
The Application of Pomegranate, Sugar Apple, and Eggplant Peel Extracts Suppresses Aspergillus flavus Growth and Aflatoxin B1 Biosynthesis Pathway

 ,
,  ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungus Isolation and Identification
2.2. Peel Extracts Preparation
2.3. Total Phenolics Content
2.4. DPPH Radical Scavenging Ability
2.5. Gas Chromatography-Mass Spectroscopy Analysis
2.6. Effect of Fruit Peel Extracts on Fungal Biomass and Aflatoxin Production
2.6.1. Fungal Biomass Determination
2.6.2. Maize Storage Experiment
2.6.3. Aflatoxin B1 Extraction
2.6.4. Preparation of AFB1 Standard and HPLC Conditions
2.6.5. RNA Extraction, cDNA Synthesis, and qRT-PCR Assay
2.7. Statistical Analysis
3. Results and Discussion
3.1. Identification of the Aflatoxigenic Fungal Isolate
3.2. Fruit Peel Extracts Effect on A. flavus Biomass and AFB1 Production
3.3. Maize Storage Experiment
3.3.1. AFB1 Production
3.3.2. Grain Shape and Odor as Affected by Applied Fruit Peel Extract Treatments
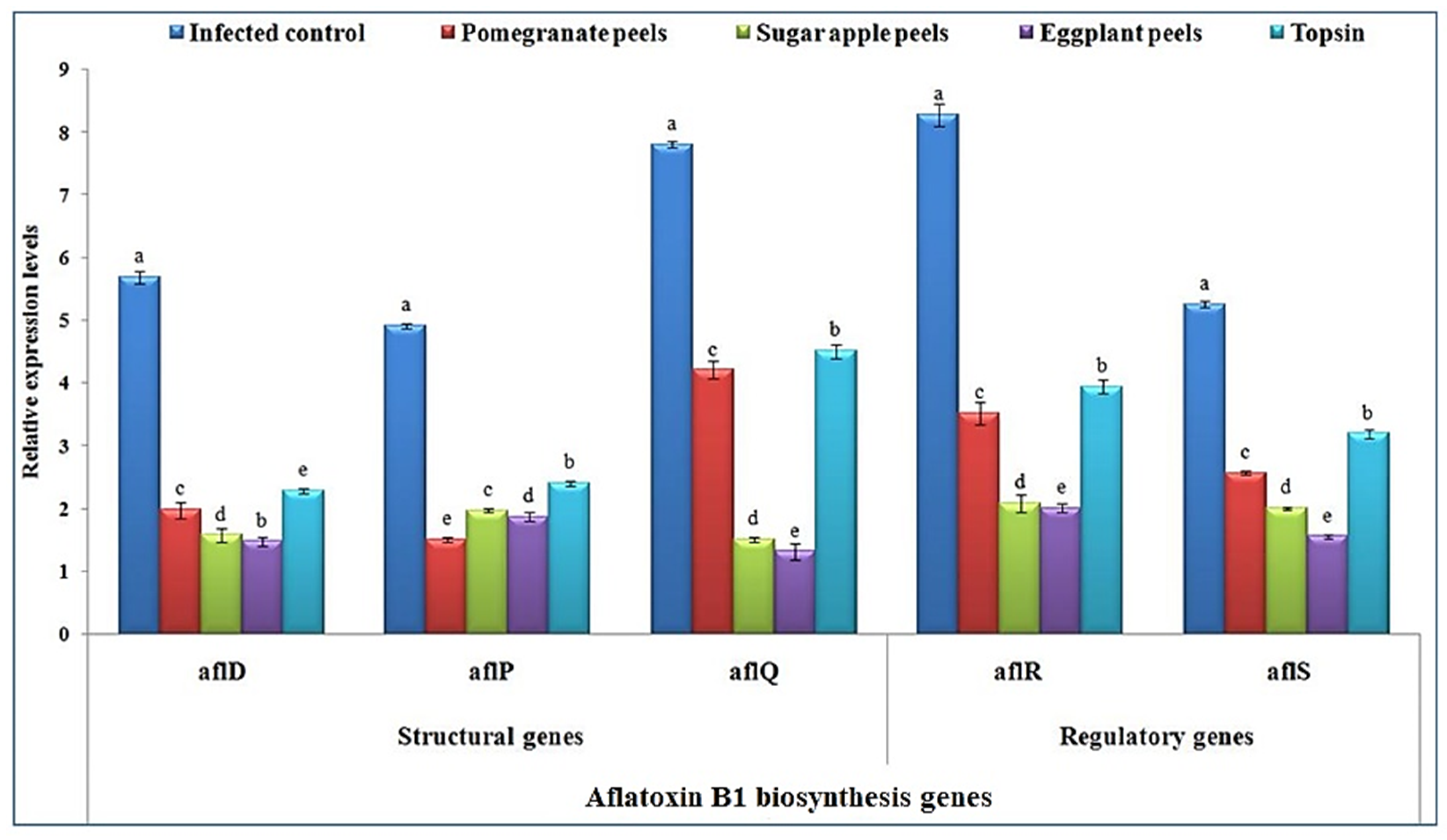
3.4. Transcriptional Levels of AFB1 Biosynthesis Genes
3.5. Total Phenolics Content of the Fruit Peel Extracts
3.6. DPPH Scavenging Ability
3.7. Bioactive Compounds Identified in Fruit Peel Extracts
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, F.; Bhatnagar, D.; Bui-Klimke, T.; Carbone, I.; Hellmich, R.; Munkvold, G.; Paul, P.; Payne, G.; Takle, E. Climate change impacts on mycotoxin risks in US maize. World Mycotoxin J. 2011, 4, 79–93. [Google Scholar] [CrossRef]
- Perrone, G.; Haidukowski, M.; Stea, G.; Epifani, F.; Bandyopadhyay, R.; Leslie, J.F.; Logrieco, A. Population structure and Aflatoxin production by Aspergillus Sect. Flavi from maize in Nigeria and Ghana. Food Microbiol. 2014, 41, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.N.; Bandyopadhyay, R.; Cotty, P.J. Degeneration of aflatoxin gene clusters in Aspergillus flavus from Africa and North America. AMB Express 2016, 6, 62. [Google Scholar] [CrossRef]
- Chauhan, N.M.; Washe, A.P.; Minota, T. Fungal infection and aflatoxin contamination in maize collected from Gedeo zone, Ethiopia. Springerplus 2016, 5, 753. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, M.N.; Omafe, O.M. Activity of three medicinal plants on fungi isolated from stored maize seeds (Zea mays (L.). Glob. J. Med. Plant Res. 2013, 1, 77–81. [Google Scholar]
- Galvano, F.; Ritieni, A.; Piva, G.; Pietri, A. Mycotoxins in the human food chain. Mycotoxin Blue Book 2005, 1, 187–224. [Google Scholar]
- Pietri, A.; Bertuzzi, T.; Pallaroni, L.; Piva, G. Occurrence of mycotoxins and ergosterol in maize harvested over 5 years in Northern Italy. Food Addit. Contam. 2004, 21, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Strosnider, H.; Azziz-Baumgartner, E.; Banziger, M.; Bhat, R.V.; Breiman, R.; Brune, M.-N.; DeCock, K.; Dilley, A.; Groopman, J.; Hell, K. Workgroup report: Public health strategies for reducing aflatoxin exposure in developing countries. Environ. Health Perspect. 2006, 114, 1898–1903. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Kareem, M.M.; Rasmey, A.M.; Zohri, A.A. The action mechanism and biocontrol potentiality of novel isolates of Saccharomyces cerevisiae against the aflatoxigenic Aspergillus flavus. Lett. Appl. Microbiol. 2019, 68, 104–111. [Google Scholar] [CrossRef]
- Naseer, R.; Sultana, B.; Khan, M.Z.; Naseer, D.; Nigam, P. Utilization of waste fruit-peels to inhibit aflatoxins synthesis by Aspergillus flavus: A biotreatment of rice for safer storage. Bioresour. Technol. 2014, 172, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Rahim, W.M.; Moawad, H.; Khalafallah, M. Enhancing the growth of promising fungal strains for rapid dye removal. Fresenius Environ. Bull. 2003, 12, 764–770. [Google Scholar]
- Lagogianni, C.; Tsitsigiannis, D. Effective chemical management for prevention of aflatoxins in maize. Phytopathol. Mediterr. 2018, 57, 186–197. [Google Scholar]
- Ashmawy, N.A.; Behiry, S.I.; Ali, H.M.; Salem, M.Z.M. Evaluation of Tecoma stans and Callistemon viminalis extracts against potato soft rot bacteria in vitro. J. Pure Appl. Microbiol. 2014, 8, 667–673. [Google Scholar]
