Abstract
The effect of thermal condition on the uptake of autumn and winter applied N and its subsequent utilization in spring tea (Camellia sinensis) was investigated by applying 15N enriched urea as single or split applications between October and February in two commercial plantations at Xingyang of Henan province and Yongchuan of Chongqing with different thermal conditions. The proportion of N derived from 15N-labeled urea (Ndff%) in fibrous root and mature leaves 15 days after application at Xingyang and the Ndff% of mature leaves on the day of the first spring tea harvest at both sites were the highest in the single October application. The Ndff% of the following spring tea was also the highest in the single October application at both sites. The results showed that application of N fertilizer in October relative to other later months most significantly improves the accumulation of plant N reserves and consequently contributes more significantly to the early spring tea. Such timing effect was related to the thermal condition, i.e., the growing degree days (°C•d, T > 8 °C) between the dates of fertilization and harvest of young shoots, which represents the combining effect of the temperature and the residence time of N fertilizer in the soil.
1. Introduction
Tea is one of the most popular beverages in the world, possessing many health benefits [1]. Between October and February, tea plants are usually in dormant state without active shoot growth because of the inappropriate low temperature. Young shoots of evergreen woody tea plants are harvested during March through the end of September to produce spring (March and April), summer (May, June, and July), and autumn (August and September) teas in subtropical areas of China. The quality of green tea is determined by the season of production and harvest standards. Spring tea has abundant free amino acids, reasonable polyphenol or catechin contents, and, consequently, a low but desirable ratio of polyphenol to total amino acids [2,3]. The high quality (so-called premium) green teas commanding good prices are mostly tip-harvested at the developmental stage of bud with one expanding young leaf in early spring (March and April) [4]. The average price of premium green tea is approximately three times that of common tea produced from later spring and other seasons. Today, the production of premium green teas, which account for over 70% of total tea value with approximately 49% of tea quantity, has become the prevailing pattern for much better economic benefits and marketing prospects [5]. Our previous work showed that the ability of nitrogen (N) storage and remobilization to young shoots is of particular importance to ensure good quality of spring tea [4].
The internal cycling of N reserves contributes a major proportion of N (up to 100%) to support the spring growth of trees, especially when the mineral N availability is low because of the slow mineralization rate under low temperature [6]. In deciduous trees, N reserves are built up by the re-translocation of nutrients in senescing leaves back to branches and roots and/or by absorption in autumn [7,8,9]. In evergreen trees, the N reserves are contributed from the previous absorption during active vegetative growth or the absorption from the soil during vegetative dormancy [10,11,12,13]. The size of N reserves is a determinant factor influencing the amounts subsequently remobilized and its capability to support spring growth [11,13]. Increasing N reserves improves the agronomic performance (e.g., canopy development, flowering, and fruit quality) of horticultural crops [14,15,16,17,18]. External N availability and fertilizer application timing greatly affect the size of N reserves. N supplied in the early phenological stage is absorbed at a high rate and mainly allocated to vegetative and/or fruit growth [19,20,21]. By contrast, postponing N supply to later stages is preferentially allocated to storage organs; therefore, this effectively increases the size of winter reserves, although the total N uptake is often reduced due to decreasing growth demand [22,23,24,25]. However, some works in field-grown peach and orange trees have reported controversial findings that adequate storage does not benefit from fertilization and soil N uptake in the late phenological stages [26,27].
N uptake and distribution in tea plants are determined by the demand imposed from shoot growth and phenological stage [28,29,30]. Tea yield and quality significantly respond to N nutrition, requiring frequent N fertilization to sustain yield with desirable quality [29,31,32,33]. Multiple N applications are frequently made in attempts to synchronize the supply and seasonal nutrient demand in accordance with local weather conditions. Fertilizers are applied as base fertilization between October and December and/or top-dressed sometime before the harvests of spring, summer, and autumn teas. Currently, in China, the production of premium green teas of very young tips harvested only in early spring (March until early April) has been the prevailing pattern. Our previous field experiment described a substantial absorption of N fertilizer applied in winter (January and February) by dormant tea plants [30] and contributed to the buildup of plant N reserve which is remobilized and makes a crucial contribution to the quality of spring tea [4,34]. The strategy of proper use of internal N reserves, which has been advocated in the optimization of N nutrition management in fruit trees, should also be considered in tea plantations to increase N use efficiency and avoid environmental contamination [35].
The timing of N fertilization must be finely tuned to focus on spring tea production, of which the N demand dynamics are largely different from summer and autumn teas. Few field experiments have been conducted to evaluate the timing effect of N applied in dormant stage on its absorption by tea plants, limited to just a few pot experiments [28,29,30]. In the present field experiments, 15N enriched urea was applied on different dates from late October through February at two tea plantations with different heat conditions. The enriched 15N tracing method has been successfully and widely applied to investigate N absorption, storage, remobilization, and cycling by horticultural plants [24,30,36]. We also compared the effect of a single application in late October or split applications in late October and January with identical N rates. The aim of the presented study was to investigate the timing effect of N fertilization on N adsorption and storage of tea plants in vegetative dormancy and subsequent utilization in the following spring tea.
