Abstract
Adequate chill is of great importance for successful production of deciduous fruit trees. However, temperate fruit trees grown under tropical and subtropical regions may face insufficient winter chill, which has a crucial role in dormancy and productivity. The objective of this review is to discuss the challenges for dormancy and chilling requirements of temperate fruit trees, especially in warm winter regions, under climate change conditions. After defining climate change and dormancy, the effects of climate change on various parameters of temperate fruit trees are described. Then, dormancy breaking chemicals and organic compounds, as well as some aspects of the mechanism of dormancy breaking, are demonstrated. After this, the relationships between dormancy and chilling requirements are delineated and challenging aspects of chilling requirements in climate change conditions and in warm winter environments are demonstrated. Experts have sought to develop models for estimating chilling requirements and dormancy breaking in order to improve the adaption of temperate fruit trees under tropical and subtropical environments. Some of these models and their uses are described in the final section of this review. In conclusion, global warming has led to chill deficit during winter, which may become a limiting factor in the near future for the growth of temperate fruit trees in the tropics and subtropics. With the increasing rate of climate change, improvements in some managing tools (e.g., discovering new, more effective dormancy breaking organic compounds; breeding new, climate-smart cultivars in order to solve problems associated with dormancy and chilling requirements; and improving dormancy and chilling forecasting models) have the potential to solve the challenges of dormancy and chilling requirements for temperate fruit tree production in warm winter fruit tree growing regions.
1. Introduction
Climate change refers to any long-term changes in climate status (i.e., decades or longer) such as global atmospheric CO2 or mean temperature due to human activities or natural variability [1]. Climate change is defined in the United Nations Framework Convention on Climate Change (UNFCCC) as “a change of climate which is attributed directly or indirectly to human activity that alters the composition of the global atmosphere and which is in addition to natural climate variability observed over comparable time periods” [2]. Climate change is considered one of the most impactful phenomena on the production of temperate fruit crops [3]. All climatic elements affect plant growth, development and the productivity of different fruit crops. Temperate fruit crops undergo several physiological processes related to winter dormancy and chilling requirements; this is especially the case under tropical and subtropical conditions [4]. The major effect of climate change on temperate fruit crops may be attributed to increased atmospheric CO2 (413.95 ppm, December 2020) and temperature (0.97 °C, November 2020) [1,5].
Temperate fruit trees originate from different locations characterized by four well-defined seasons (spring, summer, fall and winter) with a very cold winter and warm spring–summer seasons for the dormancy and growth periods, respectively [6]. Temperate fruit trees account for about 48% of global fruit production, and therefore generate essential revenue and food [7]. Temperate fruits are adapted to the temperate zone climate in mid-latitudes [8]; however, about 50% of temperate fruit are cultivated in different climatic zones in China, Turkey, USA, Brazil, Italy, and Spain; these countries make up most of the world’s total production [9]. The rapid climatic changes that have occurred during the last few decades have led to undesirable physiological changes such as abnormal bud break, advanced or delayed flowering, interrupted fruit growth and ripening, and advanced leaf senescence [10]. Climatic changes may also alter the adaptability and sustainability of temperate fruit trees to their production regions. This is inciting scientists to develop strategies that may improve the adaptability of trees to such changes, particularly winter warming, which may be helpful in the interpretation of some ambiguous physiological and biochemical dormancy-related issues [11].
Dormancy, as a plant developmental phase, occurs in deciduous fruit trees to overcome the unfavorable environmental (e.g., weather) conditions that occur during winter [12,13,14]. Several trials have evaluated the role of various bioactive compounds incompensating for the chill deficit for dormancy breaking [15]. Most research during the past five decades has focused on phenology modeling, where it maybe possible to correlate plant phenological events with the surrounding temperature in order to identify the most effective temperature range to break dormancy [16,17]. Lang et al. [18] proposed different dormancy terminologies to distinguish among the various types of bud dormancy. Para-dormancy is growth suppression due to the effect of an inhibitor transported from one plant part to another one (e.g., apical dominance). Endo-dormancy describes growth inhibition related to a chill deficit; in this case, buds never open, even if all other climatic conditions are suitable. Eco-dormancy refers to growth inhibition due to insufficiently warm temperatures after winter. By the beginning of the 21st century, dormancy research had expanded to the molecular level to discover dormancy-related genes, such as Mads-box (DAM) in peach (Prunus persica) [19]. Gene expression research associates dormancy with hormonal and metabolic changes during the dormancy period [20]. Epigenetics (the modification of the gene expression without any change of gene sequences) is considered a promising mechanism for dormancy breaking [21]. Reactive oxygen species (ROS) were demonstrated to be key molecules, integrating metabolic processes and environmental cues in regulating physiological and phenological plant stress processes [22].
Temperatures on Earth are predicted to rise up to 6 °C by the end of this century, compared to those in the pre-industrial era [23]. This agroclimatic metric is unlikely to remain stable [24]. The advanced trend in the blooming time of several fruit trees indicates that the dormancy breaking process is significantly changing, most likely in response to climate change [25]. Plant breeding techniques to develop new cultivars suited for such a change in temperature would be a proper solution, but this may be a long process; therefore, other proper practices may be faster solutions at present. Dormancy breaking chemical products have been used in a wide range of applications to enhance earlier budburst in deciduous fruit trees [26]. These products include, for instance, mineral oils, cyanamides, thiourea, potassium nitrate, and growth regulators such as gibberellic acid and cytokinins [27]. Although current research studies on dormancy have made good progress to unveil and clarify some unclear issues [28], a comprehensive framework that integrates knowledge between dormancy and climate change for a clearer perspective is still missing.
Therefore, the aim of this review was to discuss the events relating to dormancy and chilling requirements of temperate fruit trees in warm winter regions in light of global climatic change.
2. Climate Change and Temperate Fruit Productivity
It is fundamental knowledge that the yield of temperate fruit trees, like that of other horticultural crops, is controlled by climate elements such as temperature, humidity, atmospheric CO2, precipitation, wind speed, sunlight, and UV index. Among these, temperature and atmospheric CO2 are the most highlighted climate elements for temperate tree crops. Extensive research in tree phenology has demonstrated that the increase in temperature and atmospheric CO2 levels ultimately influences the number of chill hours required for dormancy breaking and the yield and fruit quality of temperate fruit crops [10,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43] (examples are given in Table 1 and Table 2).

Table 1.
Some examples for the impacts of elevated temperature on deciduous fruit tree crops including countries and literature references (Ref.).
Table 2.
Some examples for the impacts of elevated CO2 on some fruit crops including countries and literature references (Ref.).
Several studies have evaluated the productivity of temperate fruit crops with the current status of global climate change [4,33,44,45,46,47,48,49,50,51,52,53]. These studies addressed the various challenges that impact temperate fruit production [4]. It was reported that changes in both agroclimatic and temperature indices were higher than the vulnerability threshold of apple production [33]. Research has focused on different phenological stages such as bud swelling, the onset and end of blooming of apple and pear trees [44], as well as the adaptability of fruit crops to climate change conditions [45]. In this regard, an evaluation of various adaptation strategies for grapevines was conducted [46,47,48,49]. The projected impacts of climate change on frost effect for some temperate-zone-grown crops such as almond, avocado, and orange were also studied through evaluating plant requirements of energy and water to mitigate frost damage [50]. Climate change impacts on yield and phenology of hazelnut trees in the maritime climate of Australia were also discussed [51]. The impact of climate on plant phenology as a decisive factor in defining the geographical range of fruit crops was evaluated for peach to create an ecological thermal niche map [52]. In addition, apple-based agroforestry systems for C-sequestration and biomass production were reported with the effect of climate change in the temperate region of Northern Himalaya, India [53]. Bioclimatic indices and plant phenology models assessing the suitability of temperate fruit species to adapt to climate changes were used to provide spatial and temporal information regarding the future changes in fruit quality [3]. In this regard, a model study revealed future climate changes and their effects on plant phenology of temperate deciduous trees in spring [17], including the role of climate changes in the re-distribution of cultivated temperate fruit crops, based on chilling accumulation [7]. It is also important to note that climate changes could lead to changes in the plant phenological processes (Figure 1) and susceptibility to different species of pests and diseases [10,34], which may negatively impact total crop yield and quality.

Figure 1.
Dormancy and key phenological stages of apple and sour cherry trees up to harvest, which can be seriously affected by climate change. Key growth stages for apple: dormancy (1), silver tip (2), pink tip (3), blossom (4), pollination (5), petal fall (6), fruit set (7), fruit growth (8), and mature fruit (9). Key growth stages for sour cherry: dormancy (a), bud swelling (b), pink tip (c), full bloom (d), pollination (e), petal fall (f), fruit set (g), and mature fruit with anthracnose disease (h).