- Abdelkhalek, A.; Salem, M.Z.M.; Ali, H.M.; Kordy, A.M.; Salem, A.Z.M.; Behiry, S.I. Antiviral, antifungal, and insecticidal activities of Eucalyptus bark extract: HPLC analysis of polyphenolic compounds. Microb. Pathog. 2020, 147, 104383. [Google Scholar] [CrossRef] [PubMed]
- Youssef, N.H.; Qari, S.H.; Matar, S.; Hamad, N.A.; Dessoky, E.S.; Elshaer, M.M.; Sobhy, S.; Abdelkhalek, A.; Zakaria, H.M.; Heflish, A.A. Licorice, Doum, and Banana Peel Extracts Inhibit Aspergillus flavus Growth and Suppress Metabolic Pathway of Aflatoxin B1 Production. Agronomy 2021, 11, 1587. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Alsubaie, M.M.; Behiry, S.I. First Report of Protective Activity of Paronychia argentea Extract against Tobacco Mosaic Virus Infection. Plants 2021, 10, 2435. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.G.P.; Porto, E.; Corrêa, C.B.; Alencar, S.M.; Gloria, E.M.; Cabral, I.S.R.; Aquino, L.M. Antimicrobial potential and chemical composition of agro-industrial wastes. J. Nat. Prod. 2012, 5, 27–36. [Google Scholar]
- Stabnikova, O.; Wang, J.-Y.; Ding, H.B. Biotransformation of vegetable and fruit processing wastes into yeast biomass enriched with selenium. Bioresour. Technol. 2005, 96, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.Y.; Lim, T.T.; Tee, J.J. Antioxidant properties of several tropical fruits: A comparative study. Food Chem. 2007, 103, 1003–1008. [Google Scholar] [CrossRef]
- Oliveira, L.; Freire, C.S.R.; Silvestre, A.J.D.; Cordeiro, N. Lipophilic extracts from banana fruit residues: A source of valuable phytosterols. J. Agric. Food Chem. 2008, 56, 9520–9524. [Google Scholar] [CrossRef]
- Adámez, J.D.; Samino, E.G.; Sánchez, E.V.; González-Gómez, D. In vitro estimation of the antibacterial activity and antioxidant capacity of aqueous extracts from grape-seeds (Vitis vinifera L.). Food Control 2012, 24, 136–141. [Google Scholar] [CrossRef]
- Katalinić, V.; Možina, S.S.; Skroza, D.; Generalić, I.; Abramovič, H.; Miloš, M.; Ljubenkov, I.; Piskernik, S.; Pezo, I.; Terpinc, P. Polyphenolic profile, antioxidant properties and antimicrobial activity of grape skin extracts of 14 Vitis vinifera varieties grown in Dalmatia (Croatia). Food Chem. 2010, 119, 715–723. [Google Scholar] [CrossRef]
- Hama, J.R.; Omer, R.A.; Rashid, R.S.M.; Mohammad, N.-E.-A.; Thoss, V. The Diversity of Phenolic Compounds along Defatted Kernel, Green Husk and Leaves of Walnut (Juglansregia L.). Anal. Chem. Lett. 2016, 6, 35–46. [Google Scholar] [CrossRef]
- Panichayupakaranant, P.; Tewtrakul, S.; Yuenyongsawad, S. Antibacterial, anti-inflammatory and anti-allergic activities of standardised pomegranate rind extract. Food Chem. 2010, 123, 400–403. [Google Scholar] [CrossRef]
- Sorrenti, V.; Randazzo, C.L.; Caggia, C.; Ballistreri, G.; Romeo, F.V.; Fabroni, S.; Timpanaro, N.; Raffaele, M.; Vanella, L. Beneficial effects of pomegranate peel extract and probiotics on pre-adipocyte differentiation. Front. Microbiol. 2019, 10, 660. [Google Scholar] [CrossRef]
- Glazer, I.; Masaphy, S.; Marciano, P.; Bar-Ilan, I.; Holland, D.; Kerem, Z.; Amir, R. Partial identification of antifungal compounds from Punica granatum peel extracts. J. Agric. Food Chem. 2012, 60, 4841–4848. [Google Scholar] [CrossRef] [PubMed]
- Pangallo, S.; Nicosia, M.G.L.D.; Agosteo, G.E.; Abdelfattah, A.; Romeo, F.V.; Cacciola, S.O.; Rapisarda, P.; Schena, L. Evaluation of a pomegranate peel extract as an alternative means to control olive anthracnose. Phytopathology 2017, 107, 1462–1467. [Google Scholar] [CrossRef]
- Rosas-Burgos, E.C.; Burgos-Hernández, A.; Noguera-Artiaga, L.; Kačániová, M.; Hernández-García, F.; Cárdenas-López, J.L.; Carbonell-Barrachina, Á.A. Antimicrobial activity of pomegranate peel extracts as affected by cultivar. J. Sci. Food Agric. 2017, 97, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Oliveira, M.; Furlong, E.B. Screening of antifungal and antimycotoxigenic activity of plant phenolic extracts. World Mycotoxin J. 2008, 1, 139–146. [Google Scholar] [CrossRef]
- Bouchra, C.; Mohamed, A.; Mina, I.H.; Hmamouchi, M. Antifungal activity of essential oils from several medicinal plants against four postharvest citrus pathogens. Phytopathol. Mediterr. 2003, 42, 251–256. [Google Scholar]