2. Materials and Methods
Two tea plantations were selected for the field experiments. One was located at Yongchuan, Chongqing province (longitude 105°53′, latitude 29°23′, altitude 570 m) belonging to Chongqing Academy of Agricultural Sciences. Another tea plantation was located at Xingyang, Henan province (longitude 114°31′, latitude 32°16′, altitude 57 m) belonging to Xingyang Academy of Agricultural Sciences. The two plantations were established from rooted-cuttings of variety Fudingdaibai and Pingyangtezao, respectively. Both varieties are officially certified clones suitable for green tea. At both sites, bushes, each consisting of two rooted-cuttings, were cultivated in lines with a distance of 1.2 m. The distance between bushes of the same lines was 0.33 m. Weather data were collected from the local meteorological observatory. The mean annual air temperature and rainfall of the previous 20 years were 17.7 °C and 1015 mm at Yongchuan and 15.5 °C and 1106 mm at Xingyang, respectively. The two plantations were commercially managed before the present experiments to produce nationally known premium green teas, Yongchuan- Xiuya and Xingyang-Maojian, respectively. Prior to the experiment (on October 10), one composite soil sample was taken by auger from about 15 points from each experimental field, thoroughly mixed, and air dried for the analysis of soil chemical properties. Soil pH was measured in a 1:2.5 soil–water solution using a pH meter (Orion 3 Star, Thermo Ltd., Waltham, MA, USA). Contents of soil organic carbon and total nitrogen were measured using a C/N elemental analyzer (Vario Max, Elementar, Langenselbold, Germany). Soil available P, K, and Mg were extracted using the Mehlich 3 method, and then measured using Inductive Coupled Plasma (Thermo Jarrell Ash Ltd., Franklin, MA, USA). The soil properties are presented in Table 1. A total of 48 bushes with visually uniform canopy sizes were selected at each site. Plastic boards (0.4 cm thick) were buried vertically to 30 cm depth to isolate a mini-plot of 0.4 m2 (0.66 m in length and 1.2 m in width) containing two bushes [30]. Each plot was separated by one blank bush.

Table 1.
Selected soil properties of the experimental tea plantations in Yongchuan, Chongqing, and Xingyang, Henan province.
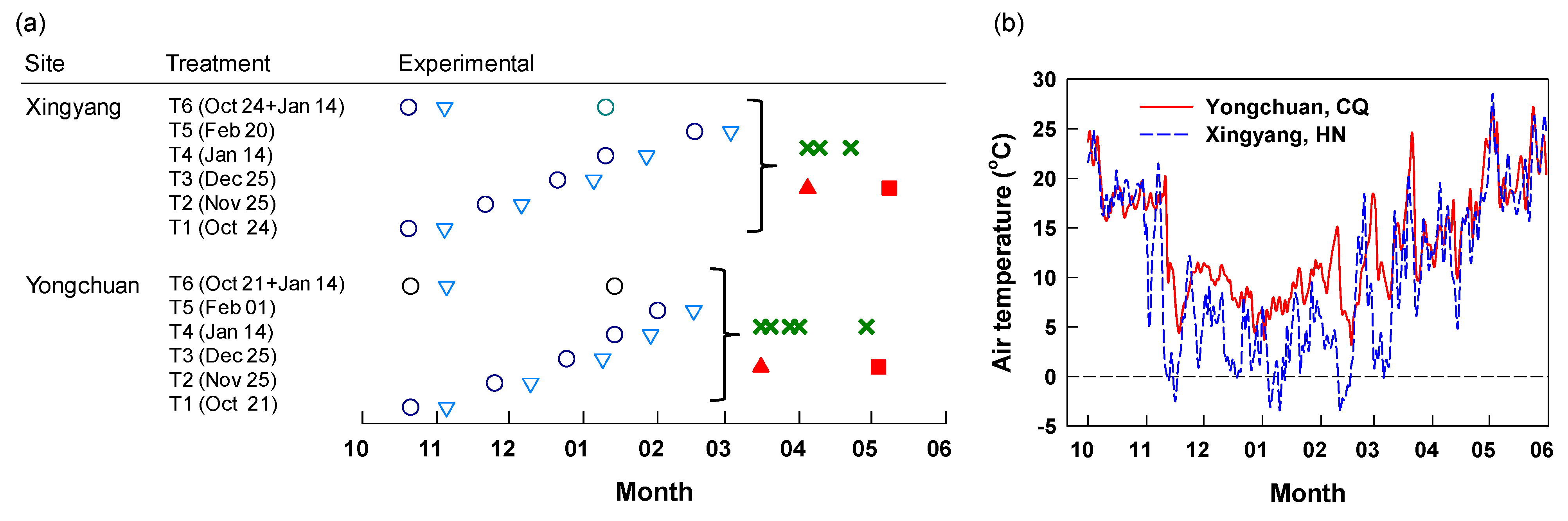
15N enriched urea (5.25% abundance, purchased from Shanghai Research Institute of Chemical Industry Co., Ltd., Shanghai, China) was applied to the soil of isolated mini-plot at a rate equivalent to 180 kg N ha−1 on five different dates or split into two applications. At Yongchuan, the dates of single N fertilization were October 21, November 25, and December 25, and January 14 and February 1 of the subsequent year. Split applications were implemented on October 21 and January 14 (Figure 1a). At Xingyang, individual N application dates were October 24, November 25, and December 25, and January 14 and February 20 of the subsequent year, or split applications on October 24 and January 14 (Figure 1a). The application in February at Xingyang was intentionally postponed because of the later harvest of spring tea. Other nutrients were applied at identical rates to all plots in case of low availability indicated by soil analysis. There were six treatments (five different dates and split applications) with four replications at each site. At Yongchuan, potash of sulfate (K2O, 150 kg ha−1) and kieserite (MgO, 30 kg ha−1) were applied on October 21 to compensate for the low availability of K and Mg of the soil. At Xingyang, single superphosphate (P2O5, 100 kg ha−1) and potash of sulfate (K2O, 150 kg ha−1) were applied on October 24 to compensate for its low availability of P and K.
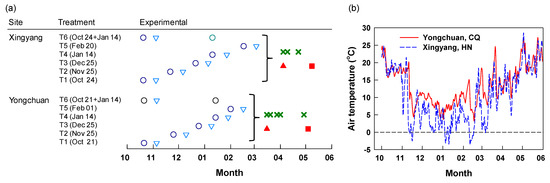
Figure 1.