3. Understanding the Role Dormancy
It is a natural phenomenon in deciduous fruit trees that dormancy occurs during the coldest period/season of the year, at which point growth is halted with modified cell physiology to avoid damage caused by cold temperature [7]. The decline in temperature and photoperiod during winter induces vital biological processes during dormancy that ultimately affect plant re-growth in spring [54]. This phenomenon was extensively studied from different perspectives [11,14,55] as a type of plant resistance to severe reduction in temperature [15]. During dormancy, trees show no visible growth and minimize all physiological processes during winter, and eventually resume growth in spring [11]. Physiological-wise, the trees can recognize the fluctuation in temperature among the seasons. Thus, trees start dormancy as the temperature decreases in late fall and early winter and then begin re-growth as the temperature increases in spring after being vulnerable to particular chilling hours during winter [12,56]. Each temperate fruit cultivar has certain agroclimatic phenological process requirements for the spring to fall seasons [7]. Each cultivar has its own chilling and warmth requirements, as reported for many species such as pome fruits and nuts, and such information is considered in theplanting of a specific cultivar in a given growing area [55]. Researchers investigated the physiological mechanisms of extended dormancy in spring alongside the rise in temperature [57,58]. Buds undergo various physiological changes during dormancy until flowering in spring [59]. However, unfavorable conditions (such as severe cold, drought, or salinity) may induce the physiological events of extended dormancy (eco-dormancy) [6]. Extended dormancy could also occur as a result of hormonal imbalance such as growth promoting hormones, gibberellins, and growth retardants (e.g., abscisic acid) even under suitable weather conditions. This event results in stopping plant growth and blooming (para-dormancy) [14,18]. Dormancy can also occur when insufficient chilling requirements in winter or warm temperature in spring causes an extended dormancy (endo-dormancy) [57,60]. This event is considered the most limiting blooming factor in deciduous fruit trees and may lead to reduced fruit set with increased percentage of malformed fruit affecting total fruit yield [14]. Therefore, the physiological, biochemical, and molecular aspects of flower bud dormancy for temperate fruit species are essential factors in successful fruit production, especially under climate change conditions [61].
3.1. Dormancy Breaking Chemical Compounds
Under temperate climatic conditions, deciduous fruit trees need certain chilling hours during winter for dormancy breaking by the beginning of spring. Chilling requirements differ from one species/cultivar to another, and inadequate chilling hours may lead to several physiological disorders such as uneven bud break, weak vegetative growth, partial anthesis, and poor flower development [13]. Therefore, insufficient chilling hours of a fruit tree in a mild-winter region could be compensated through exogenous application of some chemical products [13] such as copper sulphate (10%), zinc sulphate (5%), and urea (10%) during the period from September to January [62]. These products are commonly categorized as S-based (e.g., copper sulphate, zinc sulphate), N-based (e.g., potassium nitrate, calcium cyanamide, hydrogen cyanamide (HC), sodium azide, urea), and S- and N-based chemical components (e.g., thidiazuron) [63], of which some examples are given in Table 3.
3.2. Dormancy Breaking Organic Compounds
There are several organic compounds that can be applied to trees for dormancy breaking such as mineral oil, garlic extract, and thiourea [64,65]. Due to environmental concerns and toxicity of some chemical compounds, organic compounds have been used for breaking bud dormancy such cinnamon, coffee, ginger, clove, colocynth, olive, garlic, red chilies, nigella, and turmeric extracts [66], of which some examples are given in Table 3. These organic agents are rich in volatile compounds, pigments, tannins, phenols, antioxidants, vitamins, and nutrients [66].
The recent advancement in organic farming has increased the demand for dormancy breaking organic compounds as safe alternatives for the chemical ones. Garlic (Allium sativum L.) extract is considered as one of the most effective organic compounds for dormancy breaking [67,68,69]. The application of garlic extract either alone (5–10%) or combined with mineral oil (2–4%) was highly effective in dormancy breaking of grape, apple, and kiwifruit [67,68,69]. Foliar application of garlic extract (1%) induced 5-day-early bud break in peach [70]. The promotive effect of garlic extract is mainly related to its sulfur constituents: di-allyl-sulfides (mono-, di- and tri-), and dimethyl disulfide [68]. The application of fresh garlic extract, garlic oil, or di-allyl-sulfide stimulated dormancy breaking in grapevine without causing phytotoxicity in seedlings [68]. Garlic extract is rich in sulfur molecules, which produce cysteine as a result of reduced sulfur [69]. Cysteine metabolism leads to glutathione, which detoxifies ROS and free radicals. The reduced glutathione can promote the up-regulation of the transcription of 1,3-β-d-glucanase, which is considered as a fundamental enzyme in the mechanism of dormancy breaking [71]. The exogenous application of reduced glutathione showed promoted bud break in grapevines [72].
S-methyl cysteine sulfoxide led to 100% dormancy breaking in buds of various table grape cultivars [73]. Foliar sprays of onion extract on apple promoted bud break, reduced the number of days to full bloom, and increased the contents of total free amino acids, hydrogen peroxide, proline, auxins, and anthocyanin but reduced catalase activity and free phenol content [64].
Combinations of amino acids, polysaccharides, nitrogen, and calcium (e.g., Bluprins, Biolchim Spa, Bologna, Italy) can work as bud dormancy breaking agents [74] and can cause bud break induction and uniform flowering in grapes, cherries, and kiwifruit [75]. In a four-year comparative study, the combined solution of mineral oil (3.5%) + HC (0.35%) was more effective on bud break of apple cultivars “Maxi Gala” and “Fuji Suprema” compared to Bluprins, but all treatments showed significant results compared to the control plots [21].
Table 3.
List of some dormancy breaking compounds, their applied and effective concentrations, used crops, and literature references (Ref.).
Table 3.
List of some dormancy breaking compounds, their applied and effective concentrations, used crops, and literature references (Ref.).
| Compound | Application Time | Applied Concentrations (Effective Concentrations) | Crop (Scientific Name) | Ref. |
|---|---|---|---|---|
| Potassium nitrate (KNO3) | During germination | 0.2, 1, and 3% (0.2%) | Agrimony (Agrimonia eupatoria L.) | [76] |
| Gibberellic acid (GA3) | During germination | 100, 200, and 300 mg L−1 (100 mg L−1) | Agrimony (Agrimonia eupatoria L.) | [76] |
| Hydrogen cyanamide (HC) | During bud cutting | 5% for 4-year-old cuttings | Grapes (Vitis vinifera L. × Vitis labruscana Bailey) | [77] |
| GA3 or KNO3 | During pre-germination | Seeds soaking in GA3 (400 ppm) for 24 h or KNO3 (1 M) for 30 min | Highland papaya (Vasconcelle aquercifolia L.) | [78] |
| GA3 | During seed priming | Seeds primed in GA3 (750 ppm) for 48h | Common poppy (Papaver rhoeas L.) and (P. dubium L.) | [79] |
| KNO3 | During seed priming | Seed treatment with KNO3 (0.5 g L−1) for 24 h | ||
| GA3 or KNO3 | During germination | Dormant seeds treated with GA3 (150 ppm) or KNO3 (1000 ppm) for 24 h. | Wall rocket (Diplot axiserucoides L.) | [80] |
| Garlic extract (GE) or HC | Before winter dormancy | Cuttings immersed in GE (10%) or HC (4%) (v/v) for 10 s | Grape (Vitis vinifera L.) | [68] |
| HC (Dormex) | During bud cutting | 6-year-old cuttings treated with HC (5%) | Grape (Vitis vinifera L. × Vitis labruscana Bailey) | [13,64] |
| GA3 | During germination | Seed soaking in GA3 (10, 100, or 1000 mg L−1) and scoring at 30 d | Ashitaba Angelica keiskei | [81] |
| Zinc sulphate (ZnSO4) | Before winter dormancy | Foliar application (1000, 1500, and 2000 mg L−1) (2000 mg/L) | Kiwifruit (Actinidia deliciosa L.) | [82] |
| HC | 45 days before natural bud break | 2, 4 and 6% (4%) + mineral oil (2%) | Kiwifruit (Actinidia deliciosa L.), cv. Hayward | [83] |
| Erger Biostimulant | At week no. 5, 9, and 13 from bud break | Foliar application (6%) once at three different times in fall | Kiwifruit (Actinidia deliciosa L.) | [84] |
| KNO3 or GA3 | At stratification of seeds | Seed soaking in KNO3 (0.2%) or GA3 (5 mM) | Apple (Malus domestica L.), cv. Ligol | [85] |
| HC | In late August | Foliar application (12.5 mM) | Grapevines (V. vinifera L. x V. labruscana Bailey), cv. Kyoho | [86] |
| HC, KNO3, mineral oil, thiourea, or Ca(NO3)2 | In December | Foliar application of HC (4%), KNO3 (8%), mineral oil (6%), thiourea (2%), or Ca(NO3)2 (6%) | Apple (Malus sylvestris Mill), cv. Anna | [64] |
3.3. Some Aspects of the Mechanism of Dormancy Breaking
The seed or bud dormancy breaking mechanism is a vital biological process for the proper timing of flowering and fruiting and hence the survival of a plant species in its geographical zone [63]. Dormancy breaking in seeds is often hampered by a hard seed cover, a mechanical dormancy that greatly prevents moisture absorption, while in some other species, seeds may have undergone physiological dormancy, which hinders seed germination due to internal factors (i.e., hormonal imbalance) [87]. Seed dormancy is an adaptation strategy for several plant species to survive the unfavorable environmental conditions [88]. Bud dormancy in deciduous fruit trees is a crucial plant phase that is mainly related to cold temperature in the winter season and the accumulation of specific cold hours that eventually trigger bud growth due to thewarm temperature in spring [89]. Phytohormones such as indole-3-butyric acid (IBA), indole-3-acetic acid (IAA), gibberellins (GAs), and abscisic acid (ABA) play a crucial role in dormancy induction and breaking mechanisms of seeds and buds. During seed maturation, ABA induces dormancy, but GAs are essential in dormancy breaking [90]. The balance of both hormones and ROS regulates bud endo-dormancy, and buds resume their growth once they are exposed to sufficient chilling requirements [89,91]. Different oxidative and reductive reactions are important for dormancy breaking [92]. For example, foliar spray of HC inhibited catalase activity and led to increased levels of H2O2 that ultimately broke bud endo-dormancy [71]. Using thiourea has also showed increased levels of proline, putrescine, and biogenic amines (i.e., tryptamine, tyramine, histamine, methyl-butylamine, and serotonin), calcium, and ethylene but reduced levels of ABA that promoted bud break in apple trees [64,65,93]. Within 5 days of sprayed HC application to grapes, a dramatic rise in starch hydrolysis and soluble sugar contents in buds and internodes was noticed. The reduction in starch content was accompanied with an induction in α-amylase activity, and flowering time was dependent on the rate of sugar accumulation in buds [94]. Dormex (AlzChemTrostberg GmbH, Trostberg, Germany) can release cyanide (CN), which also increases due to the role of other enzymes (e.g., glucosidase, lyase), associated with increased levels of ROS in the cytosol that eventually affect gene expression related to dormancy breaking [22]. When chilling requirements are fulfilled, dormancy-associated (DA) transcript levels are also affected, so dormant flower buds start to accumulate heating hours, and then flowering takes place [24]. It was reported that GA4 could induce bud break in poplar (Populus balsamifera L.). Long-term chilling exposure is associated with GA biosynthesis and correlated with plant re-growth ability [95]. It was observed that GA4 application can compensate for chilling requirements and accelerate bud break in Japanese apricot through the inductive expression of important metabolites (e.g., galactose, glyoxylate, dicarboxylate, starch, sucrose) involved with energy metabolism and oxidation-reduction reactions (redox system) [96,97]. The complete series of bud dormancy starting from bud formation to flowering has been thoroughly described for various deciduous fruit crops [14,22,88,93,98,99,100,101,102,103,104,105].