- Walters, D.; Raynor, L.; Mitchell, A.; Walker, R.; Walker, K. Antifungal activities of four fatty acids against plant pathogenic fungi. Mycopathologia 2004, 157, 87–90. [Google Scholar] [PubMed]
- Nakasugi, L.P.; Silva Bomfim, N.; Romoli, J.C.Z.; Botião Nerilo, S.; Veronezi Silva, M.; Rocha Oliveira, G.H.; Machinski, M., Jr. Antifungal and antiaflatoxigenic activities of thymol and carvacrol against Aspergillus flavus. Saúde Pesqui. 2021, 14, 113–123. [Google Scholar] [CrossRef]
- Tian, F.; Woo, S.Y.; Lee, S.Y.; Chun, H.S. p-Cymene and its derivatives exhibit antiaflatoxigenic activities against Aspergillus flavus through multiple modes of action. Appl. Biol. Chem. 2018, 61, 489–497. [Google Scholar] [CrossRef]
- Khan, R.; Ghazali, F.M.; Mahyudin, N.A.; Samsudin, N.I.P. Aflatoxin Biosynthesis, Genetic Regulation, Toxicity, and Control Strategies: A Review. J. Fungi 2021, 7, 606. [Google Scholar] [CrossRef] [PubMed]
- Caceres, I.; Al Khoury, A.; El Khoury, R.; Lorber, S.; Oswald, I.P.; El Khoury, A.; Atoui, A.; Puel, O.; Bailly, J.-D. Aflatoxin biosynthesis and genetic regulation: A review. Toxins 2020, 12, 150. [Google Scholar] [CrossRef]
- Ehrlich, K.C. Predicted roles of the uncharacterized clustered genes in aflatoxin biosynthesis. Toxins 2009, 1, 37–58. [Google Scholar] [CrossRef]
- Roze, L.V.; Hong, S.-Y.; Linz, J.E. Aflatoxin biosynthesis: Current frontiers. Annu. Rev. Food Sci. Technol. 2013, 4, 293–311. [Google Scholar] [CrossRef]
- Yu, J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef]
- Caceres, I.; El Khoury, R.; Medina, Á.; Lippi, Y.; Naylies, C.; Atoui, A.; El Khoury, A.; Oswald, I.P.; Bailly, J.-D.; Puel, O. Deciphering the anti-aflatoxinogenic properties of eugenol using a large-scale q-PCR approach. Toxins 2016, 8, 123. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, R.; Caceres, I.; Puel, O.; Bailly, S.; Atoui, A.; Oswald, I.P.; El Khoury, A.; Bailly, J.-D. Identification of the anti-aflatoxinogenic activity of Micromeria graeca and elucidation of its molecular mechanism in Aspergillus flavus. Toxins 2017, 9, 87. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Behiry, S.I.; Al-Askar, A.A. Bacillus velezensis PEA1 Inhibits Fusarium oxysporum Growth and Induces Systemic Resistance to Cucumber Mosaic Virus. Agronomy 2020, 10, 1312. [Google Scholar] [CrossRef]
- Salem, M.Z.M.; Behiry, S.I.; EL-Hefny, M. Inhibition of Fusarium culmorum, Penicillium chrysogenum and Rhizoctonia solani by n-hexane extracts of three plant species as a wood-treated oil fungicide. J. Appl. Microbiol. 2019, 126, 1683–1699. [Google Scholar] [CrossRef]
- Di Pinto, A.; Forte, V.; Guastadisegni, M.C.; Martino, C.; Schena, F.P.; Tantillo, G. A comparison of DNA extraction methods for food analysis. Food Control 2007, 18, 76–80. [Google Scholar] [CrossRef]
- Turkmen, N.; Sari, F.; Velioglu, Y.S. Effects of extraction solvents on concentration and antioxidant activity of black and black mate tea polyphenols determined by ferrous tartrate and Folin–Ciocalteu methods. Food Chem. 2006, 99, 835–841. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, D.-O.; Chun, O.K.; Shin, D.-H.; Jung, H.; Lee, C.Y.; Wilson, D.B. Phenolic extraction from apple peel by cellulases from Thermobifida fusca. J. Agric. Food Chem. 2005, 53, 9560–9565. [Google Scholar] [CrossRef] [PubMed]
- Asnaashari, M.; Farhoosh, R.; Sharif, A. Antioxidant activity of gallic acid and methyl gallate in triacylglycerols of Kilka fish oil and its oil-in-water emulsion. Food Chem. 2014, 159, 439–444. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Salem, M.Z.M.; Hafez, E.; Behiry, S.I.; Qari, S.H. The Phytochemical, Antifungal, and First Report of the Antiviral Properties of Egyptian Haplophyllum tuberculatum Extract. Biology 2020, 9, 248. [Google Scholar] [CrossRef] [PubMed]
- Youssef, N.H.; Qari, S.H.; Behiry, S.I.; Dessoky, E.S.; El-Hallous, E.I.; Elshaer, M.M.; Kordy, A.; Maresca, V.; Abdelkhalek, A.; Heflish, A.A. Antimycotoxigenic Activity of Beetroot Extracts against Altenaria alternata Mycotoxins on Potato Crop. Appl. Sci. 2021, 11, 4239. [Google Scholar] [CrossRef]
- Okla, M.K.; Alamri, S.A.; Salem, M.Z.M.; Ali, H.M.; Behiry, S.I.; Nasser, R.A.; Alaraidh, I.A.; Al-Ghtani, S.M.; Soufan, W. Yield, phytochemical constituents, and antibacterial activity of essential oils from the leaves/twigs, branches, branch wood, and branch bark of Sour Orange (Citrus aurantium L.). Processes 2019, 7, 363. [Google Scholar] [CrossRef]