The experimental design (a) and daily air temperature (b) at Yongchuan and Xingyang. Fertilization (○); sampling of fibrous roots and mature leaves 15 days after application (DAA15, ∇), on days of the first harvest (DFH, ▲) and post-harvest (DPH, ■); harvest of the following spring tea (Χ).
Samples of fibrous roots and mature leaves were taken from the mini-plots in three batches (Figure 1a). The first batch of samples was taken 15 days after each 15N application (DAA15). Soil cores were taken using an auger (4 cm diameter) from both sides within the application area at a depth of 0–20 cm. Fibrous roots < 2 mm in diameter were carefully sifted through a 2-mm mesh sieve, picked out, and rinsed thoroughly with deionized water, then blotted dry. Mature leaves were taken from the canopy surface at the same time as fibrous roots. The second batch of samples was taken on the day of the first harvest of young spring shoots (referred to as DFH) (March 16 at Yongchuan and April 8 at Xingyang) (Figure 1a). The third batch of samples was taken after the termination of spring teas (the day post-harvest, referred to as DPH) (May 4 at Yongchuan and May 12 at Xingyang) (Figure 1a). A few days were allowed to the sampling after the last harvest of young shoots to secure the termination of spring tea. Root and leaf samples were dried in an electric oven at 65 °C for constant weight.
Young spring shoots consisting of a bud with one young expanding leaf were harvested by hand, according to the standard for commercial green teas (Yongchuan-Xiuya and Xingyang-Maojian) (Figure 1a). At Yongchuan, the harvest of spring tea started on March 16 and finished on April 29. At Xingyang, the harvest of spring tea stared on April 8 and finished on April 26. All harvested young shoots were immediately treated in a microwave oven for 2 min to inactivate the polyphenol oxidase enzyme and finally dried in an electric oven at 65 °C for constant weight. Plant samples were ground to homogeneity with a ball mill (Mixer Mill MM300, Retsch, Haan, Germany) before measurements. Total N concentrations and the abundance of 15N were determined using an elemental analyzer (Thermo NE 1112) coupled (ConFlo III) to an isotope ratio mass spectrometer (∆PlusAD, Thermo Finnigen, Germany). To reduce analysis costs, samples of March 20 and 28 from the same plot at Yongchuan were pooled and measured. The uptake and utilization of 15N fertilizer were indicated by Ndff%, the proportion of N derived from 15N-labeled urea, which was calculated according to the following Equation (1).