4. Chilling Requirements
Chilling requirements could be defined as a certain range of hours at a specific cold temperature range required during winter for endo-dormancy breaking and re-growth by early spring [14]. The most important criteria when choosing cultivation sites in the warm winter fruit growing regions are the quality and/or quantity of chill during winter. Therefore, the cultivation of deciduous fruit trees species in a low-chill region may require the use of dormancy breaking agents, along with plant varieties that have low chilling requirements. After trees fulfill their chilling requirements and exit endo-dormancy, they also need sufficient heat hours for re-growth. Thus, it is important to find a strategy to calculate the chilling and heat requirements, and this may require experiments at a very large scale, which is impractical [29]. A very close relationship between dormancy and chilling requirements, particularly under the conditions of climate change, has been reported generally [14,106,107] but also at species level, e.g., apricot [108], apple and almond [109], sweet cherry [110] and pomegranate [111].
4.1. Chilling Requirements Versus Climate Change and Warm Winter Conditions
Many species and cultivars of deciduous fruit trees require cold temperature during winter and warm temperature for endo-dormancy breaking in spring [112]. The effective chilling temperatures range from 0 to 7 °C for most deciduous fruit. Temperatures below 0 or above 7 °C are not sufficient to fulfill plant chilling requirements [113]. The initiation of vegetative and reproductive buds starts in summer and late fall, and then buds go into dormancy during winter when day length became shorter and temperature decreases [114]. The average Earth temperature is currently higher than it was 1200 years ago [115]; therefore, global warming affects temperate and polar latitudes significantly more than the equatorial ones [23]. It is expected that average temperature will increase over 4 °C by the end of this century [116]. In this regard, it is expected that problems related to floral bud induction will be more significant in the coming years, and this requires creating a novel system to understand the mechanism that induces bud break. Different fruit crops showed fluctuation in bud break and plant phenological stages because of climate change. For example, olive in Spain and Italy showed fluctuation in flowering pattern with temperature fluctuations between cold and warm winter, and this can provoke frost damage by the beginning of spring [117,118]. Global warming has induced early flowering of pear and apple trees in France due to the change in tree response to different chilling and heating temperature patterns, which enhance the development of floral primordia within buds and induce bud break [25]. The significant effect of global warming has been more pronounced by the end of winter and beginning of spring (February–March) since 1988. During this time, the mean temperature increased by 1.6 °C during early spring, and 0.8 °C during winter (November–December) [25]. Warm winter has extended the bud dormancy period causing fluctuation in bud break and phenological stages. This has affected flowering time and duration, pollen grain quality, pistil malformation, fruit yield, and fruit quality. Fruit size and weight were higher during seasons with cold winters, since fruit were subjected to sufficient chill [29]. In temperate climates, spring frost is considered the most important factor of crop damage. A temperature decline below zero for a few hours was sufficient to kill apricot flower buds in Hungary. In addition, early flowering resulted in floral damage and low fruit quality [119]. In the subtropics of India where there is insufficient chill during winter, most pear orchards remained unproductive even after plants reached maturation [120]. In general, global warming leads to less vigor in plants and reduction in fruit set, yield, size, color, juice content, and shelf life with increased pest infestation in apple [114,121]. Continuous insufficient chilling leads to physiological symptoms such as delayed foliation, low yield, and fruit physiological disorders [122]. Long drought periods during summer and reduced snowfall during winter have decreased the suitable area for apple cultivation, which has changed the flora in such areas because growers shifted to other cash crops [114]. This has also increased the need to replace temperate fruit species and cultivars grown in tropical and subtropical areas with others that have low chilling requirements [123]. Mild winter (subtropical) regions in the middle latitudes, which include the transition from tropical to temperate zones, are dominant areas for stone fruit production. Trees grown in mild winters may have fewer chilling units in some years and may also be damaged by spring frost in others. Therefore, these regions of mild winter have been classified into three zones according to chilling requirements: transitional zone (650–950 chilling units), medium-chill zone (400–650 chilling units), and low-chill zone (below 400 chilling units) [124]. In Egypt, apple trees that were artificially chill-treated at 7 or 10 °C continuously took about 600 hours for bud break [125]. Therefore, winter warming may increase heat accumulation rate, which could compensate for any phenology-delaying impacts of reduced chilling [107].
4.2. Models to Calculate Chilling Requirements
Many mathematical models have been developed to determine chilling requirements for different fruit tree species and cultivars in various growing regions (Table 4). The approaches of these models make them reliable and plausible, although they are not suitable for all growing regions. An accurate model is essential for the quantitative development of suitable adaptation strategies of climate change for temperate fruit trees [126]. The use of many mathematical models is dependent on the plant species and growing regions, and new models are constructed for various climatic conditions (Table 4).
Simple classical models include the growing degree hours (GDH) model to calculate heat requirement [127]. Some other models simply count the “chilling hours” during winter by calculating hours below 7.2 or between 0 and 7.2 °C. However, these models neglect the unfavorable effect of warm winter temperatures on chilling-hour accumulation, especially in subtropical or tropical climates. This error was corrected by the “Utah model” for peach [128] and by using advanced statistical approaches, such as R language, to facilitate and standardize the use of various models [6]. Many other models were also developed to calculate winter chill. The time at which trees were exposed to cold temperature can be used to estimate the approximate date of dormancy breaking. The importance of predicting climate changes and their impact on available chilling hours is critical for deciduous fruit because well-established orchards are planted for many years, and it will take about 15–30 years obtain a full return when replacing these orchards with new cultivars with fewer chilling requirements [129]. In addition, breeding programs may also take several years to develop such new cultivars. Moreover, the qualitative characteristics of a new cultivar are the result of many factors (e.g., eco-pedology, cultivation methods, and cultural and socioeconomic parameters) that eventually affect its success in a specific area [28].
Chilling models use climate change scenarios to predict future chilling hours in a specific region. Knowledge of plant chilling requirements can be used in breeding programs to evaluate plant capability to adapt in different climates [28]. The outcomes of chilling models are largely dependent on the type of used model and the conditions of the growing region [11]. The choice of appropriate model is important for the determination of climatic change impacts on winter chill [130]. To determine the chilling requirements of a particular cultivar, two steps can be conducted (Figure 2). The first step is to establish a dormancy phase, and this can be carried out by running experiments or by statistical analyses. The second step is to calculate the chilling requirements using temperature-based models.
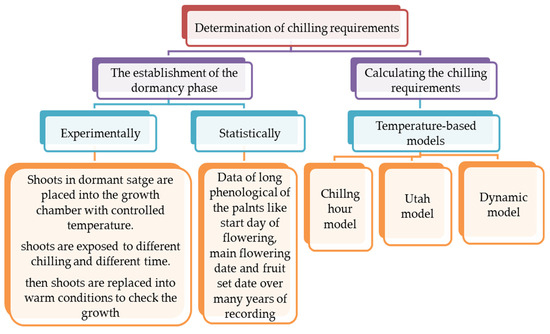
Figure 2.
A chart flow for determination of chilling requirements for deciduous fruit trees.
Table 4.
Names, descriptions and literature references (Ref.) for models to calculate chilling requirements.
Table 4.