- Velluti, A.; Sanchis, V.; Ramos, A.J.; Egido, J.; Marın, S. Inhibitory effect of cinnamon, clove, lemongrass, oregano and palmarose essential oils on growth and fumonisin B1 production by Fusarium proliferatum in maize grain. Int. J. Food Microbiol. 2003, 89, 145–154. [Google Scholar] [CrossRef]
- Alshannaq, A.F.; Gibbons, J.G.; Lee, M.-K.; Han, K.-H.; Hong, S.-B.; Yu, J.-H. Controlling aflatoxin contamination and propagation of Aspergillus flavus by a soy-fermenting Aspergillus oryzae strain. Sci. Rep. 2018, 8, 16871. [Google Scholar] [CrossRef]
- Hoeltz, M.; Welke, J.E.; Noll, I.B.; Dottori, H.A. Photometric procedure for quantitative analysis of aflatoxin B1 in peanuts by thin-layer chromatography using charge coupled device detector. Quim. Nova 2010, 33, 43–47. [Google Scholar] [CrossRef]
- Jangampalli Adi, P.; Matcha, B. Analysis of aflatoxin B1 in contaminated feed, media, and serum samples of Cyprinus carpio L. by high-performance liquid chromatography. Food Qual. Saf. 2018, 2, 199–204. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Sanan-Mishra, N. Differential expression profiles of tomato miRNAs induced by tobacco mosaic virus. J. Agric. Sci. Technol. 2019, 21, 475–485. [Google Scholar]
- Abdelkhalek, A.; Elmorsi, A.; Alshehaby, O.; Sanan-Mishra, N.; Hafez, E. Identification of genes differentially expressed in onion infected with Iris yellow spot virus. Phytopathol. Mediterr. 2018, 57, 334–340. [Google Scholar] [CrossRef]
- Hafez, E.E.; El-Morsi, A.A.; El-Shahaby, O.A.; Abdelkhalek, A.A. Occurrence of iris yellow spot virus from onion crops in Egypt. VirusDisease 2014, 25, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalek, A.; Al-Askar, A.A.; Hafez, E. Differential induction and suppression of the potato innate immune system in response to Alfalfa mosaic virus infection. Physiol. Mol. Plant Pathol. 2020, 110, 101485. [Google Scholar] [CrossRef]
- Abo-Zaid, G.A.; Matar, S.M.; Abdelkhalek, A. Induction of Plant Resistance against Tobacco Mosaic Virus Using the Biocontrol Agent Streptomyces cellulosae Isolate Actino 48. Agronomy 2020, 10, 1620. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Al-Huqail, A.A.; Behiry, S.I.; Salem, M.Z.M.; Ali, H.M.; Siddiqui, M.H.; Salem, A.Z.M. Antifungal, antibacterial, and antioxidant activities of Acacia saligna (Labill.) HL Wendl. flower extract: HPLC analysis of phenolic and flavonoid compounds. Molecules 2019, 24, 700. [Google Scholar] [CrossRef]
- Sadhasivam, S.; Shapiro, O.H.; Ziv, C.; Barda, O.; Zakin, V.; Sionov, E. Synergistic inhibition of mycotoxigenic fungi and mycotoxin production by combination of pomegranate peel extract and azole fungicide. Front. Microbiol. 2019, 10, 1919. [Google Scholar] [CrossRef] [PubMed]
- Basudan, N. Antioxidant, total phenolic content as well as antimicrobial potentiality effect of peel white and black eggplant extracts. Int. J. ChemTech Res. 2018, 11, 161–167. [Google Scholar] [CrossRef]
- Bernardo, J.S.; Sagum, R.S. Eggplant (Solanum melongena L.) peel as a potential functional ingredient in pan de sal. J. Nutr. Food Sci. 2016, 6, 67. [Google Scholar] [CrossRef]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemicals and antioxidant activity of milled fractions of different wheat varieties. J. Agric. Food Chem. 2005, 53, 2297–2306. [Google Scholar] [CrossRef] [PubMed]
- Laddomada, B.; Caretto, S.; Mita, G. Wheat bran phenolic acids: Bioavailability and stability in whole wheat-based foods. Molecules 2015, 20, 15666–15685. [Google Scholar] [CrossRef]
- Gemeda, N.; Woldeamanuel, Y.; Asrat, D.; Debella, A. Effect of essential oils on Aspergillus spore germination, growth and mycotoxin production: A potential source of botanical food preservative. Asian Pac. J. Trop. Biomed. 2014, 4, S373–S381. [Google Scholar] [CrossRef]
- El-Aziz, A.; Abeer, R.M.; Mahmoud, M.A.; Al-Othman, M.R.; Al-Gahtani, M.F. Use of selected essential oils to control aflatoxin contaminated stored cashew and detection of aflatoxin biosynthesis gene. Sci. World J. 2015, 2015, 958192. [Google Scholar] [CrossRef]