The growing degree days of heat accumulation (GDD) (the sum of air temperature units above a threshold, T0) between the date of fertilization and the day of the first harvest (n) was calculated according to the following Equation (2) [37].
ANOVA was performed on datasets using the analysis module embedded in SigmaPlot 12.5 software (Systat Software, Inc.). Means of parameters from different fertilization dates at the same sampling times and those from different sampling times of the same fertilization dates were further distinguished by the Fisher LSD test in case of initial significance (p < 0.05). Statistical analysis was performed separately for each site.
3. Results
3.1. Thermal Condition at Two Sites
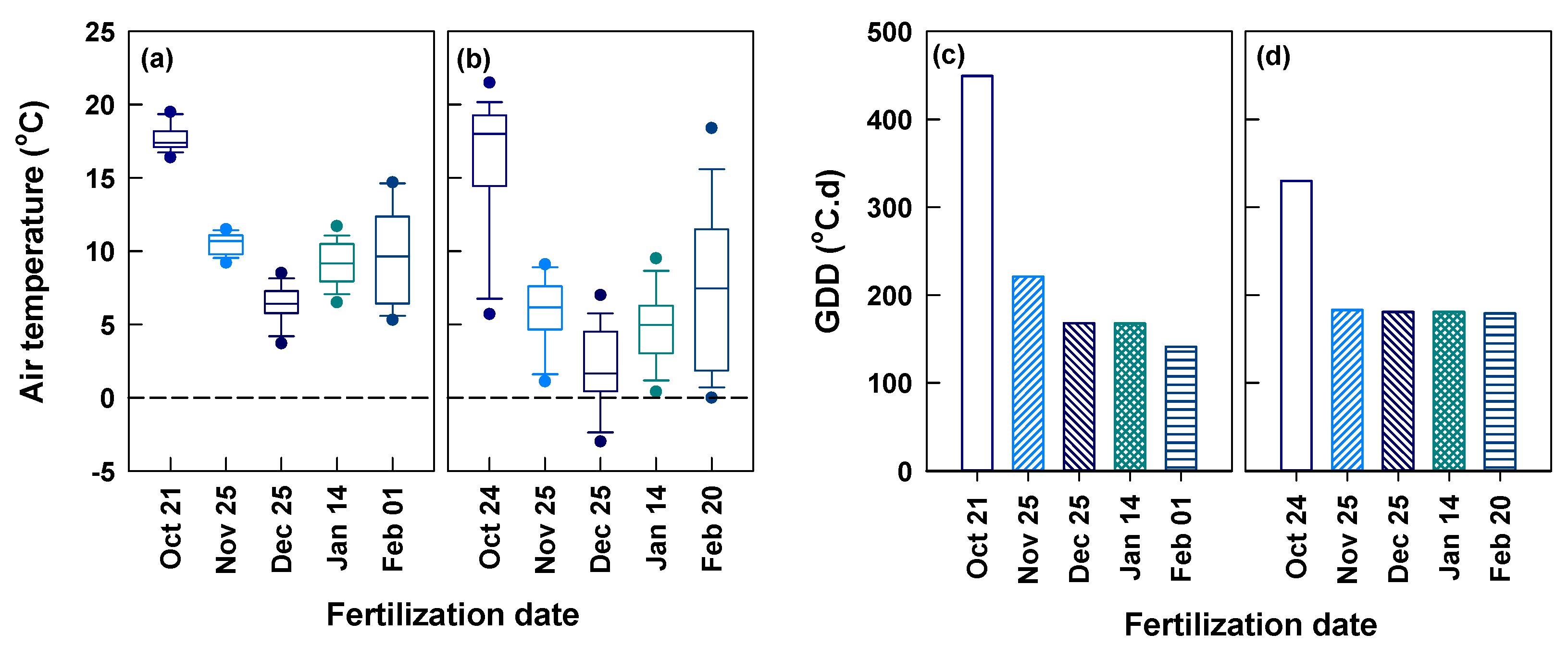
The daily air temperature within the experimental period varied between 3.2–27.2 °C and −3.5–28.2 °C from October 1 through May 30 at Yongchuan and Xingyang, respectively (Figure 1b). The air temperature was always above zero at Yongchuan. At Xingyang, it occasionally went down to below zero in November, January, and February. The two sites had the same magnitudes of air temperature in October and in the time beyond late March. In other dates, Yongchuan had higher air temperature than Xingyang. Periodic mean air temperature within 15 days after fertilization (DAA15) was the highest and lowest for applications in October and December at both sites, respectively (Figure 2a,b). The two sites had a similar mean temperature for the application in late October, except for low air temperature in a few days at Xingyang. In other applications, the periodic mean air temperature was lower at Xingyang than Yongchuan. We calculated the growing degree days (GDD, T0 > 8 °C) between the day of fertilization and the first harvest of spring tea (Figure 2c,d). The GDD at Yongchuan was the highest (449 °C•d) for the application in October and decreased in later applications between November and February. At Xingyang, GDD was 330 °C•d for the applications of October and decreased to a constant for later applications. Both sites had relatively low rainfall between October and February and rain increased in March (Figure S1). Yongchuan had higher cumulative rainfall between October and December but Xingyang turned to be higher after December.