Names, descriptions and literature references (Ref.) for models to calculate chilling requirements.
| Model Name | Description | Ref. |
|---|---|---|
| Chilling hours | Simple and widely used. Counts the number of hours below 7.2 °C. | [127] |
| Utah | Uses temperatures that trees are exposed to, expressed as chilling units (CU): 1 CU for 2.5–9.1 °C, 0.5 CU for 9.2–12.4 °C, 0 CU for <1.4 or 12.5–15.9 °C, −0.5 CU for 16–18 °C, and -1 CU for >18 °C. | [128] |
| Landsberg | Uses CU based on dividing the daily temperature by a base temperature of the crop. If the base temperature is 5 °C and the daily temperature is 5 °C, then the chilling unit is 1. | [131] |
| Low Chill | Developed for the low chilling requirements of the ‘Sungold’ nectarine, it can be used for low-chilling-requirement cultivars in subtropics. Uses CU of 1 for 1.8–8 °C, 0 for <−1 and 14–17 °C, and −1 for >19.5 °C. | [132] |
| North Carolina | Uses CU of 1 for 1.6–7.2 °C, 0 for <−1.1 °C and 16.5–19 °C, and −2 for >23.3 °C. | [133] |
| Dynamic | Proposes an intermediate product formed in buds due to cold temperature that can be reversed by warm temperature. If this product reaches a certain concentration at cold temperature, the chill portions are permanently fixed, and they are considered unaffected by warm temperature. One chill portion (CP) is an exposure to 6 °C for 28 hours. | [134] |
| Positive Chill Units (PCU) | A modified version of the Utah model to avoid negative values at warm temperature. CU = 0 is used for >15.9 °C. | [135] |
| ‘Sweetheart’ cherry and ‘Gala’ apple | Uses CU = 1 for −2.1–7 °C (cherry) and −2.1–5.5 °C (apple), −1 for >18 °C (cherry and apple), and 0 for 13–16 °C (cherry and apple). | [136] |
| Jones I | Was used to study the chilling requirements of 20 blackcurrant genotypes. The model depends on a decreasing exponential curve by fitting a curve with the response to chilling temperature. The decreasing exponential = 1 CU for −2.7 °C, 0.5 CU for 2.0 °C, 0.25 CU for 6.6 °C, and 0.1 CU for 12.8 °C. | [137] |
| Jones II | Winter chill data of 0–7.2 °C (denoted by 0–7.2 CU) calculated by the summation of total chill hours from October 1st in a relevant year when the mean temperature was 0–7.2 °C. Similarly, <7.2 °C (denoted by 7.2 CU) for mean temperature <7.2 °C, and exponential units (denoted by “exp”), where exp is the summation of hours from 1 October for the function exp = 0.6702 [exp (−0.148 × Ta)], where Ta is the hourly measured mean temperature. | [138] |
| Luedeling: ’chillR’ | “chillR” contains functions for processing temperature records into chilling models (chilling hours, Utah, and dynamic models) and heat units (growing degree hours). | [139] |
The major three models are the “chilling hours”, “Utah” and “dynamic” models. The chilling hours and Utah models were applied in the temperate regions successfully, but they were less efficient in the Mediterranean and subtropical regions under warm winter conditions [140].
The dynamic model philosophy is dependent upon chilling requirements being determined in two steps started by the accumulation of an intermediate product that is promoted by cold temperature but may be reversed with warmer temperature [134]. The second step includes a certain level of this product that is irreversible even in warm temperature. Therefore, the same temperature should have the same effect using either the chilling hours or Utah model; however, the effect of the same temperature is quite different, since the dynamic model depends on the time the temperature was recorded [60,126]. These three models perform in different ways related to the variability of calculated chilling requirements from one year to another. The dynamic model showed the least variation (0.1–5.9%), followed by the chilling hours model (1.3–9.0%) and the Utah model (2.1–14.0%). Therefore, the dynamic model data were the most preferred to predict winter chill under warm winter conditions [140]. The impacts of climate change in Argentina were studied in vineyards using some bioclimatic indices that characterized the potential climate changes, which were highly related to the qualitative potential of grapes. Four bioclimatic indices have been used: (i) average growing-season temperature (GST), (ii) cool night index (CNI), (iii) average growing-season precipitation (GSP), and (iv) average monthly minimum temperature (DSTmin) during the dormant season [129]. In Argentina, the viticulture industry may face challenges in warm climate conditions. The cold climate regions in Argentina are favored for grapes either in near- or distant-future production plans, but warm regions have disadvantages for grape production, especially plans in the distant future (i.e., 2075–2099) under climate change scenarios. This means that the warm climate regions may be suitable for current grape cultivar growth or for cultivating new cultivars with fewer chilling requirements [129]. In Morocco, the reduction in chill hours during warm winter resulted in many phenological interruptions such as delayed flowering and/or prolonged flowering period, along with low fruit quality in apple. In seasons when trees had received enough chilling, fruit quality showed better results, especially for fruit weight, size, and firmness. The chilling requirements of apple were 645–677 chill hours using the chilling-hour model, 709–1157 chill units (CU) using the Utah model, and 43.4–55.2 chill portions (CP) using the dynamic model. To maintain adequate apple fruit production in Morocco, the replacement of the current varieties with other varieties that require fewer chilling hours is necessary [29]. Some other findings have also confirmed this conclusion [29,44,130,141,142,143,144].
5. Conclusions
As mentioned above, climate change is a generally a great challenge for temperate fruit tree production. The key factors of climate change are the elevated mean temperature and atmospheric CO2 [1,5]. These changes negatively impact, for instance, the chilling requirements, dormancy, and phenology of temperate fruit trees [127,143,145], especially in growing areas with warm winter conditions where trees cannot fulfill their chilling requirements for dormancy breaking and re-growth by spring. Global warming has led to chill deficit during winter, which may become a limiting factor for growing temperate fruit trees in the tropics and subtropics. Therefore, several dormancy breaking options have been used to compensate chill deficit and induce growth, flowering, and fruiting while maintaining the phenological sequences of the tree. In addition, several models have also been proposed in order to calculate and estimate the required chill conditions in specific regions for temperate fruit tree species and to evaluate the sustainability of a cultivar in a specific region. However, the current status of research on dormancy and chilling under warm winter growing conditions is incomplete, and many scientific problems are unsolved. For example, with the increasing rates of climate change,
- New and more effective dormancy breaking organic compounds have to be discovered;
- New climate-smart cultivars have to be bred in order to solve the increasing difficulties for dormancy and chilling requirements [146], i.e., a longer period from the fulfillment of chilling requirements to bud break. This would help to escape spring cold events in some years in Mediterranean areas;
- Dormancy and chilling forecasting models have to be improved further in order to successfully manage temperate fruit tree production worldwide but especially in the tropical and subtropical regions.
If these research issues are successfully addressed in the near future, temperate fruit tree plantations can be protected for a long period, especially in the warm winter regions.
Author Contributions
This review was mainly organized by A.-M.S., A.E., H.E.-R., S.M.A.-E., S.K.O., H.M.E., I.F.H. and I.J.H., who contributed to material collection and writing and revision of the manuscript. The original manuscript was written by A.-M.S., A.E., and I.J.H. A.I. read and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by the NKFI K131478 and the Thematic Excellence Programme (TKP2020-NKA-04) of the Ministry for Innovation and Technology in Hungary projects.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors would like to thank Hatem Kotb, Deciduous Fruit Department, Horticulture Research Institute Agricultural research Center, Egypt, for providing the pictures of some developmental stages of deciduous fruit trees.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef]
- Boadu, F.O. Climate change. In Agricultural Law and Economics in Sub-Saharan Africa Cases and Comments; Boadu, F.O., Ed.; Elsevier Inc.: Dordrecht, The Netherlands, 2016; pp. 555–571. [Google Scholar] [CrossRef]
- Miranda, C.; Urrestarazu, J.; Santesteban, L.G. Fruclimadapt: An R package for climate adaptation assessment of temperate fruit species. Comp. Electr. Agric. 2021, 180, 105879. [Google Scholar] [CrossRef]
- Ahmed, N.; Lone, F.A.; Hussain, K.; Kanth, R.H.; Mahdi, S.S. Impact of Climate change on temperate fruit production in Kashmir Valley, North Western Himalayan Region of India—Challenges, opportunities and way forward. In Climate Change and Agriculture in India: Impact and Adaptation; Sheraz Mahdi, S., Ed.; Springer International Publishing AG: Cham, Switzerland, 2019; pp. 251–262. [Google Scholar] [CrossRef]
- Anon. 2020. Earth’s CO2 Home Page. Available online: https://www.co2.earth (accessed on 25 December 2020).
- Pio, R.; de Souza, F.B.M.; Kalcsits, L.; Bisi, R.B.; Farias, D.H. Advances in the production of temperate fruits in the tropics. Acta Sci. 2019, 41, e39549. [Google Scholar] [CrossRef]
- Rodríguez, A.; Pérez-López, D.; Centeno, A.; Ruiz-Ramos, M. Viability of temperate fruit tree varieties in Spain under climate change according to chilling accumulation. Agric. Syst. 2021, 186, 102961. [Google Scholar] [CrossRef]
- Yahia, E.M. Classification of horticultural commodities. In Postharvest Technology of Perishable Horticultural Commodities; Yahia, E.M., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2019; pp. 71–97. [Google Scholar] [CrossRef]
- FAO Faostat: Statistical Database. Food and Agriculture Organization of the United Nation, Rome, Italy. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 2 May 2019).