- Yazdani, D.; Ahmad, Z.A.M.; How, T.Y.; Jaganath, I.B.; Shahnazi, S. Inhibition of aflatoxin biosynthesis in Aspergillus flavus by phenolic compounds extracted of Piper betle L. Iran. J. Microbiol. 2013, 5, 428. [Google Scholar] [PubMed]
- Cleveland, T.E.; Yu, J.; Fedorova, N.; Bhatnagar, D.; Payne, G.A.; Nierman, W.C.; Bennett, J.W. Potential of Aspergillus flavus genomics for applications in biotechnology. Trends Biotechnol. 2009, 27, 151–157. [Google Scholar] [CrossRef]
- Lappa, I.K.; Dionysopoulou, A.M.; Paramithiotis, S.; Georgiadou, M.; Drosinos, E.H. Dual Transcriptional Profile of Aspergillus flavus during Co-Culture with Listeria monocytogenes and Aflatoxin B1 Production: A Pathogen–Pathogen Interaction. Pathogens 2019, 8, 198. [Google Scholar] [CrossRef]
- Yu, J.; Fedorova, N.D.; Montalbano, B.G.; Bhatnagar, D.; Cleveland, T.E.; Bennett, J.W.; Nierman, W.C. Tight control of mycotoxin biosynthesis gene expression in Aspergillus flavus by temperature as revealed by RNA-Seq. FEMS Microbiol. Lett. 2011, 322, 145–149. [Google Scholar] [CrossRef]
- Ullah, N.; Akhtar, K.P.; ul Hassan, S.W.; Asi, M.R.; Sadef, Y. First report of molecular characterization of Aspergillus flavus from maize in Pakistan. J. Plant Pathol. 2019, 101, 1289–1290. [Google Scholar] [CrossRef]
- Fakruddin, M.; Chowdhury, A.; Hossain, M.N.; Ahmed, M.M. Characterization of aflatoxin producing Aspergillus flavus from food and feed samples. Springerplus 2015, 4, 159. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Gogoi, R.; Srinivasa, N.; Shekhar, M. Characterization of aflatoxigenic Aspergillus flavus associated with aflatoxin B1 (AFB1) production in maize kernel in India. Indian Phytopathol. 2020, 74, 103–112. [Google Scholar] [CrossRef]
- Bhatnagar, D.; Ehrlich, K.C.; Cleveland, T.E. Molecular genetic analysis and regulation of aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2003, 61, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Papa, K.E. Norsolorinic acid mutant of Aspergillus flavus. Microbiology 1982, 128, 1345–1348. [Google Scholar] [CrossRef]
- Mayer, Z.; Färber, P.; Geisen, R. Monitoring the production of aflatoxin B1 in wheat by measuring the concentration of nor-1 mRNA. Appl. Environ. Microbiol. 2003, 69, 1154–1158. [Google Scholar] [CrossRef]
- Rodrigues, P.; Venâncio, A.; Kozakiewicz, Z.; Lima, N. A polyphasic approach to the identification of aflatoxigenic and non-aflatoxigenic strains of Aspergillus section Flavi isolated from Portuguese almonds. Int. J. Food Microbiol. 2009, 129, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Scherm, B.; Palomba, M.; Serra, D.; Marcello, A.; Migheli, Q. Detection of transcripts of the aflatoxin genes aflD, aflO, and aflP by reverse transcription–polymerase chain reaction allows differentiation of aflatoxin-producing and non-producing isolates of Aspergillus flavus and Aspergillus parasiticus. Int. J. Food Microbiol. 2005, 98, 201–210. [Google Scholar] [CrossRef]
- Sweeney, M.J.; Pàmies, P.; Dobson, A.D.W. The use of reverse transcription-polymerase chain reaction (RT-PCR) for monitoring aflatoxin production in Aspergillus parasiticus 439. Int. J. Food Microbiol. 2000, 56, 97–103. [Google Scholar] [CrossRef]
- Degola, F.; Berni, E.; Spotti, E.; Ferrero, I.; Restivo, F.M. Facing the problem of “false positives”: Re-assessment and improvement of a multiplex RT-PCR procedure for the diagnosis of A. flavus mycotoxin producers. Int. J. Food Microbiol. 2009, 129, 300–305. [Google Scholar] [CrossRef]
- Šavikin, K.; Živković, J.; Alimpić, A.; Zdunić, G.; Janković, T.; Duletić-Laušević, S.; Menković, N. Activity guided fractionation of pomegranate extract and its antioxidant, antidiabetic and antineurodegenerative properties. Ind. Crop. Prod. 2018, 113, 142–149. [Google Scholar] [CrossRef]
- Manochai, B.; Ingkasupart, P.; Lee, S.H.; Hong, J.H. Evaluation of antioxidant activities, total phenolic content (TPC), and total catechin content (TCC) of 10 sugar apple (Annona squamosa L.) cultivar peels grown in Thailand. Food Sci. Technol. 2018, 38, 294–300. [Google Scholar] [CrossRef]
- Ji, L.; Wu, J.; Gao, W.; Wei, J.; Yang, J.; Guo, C. Antioxidant capacity of different fractions of vegetables and correlation with the contents of ascorbic acid, phenolics, and flavonoids. J. Food Sci. 2011, 76, C1257–C1261. [Google Scholar] [CrossRef]