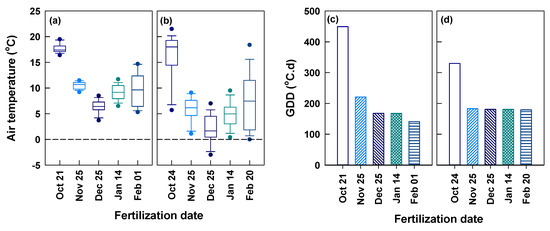
Figure 2.
Periodic mean air temperature within 15 days after fertilization (a,b) and growing degree days (GDD, ≥8 °C) between the fertilization dates and the first harvest of spring tea (c,d) at Yongchuan (a,c) and Xingyang (b,d).
3.2. 15N in Fibrous Roots and Mature Leaves
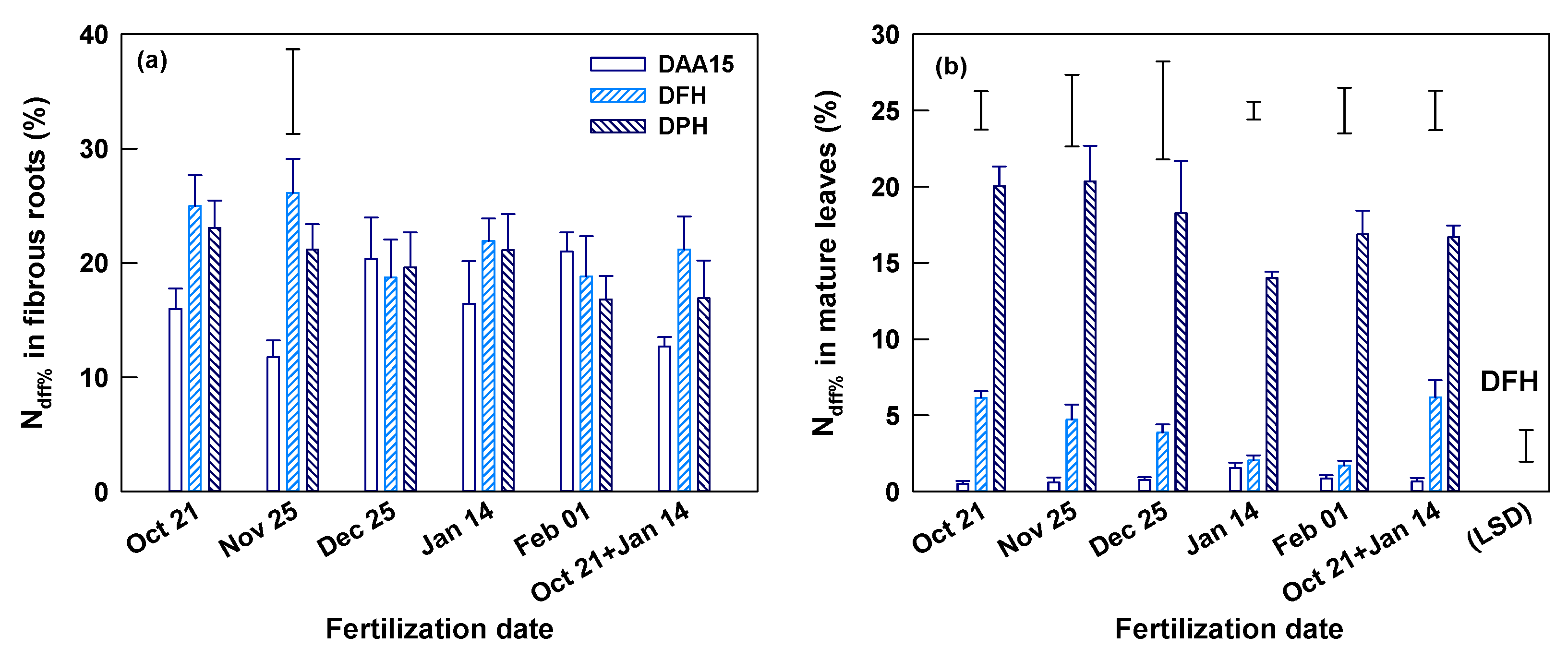
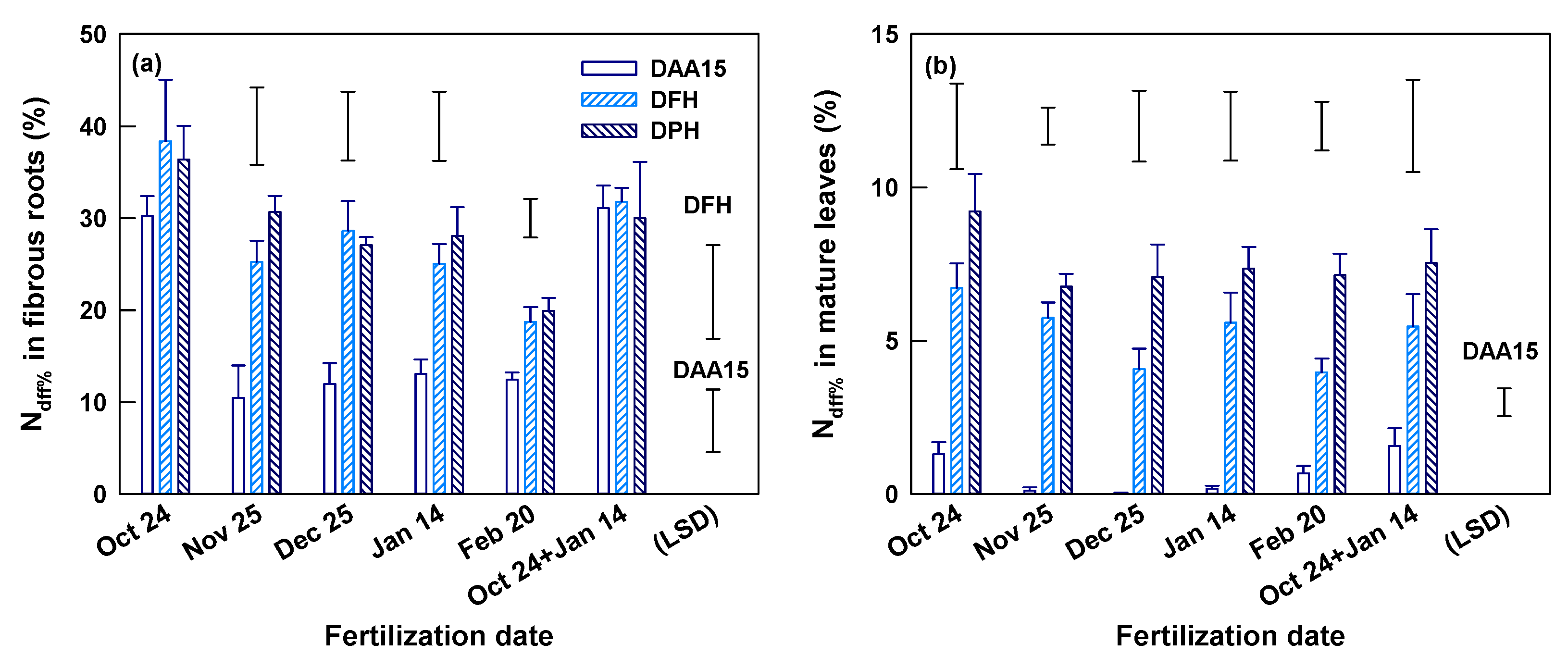
At Yongchuan, the fertilization dates insignificantly (p > 0.05) affected the total N concentrations of fibrous roots and mature leaves on DAA15, DFH (March 16) and DPH (May 5) (Table S1). The N concentrations of root and leaf also changed little in the process from DAA15 to DFH. However, compared to the previous two times, root N concentration on DPH decreased whereas the leaf N concentration increased, as indicated by their means across fertilization dates (p < 0.001). The Ndff% of fibrous roots reached 11.7–21.0% on DAA15 and changed insignificantly in the later times (DFH and DPH) in most treatments (Figure 3a). Only in the November application, the root Ndff% increased significantly from DAA15 to DFH. The Ndff% of fibrous roots in all three sampling times was insignificantly affected by fertilization dates. The Ndff% in mature leaves was generally low (only 0.5–1.6%) on DAA15, which was less than one-tenth of those in fibrous roots (Figure 3b and Table 2). In the duration from DAA15 to DFH, the leaf Ndff% increased to 1.7–6.2% and the increase was significant only in single and split October applications (Figure 3b). The leaf Ndff% at DFH was significantly affected by fertilization dates and was the highest in October application and the lowest in January and February applications. On DFH, the Ndff% leaf/root ratio (LNdff%/RNdff%) was also significantly smaller in the January and February than in other applications (Table 2). There was no significant difference in leaf Ndff% among fertilization dates on DPH. However, the leaf Ndff% increased (p < 0.05) by 2.7–9.9 folds to 14–20.4% and resulted in a sharp increase of LNdff%/RNdff% ratio during the period from DFH to DPH in all treatments (Figure 3b and Table 2).
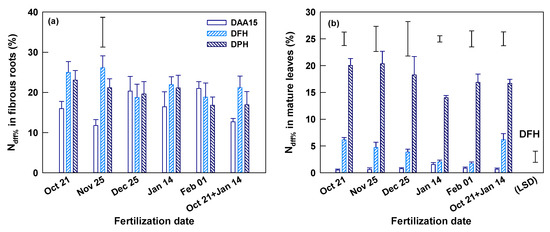
Figure 3.