- Rai, R.; Joshi, S.; Roy, S.; Singh, O.; Samir, M.; Chandra, A. Implications of changing climate on productivity of temperate fruit crops with special reference to apple. J. Hortic. 2015, 2, 2. [Google Scholar] [CrossRef]
- Fadón, E.; Fernandez, E.; Behn, H.; Luedeling, E. A conceptual framework for winter dormancy in deciduous trees. Agronomy 2020, 10, 241. [Google Scholar] [CrossRef]
- Campoy, J.A.; Ruiz, D.; Egea, J. Dormancy in temperate fruit trees in a global warming context: A review. Sci. Hortic. 2011, 130, 357–372. [Google Scholar] [CrossRef]
- Khalil-Ur-Rehman, M.; Wang, W.; Dong, Y.; Faheem, M.; Xu, Y.; Gao, Z.; Tao, J. Comparative transcriptomic and proteomic analysis to deeply investigate the role of hydrogen cyanamide in grape bud dormancy. Int. J. Mol. Sci. 2019, 20, 3528. [Google Scholar] [CrossRef]
- Hernandez, J.A.; Díaz-Vivancos, P.; Martínez-Sanchez, G.; Alburquerque, N.; Martínez, D.; Barba-Espín, G.; Acosta-Motos, J.R.; Carrera, E.; García-Brunton, J. Physiological and biochemical characterization of bud dormancy: Evolution of carbohydrate and antioxidant metabolisms and hormonal profile in a low chill peach variety. Sci. Hortic. 2021, 281, 109957. [Google Scholar] [CrossRef]
- Vegis, A. Dormancy in higher plants. Annu. Rev. Plant Physiol. 1964, 15, 185–224. [Google Scholar] [CrossRef]
- Chuine, I.; Régnière, J. Process-based models of phenology for plants and animals. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 159–182. [Google Scholar] [CrossRef]
- Zhao, H.; Fu, Y.H.; Wang, X.; Zhang, Y.; Liu, Y.; Janssens, I.A. Diverging models introduce large uncertainty in future climate warming impact on spring phenology of temperate deciduous trees. Sci. Total Environ. 2021, 757, 143903. [Google Scholar] [CrossRef]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endodormancy, paradormancy, and ecodormancy—Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar]
- Bielenberg, D.G.; Wang, Y.E.; Li, Z.; Zhebentyayeva, T.; Fan, S.; Reighard, G.L.; Scorza, R.; Abbott, A.G. Sequencing and annotation of the evergrowing locus in peach [Prunus persica (L.) Batsch] reveals a cluster of six MADS-box transcription factors as candidate genes for regulation of terminal bud formation. Tree Genet. Genom. 2008, 4, 495–507. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhuo, X.; Zhao, K.; Zheng, T.; Han, Y.; Yuan, C.; Zhang, Q. Transcriptome profiles reveal the crucial roles of hormone and sugar in the bud dormancy of Prunus mume. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Fenili, C.L.; Petri, J.L.; Sezerino, A.A.; De Martin, M.S.; Gabardo, G.C.; Daniel, E.S. Bluprins® as alternative bud break promoter for ‘Maxi Gala’ and ‘Fuji Suprema’ apple trees. J. Exp. Agric. Internat. 2018, 26, 1–13. [Google Scholar] [CrossRef]
- Beauvieux, R.; Wenden, B.; Dirlewanger, E. Bud dormancy in perennial fruit tree species: A pivotal role for oxidative cues. Front. Plant Sci. 2018, 9, 657. [Google Scholar] [CrossRef]
- AR4 Climate Change. The physical science basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; pp. 1–450. [Google Scholar]
- Luedeling, E.; Girvetz, E.H.; Brown, P.H. Climate change affects winter chill for temperate fruit and nu tress. PLoS ONE 2011, 6, e20155. [Google Scholar] [CrossRef]
- Guédon, Y.; Legave, J.M. Analyzing the time-course variation of apple and pear tree dates of flowering stages in the global warming context. Ecol. Model. 2008, 219, 189–199. [Google Scholar] [CrossRef]
- Costa, C.; Stassen, P.J.C.; Mudzunga, J. Chemical rest breaking agents for the South African pome and stone fruit industry. Acta Hortic. 2004, 636, 295–302. [Google Scholar] [CrossRef]
- Erez, A. Means to compensate for insufficient chilling to improve bloom and leafing. Acta Hortic. 1995, 395, 81–95. [Google Scholar] [CrossRef]
- Fadón, E.; Herrera, S.; Guerrero, B.I.; Guerra, M.E.; Rodrigo, J. Chilling and heat requirements of temperate stone fruit trees (Prunus sp.). Agronomy 2020, 10, 409. [Google Scholar] [CrossRef]
- El Yaacoubi, A.; El Jaouhari, N.; Bourioug, M.; El Youssfi, L.; Cherroud, S. Potential vulnerability of Moroccan apple orchard to climate change – induced phenological perturbations: Effects on yields and fruit quality. Int. J. Biometeorol. 2020, 64, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, P.; Bhaskar, R.; Papnai, G.; Joshi, N.; Supyal, V. Impact of climate change on apple phenology and adaptability of Anna variety (low chilling cultivar) in lower hills of Uttarakhand. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 453–460. [Google Scholar] [CrossRef]
- Díez-Palet, I.; Funes, I.; Savé, R.; Biel, C.; Herralde, F.; Miarnau, X.; Vargas, F.; Àvila, G.; Carbó, J.; Aranda, X. Blooming under Mediterranean climate: Estimating cultivar-specific chill and heat requirements of almond and apple trees using a statistical approach. Agronomy 2019, 9, 760. [Google Scholar] [CrossRef]
- Pfleiderer, P.; Menke, I.; Schleussner, C. Increasing risks of apple tree frost damage under climate change. Clim. Chang. 2019, 57, 515–525. [Google Scholar] [CrossRef]
- Ahmadi, H.; Ghalhari, G.F.; Baaghideh, M. Impacts of climate change on apple tree cultivation areas in Iran. Clim. Chang. 2019, 153, 91–103. [Google Scholar] [CrossRef]
- Wolfe, D.W.; De Gaetano, A.T.; Peck, G.M.; Carey, M.; Ziska, L.H.; Lea-Cox, J.; Kemanian, A.R.; Hoffmann, M.P.; Hollinger, D.Y. Unique challenges and opportunities for northeastern US crop production in a changing climate. Clim. Chang. 2018, 146, 231–245. [Google Scholar] [CrossRef]
- Funes, I.; Aranda, X.; Biel, C.; Carbó, J.; Camps, F.; Antonio, J.; Molina de Herralde, F.; Graua, B.; Savé, R. Future climate change impacts on apple flowering date in a Mediterranean subbasin. Agri. Water Manag. 2016, 164, 19–27. [Google Scholar] [CrossRef]
- Chand, H.; Verma, S.C.; Bhardwaj, S.K.; Sharma, S.D.; Mahajan, P.K.; Sharma, R. Effect of changing climatic conditions on chill units accumulation and productivity of apple in mid hill sub humid zone of Western Himalayas, India. Curr. World Environ. 2016, 11, 142–149. [Google Scholar] [CrossRef][Green Version]
- Antolín, M.C.; Toledo, M.; Pascual, I.; Irigoyen, J.J.; Goicoechea, N. The Exploitation of local Vitis vinifera L. biodiversity as a valuable tool to cope with climate change maintaining berry quality. Plants 2021, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, S.; Fattahi, M.; Bedis, K.; Nasrolahpour-moghadam, S.; Irigoyen, J.J.; Gogorcena, Y. Interactional effects of climate change factors on the water status, photosynthetic rate, and metabolic regulation in peach. Front. Plant Sci. 2020, 11, 43. [Google Scholar] [CrossRef]
- Arrizabalaga-Arriazu, M.; Gomès, E.; Morales, F.; Irigoyen, J.J.; Pascual, I.; Hilbert, G. High temperature and elevated carbon dioxide modify berry composition of different clones of grapevine (Vitis vinifera L.) cv. Tempranillo. Front. Plant Sci. 2020, 11, 603687. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Lüscher, J.; Kizildeniz, T.; Vucetic, V.; Dai, Z.; Luedeling, E.; van Leeuwen, C.; Gomès, E.; Pascual, I.; Irigoyen, J.J.; Morales, F.; et al. Sensitivity of grapevine phenology to water availability, temperature and CO2 concentration. Front. Environ. Sci. 2016, 4, 48. [Google Scholar] [CrossRef]
- Salazar-Parra, C.; Aranjuelo, I.; Pascual, I.; Erice, G.; Sanz-Sáez, Á.; Aguirreolea, J.; Sánchez-Díaz, M.; Irigoyen, J.J.; Araus, J.L.; Morales, F. Carbon balance, partitioning and photosynthetic acclimation in fruit-bearing grapevine (Vitis vinifera L. cv. Tempranillo) grown under simulated climate change (elevated CO2, elevated temperature and moderate drought) scenarios in temperature gradient greenhouses. J. Plant Physiol. 2015, 174, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Xi, W.; Zhang, Q.; Lu, X.; Wei, C.; Yu, S.; Zhou, Z. Improvement of flavour quality and consumer acceptance during postharvest ripening in greenhouse peaches by carbon dioxide enrichment. Food Chem. 2014, 164, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Han, J.-H.; Cho, J.G.; Son, I.-C.; Kim, S.H.; Lee, I.-B.; Choi, I.M.; Kim, D. Effects of elevated carbon dioxide and temperature on photosynthesis and fruit characteristics of ‘Niitaka’ pear (Pyrus pyrifolia Nakai). Hortic. Environ. Biotechnol. 2012, 53, 357–361. [Google Scholar] [CrossRef]