- Malviya, S.; Jha, A.; Hettiarachchy, N. Antioxidant and antibacterial potential of pomegranate peel extracts. J. Food Sci. Technol. 2014, 51, 4132–4137. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Blois, M. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Rao, L.J. Phenolic constituents from the lichen Parmotrema stuppeum (Nyl.) Hale and their antioxidant activity. Z. Naturforsch. C 2000, 55, 1018–1022. [Google Scholar] [CrossRef]
- Li, Y.; Guo, C.; Yang, J.; Wei, J.; Xu, J.; Cheng, S. Evaluation of antioxidant properties of pomegranate peel extract in comparison with pomegranate pulp extract. Food Chem. 2006, 96, 254–260. [Google Scholar] [CrossRef]
- Çam, M.; İçyer, N.C.; Erdoğan, F. Pomegranate peel phenolics: Microencapsulation, storage stability and potential ingredient for functional food development. LWT—Food Sci. Technol. 2014, 55, 117–123. [Google Scholar] [CrossRef]
- Calvo, H.; Marco, P.; Blanco, D.; Oria, R.; Venturini, M.E. Potential of a new strain of Bacillus amyloliquefaciens BUZ-14 as a biocontrol agent of postharvest fruit diseases. Food Microbiol. 2017, 63, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Qiao, H.; Huang, L.; Buchenauer, H.; Han, Q.; Kang, Z.; Gong, Y. Biological control of take-all in wheat by endophytic Bacillus subtilis E1R-j and potential mode of action. Biol. Control 2009, 49, 277–285. [Google Scholar] [CrossRef]
- Ali, I.; Khan, F.G.; Suri, K.A.; Gupta, B.D.; Satti, N.K.; Dutt, P.; Afrin, F.; Qazi, G.N.; Khan, I.A. In vitro antifungal activity of hydroxychavicol isolated from Piper betle L. Ann. Clin. Microbiol. Antimicrob. 2010, 9, 7. [Google Scholar] [CrossRef]
- Mathekga, A.D.M.; Meyer, J.J.M.; Horn, M.M.; Drewes, S.E. An acylated phloroglucinol with antimicrobial properties from Helichrysum caespititium. Phytochemistry 2000, 53, 93–96. [Google Scholar] [CrossRef]
- Hua, S.; Grosjean, O.; Baker, J.L. Inhibition of aflatoxin biosynthesis by phenolic compounds. Lett. Appl. Microbiol. 1999, 29, 289–291. [Google Scholar] [CrossRef]
- Lavault, M.; Landreau, A.; Larcher, G.; Bouchara, J.-P.; Pagniez, F.; Le Pape, P.; Richomme, P. Antileishmanial and antifungal activities of xanthanolides isolated from Xanthium macrocarpum. Fitoterapia 2005, 76, 363–366. [Google Scholar] [CrossRef]
- Inamori, Y.; Kobayashi, T.; Ogawa, M.; Tsujibo, H. Biological Actibities of Hexesterol. Chem. Pharm. Bull. 1988, 36, 815–818. [Google Scholar] [CrossRef]
- Stappen, I.; Tabanca, N.; Ali, A.; Wanner, J.; Lal, B.; Jaitak, V.; Wedge, D.E.; Kaul, V.K.; Schmidt, E.; Jirovetz, L. Antifungal and repellent activities of the essential oils from three aromatic herbs from western Himalaya. Open Chem. 2018, 16, 306–316. [Google Scholar] [CrossRef]
- Ragasa, C.Y.; Alimboyoguen, A.B.; Urban, S.; Raga, D.D. A bioactive diterpene from Smallanthus sonchifolius. Nat. Prod. Commun. 2008, 3. [Google Scholar] [CrossRef]
- Padla, E.P.; Solis, L.T.; Ragasa, C.Y. Antibacterial and antifungal properties of ent-kaurenoic acid from Smallanthus sonchifolius. Chin. J. Nat. Med. 2012, 10, 408–414. [Google Scholar] [CrossRef]
- Bisht, D.; Pal, A.; Chanotiya, C.S.; Mishra, D.; Pandey, K.N. Terpenoid composition and antifungal activity of three commercially important essential oils against Aspergillus flavus and Aspergillus niger. Nat. Prod. Res. 2011, 25, 1993–1998. [Google Scholar] [CrossRef] [PubMed]
- Zore, G.B.; Thakre, A.D.; Jadhav, S.; Karuppayil, S.M. Terpenoids inhibit Candida albicans growth by affecting membrane integrity and arrest of cell cycle. Phytomedicine 2011, 18, 1181–1190. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene | Primer Sequences (5′-3′) | Function in the Biosynthesis Pathway of AFB1 |
|---|---|---|
| β-tubulin | F: CTTGTTGACCAGGTTGTGGAT R: GTCGCAGCCCTCAGCCT | Reference gene |
| aflD | F: GTCCAAGCAACAGGCCAAGT R: TCGTGCATGTTGGTGATGGT | Norsolorinic acid (NOR) → Averantin (AVN) |
| aflP | F: GGCCGCCGCTTTGATCTAGG R: ACCACGACCGCCGCC | Sterigmatocystin (ST) → O-methylsterigmatocystin (OMST) |
| aflQ | F: GTGTCCGCAGTGTCTAGCTT R: GCTCAAAGGTCGCCAGAGTA | OMST → AFB1 |
| aflR | F: CTCAAGGTGCTGGCATGGTA R: CAGCTGCCACTGTTGGTTTC | Regulator gene |
