a | 0 | 0 | 0–0 |
| Pep4 | 0.40 a | 0 | 0 | 0–4 | 0.00 a | 0 | 0 | 0–0 |
| Pep5 | 0.00 a | 0 | 0 | 0–0 | 0.00 a | 0 | 0 | 0–0 |
| Medium | C. annuum | C. baccatum | C. chinense | |||
|---|---|---|---|---|---|---|
| Cotyledon | Hypocotyl | Cotyledon | Hypocotyl | Cotyledon | Hypocotyl | |
| Control | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 26.7 ± 0.1 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Pep1 + Pep1.2 | 86.7 ± 0.1 | 33.3 ± 0.1 | 86.7 ± 0.1 | 100.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Pep2 | 46.7 ± 0.1 | 0.0 ± 0.0 | 93.3 ± 0.1 | 6.7 ± 0.1 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Pep3 + Pep3.2 | 66.7 ± 0.1 | 26.7 ± 0.1 | 86.7 ± 0.1 | 86.7 ± 0.1 | 10.0 ± 0.1 | 0.0 ± 0.0 |
| Pep4 | 0.0 ± 0.0 | 6.7 ±0.1 | 46.7 ± 0.1 | 100.0 ± 0.0 | 10.0 ± 0.1 | 0.0 ± 0.0 |
| Pep5 | 0.0 ± 0.0 | 0.0 ± 0.0 | 80.0 ± 0.1 | 86.7 ± 0.1 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| C. annuum | C. baccatum | C. chinense | ||||
|---|---|---|---|---|---|---|
| Cotyledon | Hypocotyl | Cotyledon | Hypocotyl | Cotyledon | Hypocotyl | |
| Number of explants to elongate | 20 | 5 | 43 | 51 | 5 | 0 |
| Explants with elongated shoot (%) | 80.0 ± 0.1 | 80.0 ± 0.2 | 46.5 ± 0.1 | 35.3 ± 0.1 | 40.0 ± 0.2 | - |
| Average number of elongated shoots per explant | 2.13 ± 0.26 | 1.25 ± 0.25 | 13.35 ± 1.69 | 3.94 ± 0.77 | 1.00 ± 0.00 | - |
| R1 | R2 | |||
|---|---|---|---|---|
| Cotyledon | Hypocotyl | Cotyledon | Hypocotyl | |
| C. annuum | ||||
| Number of shoots to root | 34 | 5 | - | - |
| Shoots with root (%) | 0.0 ± 0.0 | 0.0 ± 0.0 | - | - |
| C. baccatum | ||||
| Number of shoots to root | 267 | 71 | 22 | 28 |
| Shoots with root (%) | 12.0 ± 0.0 | 2.8 ± 0.0 | 54.5 ± 0.1 | 17.9 ± 0.1 |
| C. chinense | ||||
| Number of shoots to root | 2 | - | 1 | - |
| Shoots with root (%) | 50.0 ± 0.4 | - | 0.0 ± 0.0 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-López, M.; García-Pérez, A.; Gimeno-Páez, E.; Prohens, J.; Vilanova, S.; García-Fortea, E. Screening of Suitable Plant Regeneration Protocols for Several Capsicum spp. through Direct Organogenesis. Horticulturae 2021, 7, 261. https://doi.org/10.3390/horticulturae7090261
Martínez-López M, García-Pérez A, Gimeno-Páez E, Prohens J, Vilanova S, García-Fortea E. Screening of Suitable Plant Regeneration Protocols for Several Capsicum spp. through Direct Organogenesis. Horticulturae. 2021; 7(9):261. https://doi.org/10.3390/horticulturae7090261
Chicago/Turabian StyleMartínez-López, Marina, Ana García-Pérez, Esther Gimeno-Páez, Jaime Prohens, Santiago Vilanova, and Edgar García-Fortea. 2021. "Screening of Suitable Plant Regeneration Protocols for Several Capsicum spp. through Direct Organogenesis" Horticulturae 7, no. 9: 261. https://doi.org/10.3390/horticulturae7090261
APA StyleMartínez-López, M., García-Pérez, A., Gimeno-Páez, E., Prohens, J., Vilanova, S., & García-Fortea, E. (2021). Screening of Suitable Plant Regeneration Protocols for Several Capsicum spp. through Direct Organogenesis. Horticulturae, 7(9), 261. https://doi.org/10.3390/horticulturae7090261