- Chitu, E.; Paltineanu, C. Timing of phenological stages for apple and pear trees under climate change in a temperate-continental climate. Int. J. Biometeorol. 2020, 64, 1263–1271. [Google Scholar] [CrossRef]
- Schattman, R.E.; Niles, M.T.; Aitken, H.M. Water use governance in a temperate region: Implications for agricultural climate change adaptation in the Northeastern United States. Ambio 2020, 50, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.; Fu, Y.H.; Hao, F.; Zhou, X.; Zhang, X.; Yin, G.; Vitasse, Y.; Piao, S.; Niu, K.; De Boeck, H.J. Climate warming increases spring phenological differences among temperate trees. Global Chang. Biol. 2020, 26, 5979–5987. [Google Scholar] [CrossRef]
- Haokip, S.W.; Shankar, K.; Lalrinngheta, J. Climate change and its impact on fruit crops. J. Pharmacogn. Phytochem. 2020, 9, 435–438. [Google Scholar]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine responses to heat stress and global warming. Plants 2020, 9, 1754. [Google Scholar] [CrossRef] [PubMed]
- Naulleau, A.; Gary, C.; Prévot, L.; Hossard, L. Evaluating strategies for adaptation to climate change in grapevine production–A systematic review. Front. Plant Sci. 2021, 11, 607859. [Google Scholar] [CrossRef]
- Parker, L.; Pathak, T.; Ostoja, S. Climate change reduces frost exposure for high-value California orchard crops. Sci. Total Environ. 2021, 762, 143971. [Google Scholar] [CrossRef]
- Jha, P.K.; Materia, S.; Zizzi, G.; Costa-Saura, J.M.; Trabucco, A.; Evans, J.; Bregaglio, S. Climate change impacts on phenology and yield of hazelnut in Australia. Agric. Syst. 2021, 186, 102982. [Google Scholar] [CrossRef]
- Vanalli, C.; Casagrandi, R.; Gatto, M.; Bevacqua, D. Shifts in the thermal niche of fruit trees under climate change: The case of peach cultivation in France. Agric. For. Meteorol. 2021, 300, 108327. [Google Scholar] [CrossRef]
- Zahoor, S.; Dutt, V.; Mughal, A.H.; Pala, N.A.; Qaisar, K.N.; Khan, P.A. Apple-based agroforestry systems for biomass production and carbon sequestration: Implication for food security and climate change contemplates in temperate region of Northern Himalaya, India. Agroforest Syst. 2021, 95, 367–382. [Google Scholar] [CrossRef]
- Anderson, J.V. Advances in Plant Dormancy; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–325. [Google Scholar] [CrossRef]
- Tanner, J.D.; Minas, I.S.; Chen, K.Y.; Jenderek, M.M.; Wallner, S.J. Antimicrobial forcing solution improves recovery of cryopreserved temperate fruit tree dormant buds. Cryobiology 2020, 92, 241–247. [Google Scholar] [CrossRef]
- Rojo, J.; Orlandi, F.; Ben Dhiab, A.; Lara, B.; Picornell, A.; Oteros, J.; Msallem, M.; Fornaciari, M.; Pérez-Badia, R. Estimation of chilling and heat accumulation periods based on the timing of olive pollination. Forests 2020, 11, 835. [Google Scholar] [CrossRef]
- Melke, A. The physiology of chilling temperature requirements for dormancy release and bud-break in temperate fruit trees grown at mild winter tropical climate. J. Plant Stud. 2015, 4, 110–156. [Google Scholar] [CrossRef]
- Vitasse, Y.; Schneider, L.; Rixen, C.; Christen, D.; Rebetez, M. Increase in the risk of exposure of forest and fruit trees to spring frosts at higher elevations in Switzerland over the last four decades. Agric. For. Meteorol. 2018, 248, 60–69. [Google Scholar] [CrossRef]
- Oliveira, I.V.M.; Lopes, P.R.C.; Silva Matos, R.R.S.; Cavalcante, I.H.L. Apple phenology, cv. ‘Countess’ in the San Francisco valley. J. Agric. Sci. 2013, 36, 23–30. (In Portuguese) [Google Scholar]
- Fadón, E.; Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environ. Exp. Bot. 2018, 152, 28–36. [Google Scholar] [CrossRef]
- Prudencio, A.S.; Díaz-Vivancos, P.; Dicenta, F.; Hernandez, J.A.; Martínez-Gomez, P. Monitoring the transition from endodormancy to ecodormancy in almond through the analysis and expression of a specific Class III peroxidase gene. Tree Genet. Genomes 2019, 15, 44. [Google Scholar] [CrossRef]
- Singh, H. Use of chemicals in fruit crops for dormancy induction: A Review. Int. J. Ecol. Environ. Sci. 2020, 2, 249–250. [Google Scholar]
- Ionescu, I.A.; Moller, B.L.; Sánchez-Pérez, R. Chemical control of Flowering time. J. Exp. Bot. 2017, 68, 369–382. [Google Scholar] [CrossRef]
- Seif El-Yazal, M.A.S.; El-Yazal, S.A.S.; Rady, M.M. Exogenous dormancy breaking substances positively change endogenous phytohormones and amino acids during dormancy release in ‘Anna’ apple trees. Plant Growth Regul. 2014, 72, 211–220. [Google Scholar] [CrossRef]
- Seif El-Yazal, M.A.S.; Rady, M.M. Trees foliar-applied Dormex™ or thiourea-enhanced proline and biogenic amine contents and hastened breaking bud dormancy in “Ain Shemer” apple. Trees 2013, 27, 161–169. [Google Scholar] [CrossRef]
- Ahmed, F.F.; Ibrahim, H.I.M.; Abada, M.A.M.; Osman, M.M.M. Using plant extracts and chemical rest breakages for breaking bud dormancy and improving productivity of superior grapevines growing under hot climates. World Rural Obs. 2014, 6, 8–18. [Google Scholar]
- Rady, M.M.; El-Yazal, M.A.S. Garlic extract as a novel strategy to hasten dormancy release in buds of ‘Anna’ apple trees. S. Afr. J. Bot. 2014, 92, 105–111. [Google Scholar] [CrossRef]
- Orrantia-Araujo, M.A.; Martínez-Téllez, M.A.; Corrales-Maldonado, C.; Rivera-Domínguez, M.; Vargas-Arispuro, I. Changes in glutathione and glutathione disulfide content in dormant grapevine buds treated with garlic compound mix to break dormancy. Sci. Hortic. 2019, 246, 407–410. [Google Scholar] [CrossRef]
- dos Santos, R.F.; Marques, L.O.D.; Mello-Farias, P.; Martins, C.R.; Konzen, L.H.; Carvalho, I.R.; Malgarim, M.B. Bud break induction in kiwifruit vines cultivated in an organic system by the biological method of single node cutting. Bragantia-Campinas 2020, 79, 260–267. [Google Scholar] [CrossRef]
- Kotb, H.R.M.; El-Abd, M.A.M.; Salama, A. Response of “White Robin” peach trees cv. (Prunus persica L.) to cultivation under plastic covering conditions and foliar application by hydrogen cyanamide and garlic extract. J. Plant Prod. 2019, 10, 1187–1194. [Google Scholar] [CrossRef]
- Pérez, F.J.; Vergara, R.; Or, E. On the mechanism of dormancy release in grapevine-buds: A comparative study between hydrogen cyanamide and sodium azide. Plant Growth Regul. 2009, 59, 145–152. [Google Scholar] [CrossRef]
- Tohbe, M.; Mochioka, R.; Horiuchi, S.; Ogata, T.; Shiozaki, S.; Kurooka, H. Roles of ACC and glutathione during breaking of dormancy in grapevine buds by high temperature treatment. Engei Gakkai Zasshi 1998, 67, 897–901. [Google Scholar] [CrossRef]
- Vargas-Arispuro, I.; Corrales-Maldonado, C.; Martínez-Téllez, M.Á. Compounds derived from garlic as bud induction agent in organic farming of table grape. Chil. J. Agric. Res. 2008, 68, 94–101. [Google Scholar] [CrossRef]
- Ziosi, V.; Giovannetti, G.; Vitali, F.; Di Nardo, A.; Costa, G.; Laghezza, L. Bluprins® induces advanced bud release from dormancy in sweet cherry. Acta Hortic. 2013, 1009, 71–77. [Google Scholar] [CrossRef]
- Ziosi, V.; Di Nardo, A.; Fontana, A.; Vitali, F.; Costa, G. Effect of Bluprins® application on bud release from dormancy in kiwifruit, cherry, and table grape. In Advances in Plant Dormancy; Anderson, J.V., Ed.; Springer International Publishing AG: Cham, Switzerland, 2015; pp. 301–308. [Google Scholar]
- Saffari, P.; Majd, A.; Jonoubi, P.; Najafi, F. Effect of treatments on seed dormancy breaking, seedling growth, and seedling antioxidant potential of Agrimonia eupatoria L. J. Appl. Res. Medic. Aromat. Plants 2021, 20, 100282. [Google Scholar] [CrossRef]