| aflS | F: CTGCAGCTATATTGCCCACA R: TAAACCCAGGCAGAGTTGGT | Regulator gene |
| Solvent Concentration (%) | Control AFB1 Production (μg/L) | Control | Weight of A. flavus Mat (g) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pomegranate | Sugar Apple | Eggplant | |||||||||||||
| Wet | Dry | Wet | Dry | AFB1 (μg/L) | AI% | Wet | Dry | AFB1 (μg/L) | AI% | Wet | Dry | AFB1 (μg/L) | AI% | ||
| Ethanol | |||||||||||||||
| 25 | 23.58 | 6 | 0.97 | 4.56 | 0.62 | 2.19 | 90.70 | 1.16 | 0.08 | 8.52 | 63.86 | 6.16 | 0.88 | 2.88 | 87.79 |
| 50 | 24.74 | 6.97 | 1.06 | 4.59 | 0.44 | 3.18 | 87.13 | 1.19 | 0.17 | 5.32 | 78.49 | 4.66 | 0.63 | 6.79 | 72.55 |
| 75 | 25.20 | 3.59 | 0.62 | 0.09 | 0.03 | 3.69 | 85.35 | 0.16 | 0.05 | 4.48 | 82.23 | 5.60 | 0.75 | 2.52 | 90.60 |
| Acetone | |||||||||||||||
| 25 | 22.49 | 4.58 | 0.67 | 3.78 | 0.69 | 2.18 | 90.32 | 0.17 | 0.02 | 5.97 | 73.45 | 3.30 | 0.62 | 9.04 | 59.79 |
| 50 | 20.12 | 5.05 | 0.74 | 4.33 | 0.93 | 3.46 | 82.79 | 0.14 | 0.02 | 6.58 | 67.29 | 5.57 | 0.56 | 9.60 | 52.30 |
| 75 | 17.56 | 4.33 | 0.73 | 5.12 | 1.00 | 2.57 | 85.37 | 0.26 | 0.01 | 3.52 | 79.97 | 5.19 | 0.82 | 10.77 | 38.66 |
| Methanol | |||||||||||||||
| 25 | 17.00 | 7.49 | 0.71 | 4.34 | 0.72 | 6.42 | 62.26 | 0.12 | 0.01 | 10.50 | 38.23 | 5.16 | 0.57 | 8.09 | 52.41 |
| 50 | 13.85 | 7.16 | 0.91 | 4.84 | 0.74 | 3.44 | 75.15 | 0.32 | 0.13 | 8.93 | 35.52 | 7.19 | 0.65 | 8.69 | 35.97 |
| 75 | 14.08 | 5.27 | 0.62 | 0.17 | 0.04 | 2.60 | 81.50 | 5.83 | 0.75 | 8.85 | 37.12 | 1.89 | 0.24 | 4.16 | 70.43 |
| Diethyl ether | |||||||||||||||
| 25 | 26.44 | 5.09 | 0.9 | 5.83 | 0.79 | 2.42 | 90.87 | 0.24 | 0.07 | 13.71 | 48.15 | 5.84 | 0.80 | 3.54 | 86.62 |
| 50 | 24.83 | 6.08 | 0.63 | 4.42 | 0.85 | 5.23 | 78.96 | 0.19 | 0.01 | 5.07 | 79.59 | 6.46 | 0.76 | 2.19 | 91.18 |
| 75 | 24.16 | 7.05 | 0.73 | 4.18 | 0.86 | 6.86 | 71.61 | 2.21 | 0.39 | 5.92 | 75.50 | 6.31 | 0.78 | 2.19 | 90.95 |
| L.S.D.0.05 | 1.47 | 0.08 | 0.47 | 0.03 | 0.16 | 0.02 | 0.03 | 0.01 | 0.49 | 0.03 | 0.12 | ||||
| Treatments | AFB1 (μg/L) | AI% |
|---|---|---|
| Healthy control | 0.00 | 100 |
| Infected control | 532 | 0.00 |
| 25%-Diethyl ether pomegranate peel extract | 112.64 | 78.83 |
| 75%-Ethanol sugar apple peel extract | 27.39 | 94.85 |
| 50%-Diethyl ether eggplant peel extract | 20.72 | 96.11 |
| Topsin fungicide (2.5 mg/mL) | 143.92 | 72.95 |
| Treatments | Scale | Odor | Shape |
|---|---|---|---|
| Healthy control | 5 | 0 | 5 |
| Infected control | 0 | 5 | 0 |
| 25% Diethyl ether pomegranate peel extract | 4 | 2 | 4 |
| 75% Ethanol sugar apple peel extract | 5 | 1 | 5 |
| 50% Diethyl ether eggplant peel extract | 5 | 1 | 5 |
| Topsin fungicide (2.5 mg/mL) | 0 | 5 | 0 |
| Fruit Peel Extract | TPC (mgGAEs/g DW) ± SD | TAA (μg/mL) ± SD |
|---|---|---|
| 25% Diethyl ether pomegranate | 116.88 ± 1.44 | 86.76 ± 0.22 |
| 75% Ethanol sugar apple | 35.09 ± 1.79 | 94.02 ± 0.08 |
| 50% Diethyl ether eggplant | 30.26 ± 1.76 | 52.94 ± 0.15 |
| Retention Time | Compound | Class | Peak Area (%) | ||
|---|---|---|---|---|---|
| Pomegranate | Sugar Apple | Eggplant | |||
| 3.70 | α-Fenchene | Bicyclic monoterpene | - | 11.03 | 4.67 |
| 4.90 | p-Allylphenol | Phenylpropene | 20.78 | 2.11 | 8.50 |
| 5.10 | Valproic acid | Saturated fatty acids | - | 5.04 | 0.79 |
| 5.31 | Ethylnorbornene | Cyclic hydrocarbons | 4.93 | 1.14 | 1.68 |
| 5.60 | Phenylglyoxylic acid | Carboxylic acids | 2.26 | 1.43 | 0.62 |
| 6.20 | 4-Methylcatechol | Polyphenols | 0.76 | 4.74 | 0.93 |
| 6.34 | Sorbitol | Sugar alcohols | 5.66 | - | - |
| 6.60 | 7,8-Dihydro-α-ionone | Carotenoids | - | 2.79 | 0.51 |
| 6.63 | 4-Ethylbezaldehyde | Aldehydes | 3.07 | - | - |
| 6.88 | Dimethoxy durene | Alkylbenzene | 0.57 | 0.77 | 3.76 |
| 7.05 | Dimethyl caffeic acid | Phenolic acids | 1.66 | - | - |
| 7.15 | Scopoletin | Phenylpropanoids | 0.62 | - | - |
| 7.45 | Farnesol | Sesquiterpenes | 0.29 | 1.87 | 0.46 |
| 7.70 | Hexestrol | Nonsteroidal estrogen | 0.35 | 5.87 | 0.29 |
| 7.91 | p-Cymene | Monoterpenes | 1.00 | - | 0.37 |
| 8.04 | α-Terpineol | Monoterpenes | - | - | 0.59 |
| 8.11 | Stevioside | Diterpene glycosides | 1.29 | 0.43 | 0.43 |
| 8.27 | γ-Terpinene | Monoterpenes | - | 3.24 | 0.32 |
| 8.55 | Levoglucosenone | Heterocyclic ketones | 4.76 | - | - |