Ndff% of fibrous root (a) and mature leaves (b) 15 days after 15N fertilization (DAA15), on the days of first harvest (DFH) and post-harvest (DPH) of spring shoots at Yongchuan, Chongqing province. The data are means and SE (n = 4). Single bars without symbols above columns represent LSD values indicating significant differences among DAA15, DFH, and DPH for each of the specified fertilization dates. A single bar without a symbol ticked with (LSD) in (b) represents LSD value indicating significant difference on DFH among fertilization dates.
Table 2.
Leaf/root ratio of Ndff% (LNdff%/RNdff%) measured 15 days after 15N application (DAA15), on the days of the first harvest (DFH) and post-harvest (DPH) of young spring shoots at Youngchuan (March 16 and May 4) and Xingyang (April 8 and May 12), respectively.
At Xingyang, total N concentration of fibrous roots was significantly affected by fertilization dates on DFH, but not on DAA15 and DPH (Table S1). Root N concentration was the highest in October application and lower in the applications of December, January, and February. The root N concentration changed little from DAA15 to DFH, but decreased significantly from DFH to DPH. The total N concentration of mature leaves was also insignificantly affected by fertilization dates. The leaf N concentration across fertilization dates declined at DPH in comparison with the previous two sampling times on DAA15 and DFH (Table S1). The root Ndff% reached 10.5–31.1% on DAA15 and continued to increase (p < 0.05) on DFH in all other applications except the October one (Figure 4a). On both DAA15 and DFH, root Ndff% was significantly affected by application dates and was the largest in single or split October application. From DFH to DPH, the root Ndff% had little change. On DAA15 the Ndff% in mature leaves was generally low (0.01–1.6%), but they were significantly affected by fertilization dates. Both the leaf Ndff% and LNdff%/RNdff% were the highest in October applications but very low in November, December, and January applications (Figure 4b and Table 2). The Ndff% of mature leaves increased to 4.0–6.7% on DFH (p < 0.05), and the increase continued until DPH (Figure 4b). Nevertheless, on both DFH and DPH, the leaf Ndff% and LNdff%/RNdff% were insignificantly affected by different fertilization dates.

Figure 4.
Ndff% of fibrous root and mature leaves 15 days after 15N fertilization (DAA15), on the days of first harvest (DFH) and post-harvest (DPH) of spring shoots at Xingyang, Henan province. The data are means and SE (n = 4). Single bars without symbols above columns represent LSD values indicating significant differences among DAA15, DFH, and DPH for each of the specified fertilization dates. A single bar without a symbol ticked with (LSD) represent LSD values indicating significant difference on DAA15, DFH (a), and DAA15 (b) among fertilization dates.
3.3. 15N in Young Spring Shoots
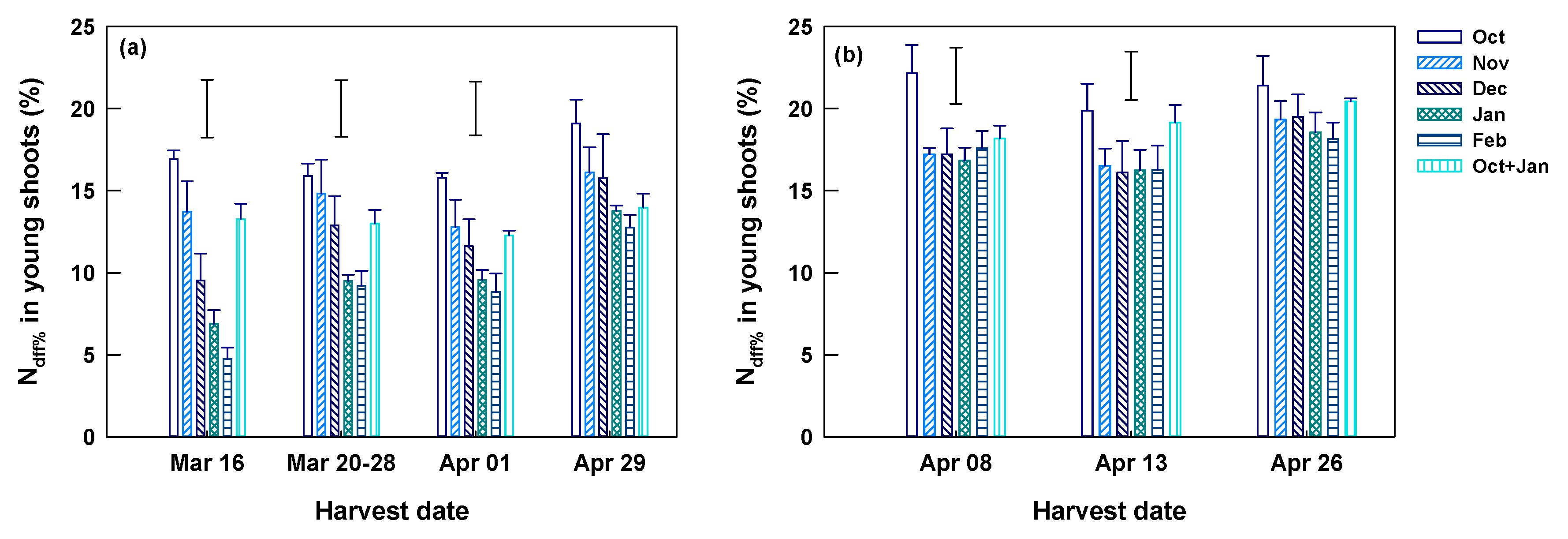
At Yongchuan, the harvest of young shoots started on March 16. Shoot N concentrations were the highest in the first harvest (March 16, mean 57.7 mg g−1) and decreased afterward to the lowest in the last harvest (April 29, mean 46.3 mg g−1) (Table 3). The young shoot Ndff% of the first harvest reached 4.8–16.9%, and they were significantly different among fertilization dates (Figure 5a). In the first harvest, young shoot Ndff% was the highest in the single October application, which was 3.6-fold of the lowest in February application. Split applications (October and January) had a slightly lower shoot Ndff% than the single October application. This tendency was continued in the following harvests on March 20–28 and April 1. Their difference became smaller (p > 0.05) in the last harvest (April 29). The mean Ndff% of young shoots increased from 10.9% of the first to 15.3% of the last harvest round (Table 3). The increases were particularly remarkable in the February and January applications (Figure 5a). Split applications had a significantly lower Ndff% than the single application.