- Khalil, U.; Rehman, K.; Wang, W.; Zheng, H.; Faheem, M.; Iqbal, S.; Shen, Z.G.; Tao, J. Role of hydrogen cyanamide (HC) in grape bud dormancy release: Proteomic approach. Biotech 2020, 10, 229. [Google Scholar] [CrossRef]
- Urtasun, M.M.; Giamminola, E.M.; Baskin, C.C.; de Viana, M.L.; Morandini, M.N.; Lamas, C.Y. Dormancy release, germination and ex situ conservation of the southern highland papaya (Vasconcellea quercifolia, Caricaceae), a wild crop relative. Sci. Hortic. 2020, 263, 109134. [Google Scholar] [CrossRef]
- Golmohammadzadeh, S.; Zaefarian, F.; Rezvani, M. Priming techniques, germination and seedling emergence in two Papaver species (P. rhoeas L. and P. dubium L., Papaveraceae). Braz. J. Bot. 2020, 43, 503–512. [Google Scholar] [CrossRef]
- Guijarro-Real, C.; Adalid-Martínez, A.M.; Gregori-Montaner, A.; Prohens, J.; Rodríguez-Burruezo, A.; Fita, A. Factors affecting germination of Diplotaxis erucoides and their effect on selected quality properties of the germinated products. Sci. Hortic. 2020, 261, 109013. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Walck, J.L.; Tao, J. Non-deep simple morphophysiological dormancy in seeds of Angelica keiskei (Apiaceae). Sci. Hortic. 2019, 255, 202–208. [Google Scholar] [CrossRef]
- Vajari, M.A.; Moghadam, J.F.; Eshghi, S. Influence of late season foliar application of urea, boric acid and zinc sulfate on nitrogenous compounds concentration in the bud and flower of Hayward kiwifruit. Sci. Hortic. 2018, 242, 137–145. [Google Scholar] [CrossRef]
- Babita Rana, V.S. Influence of Dormex on the phenology of flowering and fruit production of Hayward kiwifruit. Int. J. Chem. Stud. 2018, 6, 2941–2945. [Google Scholar]
- Hoeberichts, F.A.; Povero, G.; Ibañez, M.; Strijker, A.; Pezzolato, D.; Mills, R.; Piaggesi, A. Next Generation Sequencing to characterise the breaking of bud dormancy using a natural biostimulant in kiwifruit (Actinidia deliciosa). Sci. Hortic. 2017, 225, 252–263. [Google Scholar] [CrossRef]
- Grzesik, M.; Górnik, K.; Janas, R.; Lewandowki, M.; Romanowska-Duda, Z.; van Duijn, B. High efficiency stratification of apple cultivar Ligol seed dormancy by phytohormones, heat shock and pulsed radio frequency. J. Plant Physiol. 2017, 219, 81–90. [Google Scholar] [CrossRef]
- Sudawan, B.; Chang, C.-S.; Chao, H.-F.; Ku, M.S.B.; Yen, Y.-F. Hydrogen cyanamide breaks grapevine bud dormancy in the summer through transient activation of gene expression and accumulation of reactive oxygen and nitrogen species. BMC Plant Biol. 2016, 16, 202. [Google Scholar] [CrossRef]
- Cho, J.S.; Jang, B.K.; Lee, C.H. Breaking combinational dormancy of Rhus javanica L. seeds in South Korea: Effect of mechanical scarification and cold-moist stratification. S. Afr. J. Bot. 2020, 133, 174–177. [Google Scholar] [CrossRef]
- Kimura, E.; Fransen, S.C.; Collins, H.P.; Guy, S.O.; Johnston, W.J. Breaking seed dormancy of switchgrass (Panicum virgatum L.): A review. Biomass Bioenergy 2015, 80, 94–101. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, X.; Gao., Y.; Li, K.; Guo, W. Transcriptome-based identification of AP2/ERF family genes and their cold-regulated expression during the dormancy phase transition of Chinese cherry flower buds. Sci. Hortic. 2021, 275, 109666. [Google Scholar] [CrossRef]
- Zhang, K.; Yao, L.; Zhang, Y.; Baskin, J.M.; Baskin, C.C.; Xiong, Z.; Tao, J. A review of the seed biology of Paeonia species (Paeoniaceae), with particular reference to dormancy and germination. Planta 2019, 249, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Svystun, T.; Al-Dahmash, B.; Jonsson, A.M.; Bhalerao, R.P. Photoperiod- and temperature-mediated control of phenology in trees—A molecular perspective. New Phytol. 2017, 213, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Considine, M.J.; Foyer, C.H. Redox regulation of plant development. Antioxid. Redox Signal. 2014, 21, 1305–1326. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Halaly, T.; Acheampong, A.K.; Takebayashi, Y.; Jikumaru, Y.; Kamiya, Y.; Or, E. Abscisic acid (ABA) regulates grape bud dormancy, and dormancy release stimuli may act through modification of ABA metabolism. J. Exp. Bot. 2015, 66, 1527–1542. [Google Scholar] [CrossRef]
- Ben Mohamed, H.; Vadel, A.M.; Geuns, J.M.C.; Khemira, H. Carbohydrate changes during dormancy release in Superior Seedless grapevine cuttings following hydrogen cyanamide treatment. Sci. Hortic. 2012, 140, 19–25. [Google Scholar] [CrossRef]
- Rinne, P.L.H.; Welling, A.; Vahala, J.; Ripel, L.; Ruonala, R.; Kangasjärvi, J.; van der Schoot, C. Chilling of dormant buds hyperinduces flowering locus T and recruits GA-inducible 1, 3-β-glucanases to reopen signal conduits and release dormancy in Populus. Plant Cell 2011, 23, 130–146. [Google Scholar] [CrossRef]
- Zhuang, W.; Gao, Z.; Wang, L.; Zhong, W.; Ni, Z.; Zhang, Z. Comparative proteomic and transcriptomic approaches to address the active role of GA4 in Japanese apricot flower bud dormancy release. J. Ex. Bot. 2013, 64, 4953–4966. [Google Scholar] [CrossRef]
- Zhuang, W.; Gao, Z.; Wen, L.; Huo, X.; Cai, B.; Zhang, Z. Metabolic changes upon flower bud break in Japanese apricot are enhanced by exogenous GA4. Hortic. Res. 2015, 2, 15046. [Google Scholar] [CrossRef]
- Gillespie, L.M.; Volaire, F.A. Are winter and summer dormancy symmetrical seasonal adaptive strategies? The case of temperate herbaceous perennials. Ann. Bot. 2017, 119, 311–323. [Google Scholar] [CrossRef]
- Lloret, A.; Badenes, M.L.; Ríos, G. Modulation of dormancy and growth responses in reproductive buds of temperate trees. Front. Plant Sci. 2018, 9, 1368. [Google Scholar] [CrossRef]
- Martín-Fontecha, E.S.; Tarancon, C.; Cubas, P. To grow or not to grow, a power-saving program induced in dormant buds. Curr. Opin. Plant Biol. 2018, 41, 102–109. [Google Scholar] [CrossRef]
- Falavigna, V.S.; Guitton, B.; Costes, E.; Andrés, F.I. Want to (bud) break free: The potential role of DAM and SVP-like genes in regulating dormancy cycle in temperate fruit trees. Front. Plant Sci. 2019, 9, 1990. [Google Scholar] [CrossRef]
- Liu, J.; Sherif, S.M. Hormonal orchestration of bud dormancy cycle in deciduous woody perennials. Front. Plant Sci. 2019, 10, 1136. [Google Scholar] [CrossRef]
- Vimont, N.; Fouché, M.; Campoy, J.A.; Tong, M.; Arkoun, M.; Yvin, J.-C.; Wigge, P.A.; Dirlewanger, E.; Cortijo, S.; Wenden, B. From bud formation to flowering: Transcriptomic state defines the cherry developmental phases of sweet cherry bud dormancy. BMC Genom. 2019, 20, 974. [Google Scholar] [CrossRef]
- Buijs, G.A. Perspective on Secondary Seed Dormancy in Arabidopsis thaliana. Plants 2020, 9, 749. [Google Scholar] [CrossRef] [PubMed]
- Goeckeritz, C.; Hollender, C.A. There is more to flowering than those DAM genes: The biology behind bloom in rosaceous fruit trees. Curr. Opin. Plant Biol. 2021, 59, 101995. [Google Scholar] [CrossRef] [PubMed]
- Campoy, J.A.; Darbyshire, R.; Dirlewanger, E.; Quero-García, J.; Wenden, B. Yield potential definition of the chilling requirement reveals likely underestimation of the risk of climate change on winter chill accumulation. Int. J. Biometeorol. 2019, 63, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Dai, J.; Ranjitkar, S.; Yu, H.; Xu, J.; Luedeling, E. Chilling and heat requirements for flowering in temperate fruit trees. Int. J. Biometeorol. 2014, 58, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Campoy, J.A.; Ruiz, D.; Allderman, L.; Cook, N.; Egea, J. The fulfilment of chilling requirements and the adaptation of apricot (Prunus armeniaca L.) in warm winter climates: An approach in Murcia (Spain) and the Western Cape (South Africa). Eur. J. Agron. 2012, 37, 43–55. [Google Scholar] [CrossRef]
- El Yaacoubi, A.; Malagi, G.; Oukabli, A.; Citadin, I.; Hafidi, M.; Bonhomme, M.; Legave, J.-M. Differentiated dynamics of bud dormancy and growth in temperate fruit trees relating to bud phenology adaptation, the case of apple and almond trees. Int. J. Biometeorol. 2016, 60, 1695–1710. [Google Scholar] [CrossRef]