| 8.59 | 6-Hydroxyflavone | Flavonoids | 0.78 | 0.39 | 0.35 |
| 8.94 | Resveratrol | Polyphenols | - | 0.69 | 0.61 |
| 9.49 | N-Acetylneuraminic acid | Alpha-keto acid sugars | - | 3.01 | - |
| 9.76 | D-mannose | Carbohydrates | 4.70 | - | - |
| 9.90 | Xanthinin | Sesquiterpene lactones | 7.05 | 3.80 | 0.53 |
| 10.10 | 3,5-Dihydroxyphenol | Phenoles | 9.37 | - | - |
| 10.63 | 3,5,7-Tri-O-methylgalangin | Flavonoids | - | 0.50 | - |
| 11.00 | p-Menthone | Monoterpenes | 0.96 | - | - |
| 11.20 | δ-Elemene | Sesquiterpenes | - | - | 2.14 |
| 11.47 | β-lonol | Sesquiterpenes | 1.35 | - | - |
| 11.67 | α-Selinene | Sesquiterpenes | - | - | 0.70 |
| 12.00 | Caryophyllene | Sesquiterpenes | - | - | 0.69 |
| 12.26 | Kaempferol | Flavonoids | - | - | 0.26 |
| 12.34 | 5,7,3′,4′-Tetrahydroxflavanone | Flavonoids | - | - | 0.54 |
| 12.47 | β-Patchoulene | Polycyclic hydrocarbons | - | - | 1.09 |
| 12.55 | β-Gurjunene | Sesquiterpenes | - | - | 0.67 |
| 12.76 | γ-Muurolene | Sesquiterpenes | - | - | 0.74 |
| 13.01 | Quercetin 7, 3, 4—Trimethoxy | Flavonoids | 1.50 | - | - |
| 13.20 | α-Himachalene | Sesquiterpenes | 4.33 | - | - |
| 13.70 | Longipinene | Epoxides | - | 5.83 | 0.72 |
| 14.95 | Apigenin 8-C-glucoside | Flavonoids | - | 0.65 | - |
| 15.10 | 4-Hydroxy-2-methoxybenzaldehyde | Methoxyphenols | - | 0.19 | - |
| 15.42 | Methyl 17-methyloctadecanoate | Fatty acid methyl esters | 0.67 | 0.23 | - |
| 15.70 | Octadecanoic acid | Fatty acids | 3.65 | 10.34 | 3.98 |
| 15.85 | Glycitein | Isoflavones | 0.74 | - | - |
| 15.85 | Stearic acid | Fatty acids | - | 2.17 | 0.91 |
| 16.19 | Phyllocladene | Diterpenoids | - | - | 4.29 |
| 16.52 | Luteolin 6,8-C-diglucoside | Flavonoids | 2.03 | - | 1.42 |
| 16.91 | (S)-(-)-Citronellic acid | Monoterpenes | - | 4.75 | - |
| 16.91 | Linoleic acid | Polyunsaturated fatty acids | 7.35 | - | 5.22 |
| 17.04 | 5β, 7Βh, 10α-Eudesm-11-en-1α-ol | Sesquiterpenes | 0.78 | - | |
| 17.05 | Quinine | Alkaloids | - | - | 0.28 |
| 17.10 | 1,16-Hexadecanedioic acid | Fatty acids | - | 4.20 | 1.40 |
| 17.79 | Tetrahydroisovelleral | Sesquiterpene dialdehydes | - | - | 2.29 |
| 18.26 | 2β-hydroxy-9-oxoverrucosane | Terepnoides | - | - | 1.04 |
| 18.60 | (+)-Beyerene | Diterpenes | - | 1.92 | 3.03 |
| 18.70 | Abietic acid | Diterpenes | - | - | 1.84 |
| 19.22 | α-Kaurene | Diterpenes | - | 6.33 | 25.67 |
| 19.89 | Methyl isopimarate | Diterpenes | - | 5.67 | 6.01 |
| 21.31 | Isosteviol | Diterpenes | - | 0.36 | 2.54 |
| 21.62 | 7α-Hydroxymanool | Diterpenes | - | 0.33 | 1.15 |
| 23.08 | Sclareol | Diterpene alcohols | - | 1.56 | 1.08 |
| 23.65 | Zeaxanthin | Carotenoids | 2.09 | - | - |
| 23.71 | Rhodopin | Carotenoids | - | 5.61 | 4.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismail, I.A.; Qari, S.H.; Shawer, R.; Elshaer, M.M.; Dessoky, E.S.; Youssef, N.H.; Hamad, N.A.; Abdelkhalek, A.; Elsamra, I.A.; Behiry, S.I. The Application of Pomegranate, Sugar Apple, and Eggplant Peel Extracts Suppresses Aspergillus flavus Growth and Aflatoxin B1 Biosynthesis Pathway. Horticulturae 2021, 7, 558. https://doi.org/10.3390/horticulturae7120558
Ismail IA, Qari SH, Shawer R, Elshaer MM, Dessoky ES, Youssef NH, Hamad NA, Abdelkhalek A, Elsamra IA, Behiry SI. The Application of Pomegranate, Sugar Apple, and Eggplant Peel Extracts Suppresses Aspergillus flavus Growth and Aflatoxin B1 Biosynthesis Pathway. Horticulturae. 2021; 7(12):558. https://doi.org/10.3390/horticulturae7120558
Chicago/Turabian StyleIsmail, Ismail A., Sameer H. Qari, Rady Shawer, Moustafa M. Elshaer, Eldessoky S. Dessoky, Nesrine H. Youssef, Najwa A. Hamad, Ahmed Abdelkhalek, Ibrahim A. Elsamra, and Said I. Behiry. 2021. "The Application of Pomegranate, Sugar Apple, and Eggplant Peel Extracts Suppresses Aspergillus flavus Growth and Aflatoxin B1 Biosynthesis Pathway" Horticulturae 7, no. 12: 558. https://doi.org/10.3390/horticulturae7120558
APA StyleIsmail, I. A., Qari, S. H., Shawer, R., Elshaer, M. M., Dessoky, E. S., Youssef, N. H., Hamad, N. A., Abdelkhalek, A., Elsamra, I. A., & Behiry, S. I. (2021). The Application of Pomegranate, Sugar Apple, and Eggplant Peel Extracts Suppresses Aspergillus flavus Growth and Aflatoxin B1 Biosynthesis Pathway. Horticulturae, 7(12), 558. https://doi.org/10.3390/horticulturae7120558