Table 3.
Mean N concentration and Ndff% of young shoots across fertilization dates.
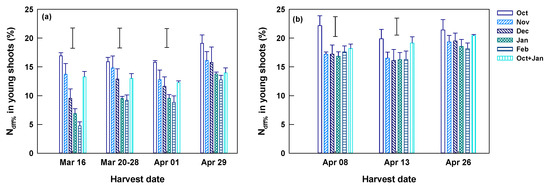
Figure 5.
Ndff% of spring young shoots harvested on different dates at Yongchuan (a) and Xingyang (b) from tea plants receiving 15N enriched urea individually in October, November, December, Janauary, and February and split in October and January. Data are means and SE (n = 4). Bars without data points above columns represent LSD values indicating significant (p < 0.05) difference among fertilization dates on each of the specified harvest dates.
At Xingyang, the harvest of young shoots started on April 8, a time much later than Yongchuan for lower winter and spring temperature. Again, the mean shoot N concentration was significantly higher in the first than in later harvests (Table 3). The young shoot Ndff% of the first harvest reached 16.8–22.2% and was significantly different among fertilization dates (Figure 5b). Ndff% was the highest in the single October application, which was 1.2–1.3 fold of those applied in other months. The split application had significantly lower Ndff% than that of the single October application. This tendency was continued in the following harvest on April 13, with smaller but significant differences (Figure 5b). The young shoot Ndff% of the last harvest (on April 26) was insignificantly different among fertilization dates. The mean Ndff% across fertilization dates of the last harvest was higher than two tier harvests (Table 3).
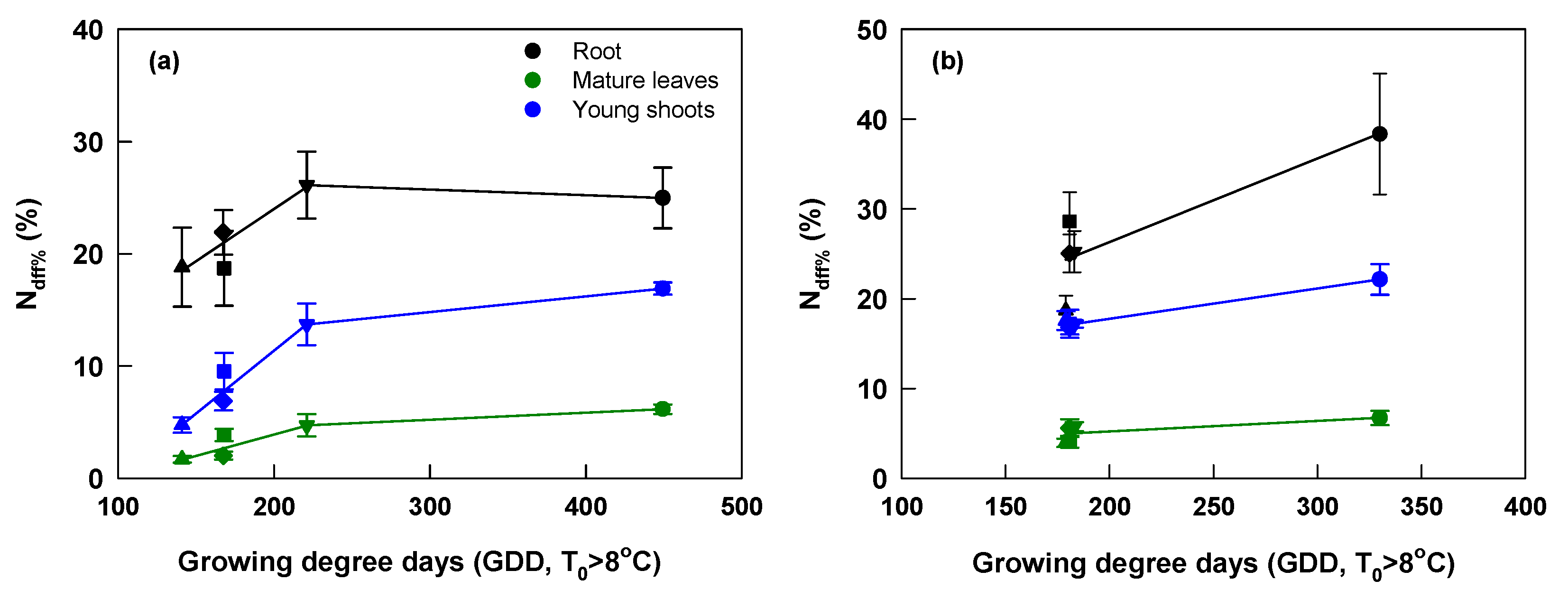
The Ndff% of root, mature leaves, and young shoots on the DFH was closely associated with the change of growing degree days at both sites (Figure 6). At Yongchuan, their relationship could be well described by two regression lines and Ndff% reached the maximum at the GDD of about 220 °C•d. At Xingyang, their relationship was described by one regression line as there was little difference among the last four fertilization dates. At this site, Ndff% reached the maximum at the GDD of about 330 °C•d.