- Kaufmann, H.; Blanke, M. Performance of three numerical models to assess winter chill for fruit trees—A case study using cherry as model crop in Germany. Reg. Environ. Chang. 2017, 17, 715–723. [Google Scholar] [CrossRef]
- Nasrabadi, M.; Ramezanian, A.; Eshghi, S.; Sarkhosh, A. Chilling and heat requirement of pomegranate (Punica granatum L.) trees grown under sustained deficit irrigation. Sci. Hortic. 2020, 263, 109117. [Google Scholar] [CrossRef]
- Kühn, N.; Ormeño-Nuñez, J.; Jaque-Zamora, G.; Pérez, F.J. Photoperiod modifies the diurnal expression profile of VvPHYA and VvPHYB transcripts in field-grown grapevine leaves. J. Plant Physiol. 2009, 166, 1172–1180. [Google Scholar] [CrossRef]
- Javanshah, A.; Alipour, H.; Hadavi, F. A model for assessing the chill units received in Kerman and Rafsanjan areas. Acta Hortic. 2006, 726, 221–225. [Google Scholar] [CrossRef]
- Jangra, M.S.; Sharma, J.P. Climate resilient apple production in Kullu valley of Himachal Pradesh. Int. J. Farm Sci. 2013, 3, 91–98. [Google Scholar]
- Esper, J.; Cook, E.R.; Schweingruber, F.H. Low-frequency signals in long tree-ring chronologies for reconstructing past temperature variability. Science 2002, 295, 2250–2253. [Google Scholar] [CrossRef]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. IPCC (2013): Climate Change The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013; pp. 1–1535. [Google Scholar]
- Perez-Lopez, D.; Ribas, F.; Moriana, A.; Rapoport, H.F.; De Juan, A. Influence oftemperature on the growth and development of olive (Olea europaea L.) trees. J. Hortic. Sci. Biotechnol. 2008, 83, 171–176. [Google Scholar] [CrossRef]
- Orlandi, F.; Garcia-Mozo, H.; Galán, C.; Romano, B.; de la Guardia, C.D.; Ruiz, L.; del Mar Trigo, M.; Dominguez-Vilches, E.; Fornaciari, M. Olive flowering trends in a large Mediterranean area (Italy and Spain). Int. J. Biometeorol. 2010, 54, 151–163. [Google Scholar] [CrossRef]
- Ezzat, A.; Amriskó, L.G.; Mikita, G.; Nyéki, T.; Soltész, J.; Szabó, Z.M. Variation between some apricot varieties in regard to flowering phenology in Boldogkôváralja, Hungary. Int. J. Hortic. Sci. 2012, 18, 7–9. [Google Scholar] [CrossRef][Green Version]
- Jana, B.R.; Das, B. Effect of dormancy breaking chemicals on flowering, fruit set and quality in Asian pear (Pyrus pyrifolia L.). Afr. J. Agric. Res. 2014, 9, 56–60. [Google Scholar] [CrossRef]
- Yuri, J.A.; Moggia, C.; Sepulveda, A.; Poblete-Echeverría, C.; Valdés-Gómez, H.; Torres, C.A. Effect of cultivar, rootstock, and growing conditions on fruit maturity and postharvest quality as part of a six-year apple trial in Chile. Sci. Hortic. 2019, 253, 70–79. [Google Scholar] [CrossRef]
- Rana, R.S.; Bhagat, R.M.; Kalia, V.; Lal, H. Impact of climate change on shift of apple belt in Himachal Pradesh. In Proceedings of the Workshop on Impact of Climate Change on Agriculture, Ahmedabad, India, 17–18 December 2009; ISPRS Ar-chives 38-8/3. Panigrahy, S., Ray, S.S., Parihar, J.S., Eds.; Excel India Publishers: New Delhi, India, 2009; pp. 131–137. [Google Scholar]
- Stockle, C.O.; Nelson, R.L.; Higgins, S.; Brunner, J.; Grove, G. Assessment of climate change impact on Eastern Washington agriculture. Clim. Chang. 2010, 102, 77–102. [Google Scholar] [CrossRef]
- Byrne, D.H.; Sherman, W.B.; Bacon, T.A. Stone fruit genetic pool and its exploitation for growing under warm climatic conditions. In Temperate Fruit Crops in Warm Climates; Erez, A., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 157–230. [Google Scholar]
- Mohamed, A.K.A. The effect of chilling, defoliation and hydrogen cyanamide on dormancy release, bud break and fruiting of Anna apple cultivar. Sci. Hortic. 2008, 118, 25–32. [Google Scholar] [CrossRef]
- Fernandez, E.; Whitney, C.; Luedeling, E. The importance of chill model selection—A multi-site analysis. Eur. J. Agron. 2020, 119, 126103. [Google Scholar] [CrossRef]
- Weinberger, J. Chilling requirements of peach varieties. Proc. Am. Soc. Hortic. Sci. 1950, 56, 122–128. [Google Scholar]
- Richardson, E.; Seeley, S.D.; Walker, D. A model for estimating the completion of rest for Redhaven and Elberta peach trees. HortScience 1974, 9, 331–332. [Google Scholar]
- Cabré, F.; Nuñez, M. Impacts of climate change on viticulture in Argentina. Reg. Environ. Chang. 2020, 20, 12. [Google Scholar] [CrossRef]
- Benmoussa, H.; Luedeling, E.; Ghrab, M.; Ben Mimoun, M. Severe winter chill decline impacts Tunisian fruit and nut orchards. Clim. Chang. 2020, 162, 1249–1267. [Google Scholar] [CrossRef]
- Landsberg, J.J. Apple fruit bud development and growth; analysis and an empirical model. Ann. Bot. 1974, 38, 1013–1023. [Google Scholar] [CrossRef]
- Gilreath, P.R.; Buchanan, D.W. Rest prediction model for low-chilling Sungold nectarine. J. Am. Soc. Hortic. Sci. 1981, 106, 426–429. [Google Scholar]
- Shaltout, A.D.; Unrath, C.R. Rest completion prediction model for Starkrimson Delicious apples. J. Am. Soc. Hortic. Sci. 1983, 108, 957–961. [Google Scholar]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature-dependence of dormancy breaking in plants - mathematical-analysis of a 2-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Linsley-Noakes, G.; Allan, P.; Matthee, G. Modification of rest completion prediction models for improved accuracy in South African stone fruit orchards. J. S. Afr. Soc. Hort. Sci. 1994, 4, 13–15. [Google Scholar]
- Guak, S.; Neilsen, D. Chill Unit models for predicting dormancy completion of floral buds in apple and sweet cherry. Hort. Environ. Biotechnol. 2013, 54, 29–36. [Google Scholar] [CrossRef]
- Jones, H.G.; Hillis, R.M.; Gordon, S.L.; Brennan, R.M. An approach to the determination of winter chill requirements for different Ribes cultivars. Plant Biol. 2013, 15, 18–27. [Google Scholar] [CrossRef]
- Jones, H.G.; Gordon, S.L.; Brennan, R.M. Chilling requirement of Ribes cultivars. Front. Plant Sci. 2015, 5, 767. [Google Scholar] [CrossRef]
- Luedeling, E. chillR: Statistical Methods for Phenology Analysis in Temperate Fruit Trees. R Package Version 0.70.21. Available online: https://cran.r-project.org/web/packages/chillR/ (accessed on 18 April 2021).
- Parkes, H.; Darbyshire, R.; White, N. Chilling requirements of apple cultivars grown in mild Australian winter conditions. Sci. Hortic. 2020, 260, 108858. [Google Scholar] [CrossRef]
- Benmoussa, H.; Ghrab, M.; Ben Mimoun, M.; Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agric. For. Meteorol. 2017, 239, 34–46. [Google Scholar] [CrossRef]
- Egea, J.A.; Egea, J.; Ruiz, D. Reducing the uncertainty on chilling requirements for endodormancy breaking of temperate fruits by data-based parameter estimation of the dynamic model: A test case in apricot. Tree Physiol. 2020, 41, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Dai, J.; Ge, Q.; Wang, H.; Tao, Z. Comparison of chilling and heat requirements for leaf unfolding in deciduous woody species in temperate and subtropical China. Int. J. Biometeorol. 2020, 65, 393–403. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, T.; Frauenfeld, O.W.; Wang, K.; Sun, W.; Luo, J. Evaluation and quantification of surface air temperature over Eurasia based on CMIP5 models. Clim. Res. 2019, 77, 167–180. [Google Scholar] [CrossRef]
- Paltineanu, C.; Chitu, E. Climate change impact on phenological stages of sweet and sour cherry trees in a continental climate environment. Sci. Hortic. 2020, 261, 109011. [Google Scholar] [CrossRef]
- Boudichevskaia, A.; Kumar, G.; Sharma, Y.; Kapoor, R.; Singh, A.K. Challenges and strategies for developing climate-smart apple varieties through genomic approaches. In Genomic Designing of Climate-Smart Fruit Crops; Kole, C., Ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 23–71. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).