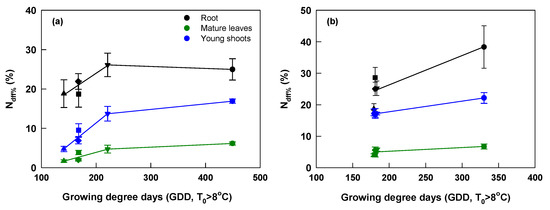
Figure 6.
Relationship of Ndff% of root, mature leaves, and young shoots on the day of the first harvest (DFH) with growing degree days (T > 8 °C, GDD) between fertilization dates and DFH at Yongchuan (a) and Xingyang (b). Black, green and blue colors of symbols and lines indicate root, mature leaves, and young shoots, respectively. Symbols ○, ▽, □, ◇, and △ indicate fertilization dates in October, November, December, January, and February, respectively.
4. Discussion
In the present study, we applied 15N enriched urea on different dates from late October through February at two tea plantations. These two sites were selected for their different thermo conditions during November and March, but very similar conditions in other months (Figure 1b). We measured 15N uptake and utilization after a short (DAA15) and two longer times (DFH and DPH) to investigate the interacting effect of fertilization date and thermo conditions on fertilizer N uptake and utilization in the following spring.
4.1. Temperature and the Short-Term Autumn 15N Absorption and Upward Transport
Temperature is a key factor affecting the rate of nutrient uptake. Low temperature reduces nutrient uptake by decreasing plant growth rate and thereby nutrient demand and root respiration then less energy supply [38]. With decreasing temperature in October, tea plants in subtropical area stop vegetative growth and turn into vegetative dormancy. Demand of nutrients therefore largely decreased, but persisted as indicated by the present experiment that fertilizer N applied in the months between October and February was taken up by fibrous roots at both sites (Figure 3, Figure 4 and Figure 5). This finding coincides with previous experiments showing N uptake in the late autumn and winter dormancy in the absence of active shoot growth in subtropical ever-green woody plants such as tea and citrus [18,30]. A significant difference between application dates was observed at Xingyang (Figure 4), where Ndff% in root and LNdff%/RNdff% within DAA15 were much lower in November, December, and January than other dates. This is likely linked to different thermo conditions. At this site, periodic air temperatures within DDA15 occasionally fell below zero and were conspicuously lower in application dates other than October (Figure 2c), possibly leading to little N uptake. Previous work showed that N uptake by cool-season turfgrasses is greatly decreased when the temperatures approach 0 °C [39]. However, at Yongchuan, we observed little difference in 15N uptake and transport with DAA15 among fertilization dates (Figure 3). At this site, the periodic air temperature never fell below zero (3.7–8.5 °C), even in the coldest December and January (Figure 1b and Figure 2a). There is no information about the lowest temperature below which N uptake by tea plant root ceases. However, it is assumed that the air/soil temperature at Yongchuan within DAA15 of all applications met the minimum requirement for the nutrient uptake as a result of weak demand at vegetative dormancy.
Within DAA15, Ndff% of mature leaves was below a tenth of those in roots, showing only weakly upward transport from root to above-ground. The low demand of above-ground at dormancy, reduced ability of roots to conduct water, and weak transpiration flow might all have contributed to the slow upward 15N translocation under low temperature [40,41]. On DAA15, the root Ndff% did not differ significantly between the two sites for each specific application date, whereas leaf Ndff% was greater (p < 0.05) at Yongchuan than Xingyang in the applications of November, December, and January. The LNdff%/RNdff% ratios in these applications were disproportionately low at Xingyang, possibly suggesting that 15N transport from the root to above-ground had been particularly inhibited by the lower temperature at this site (Figure 2 and Table 2).
4.2. Thermo Condition and the Accumulation of 15N Reserves during Winter Dormancy
For ever-green trees nitrogen absorbed in winter, dormancy is stored in plants as N reserves [18]. In the present work, we were unable to precisely evaluate the effect of fertilization timing on the size of 15N reserves in tea plants at the time when young spring shoots started to grow. On DFH, 15N reserves might have already been utilized in young shoots. However, the build-up process of N reserves and the contribution of fertilizer 15N could be partly reflected by the change of 15N in fibrous roots and mature leaves between DAA15 and DFH. Root Ndff% changed only slightly and insignificantly between DAA15 and DFH at Yongchuan in all applications, except November’s (Figure 3a). There was also little change in the root Ndff% of October applications at Xingyang during DAA15 and DFH. It therefore appeared that the root Ndff% reached their maximum within DAA15 in these applications. In contrast, at Xingyang in all other applications except October’s and at Yongchuan in November’s application the Ndff% of fibrous roots had not reached their maximum on DAA15, thereafter, continued to increase until DFH (Figure 3a and Figure 4a). In all applications at both sites, Ndff% of mature leaves increased by 5–8 fold, and LNdff%/RNdff% ratio increased by 3.6–6 fold during the period from DAA15 to DFH (Figure 3b, Figure 4b and Table 2). These increases likely were the result of an accumulating effect of 15N upward transport from roots, a process which also became stronger in response to rising temperature from DAA15 to DFH (Figure 1b) [30]. Taking fibrous roots and mature leaves together, it was concluded that N reserves were greater in the October application than those of other months at both sites (Figure 3b and Figure 4b).
4.3. Thermo Condition and 15N Utilization in the following Young Spring Shoot
From DFH to DPH, N concentrations of fibrous roots at both sites decreased significantly (Table S1). Such a decrease of N concentration during spring growth was frequently observed in other trees and might be the result of remobilization to young shoots as the new sink [42]. On the other hand, N concentrations of mature leaves showed the opposite changes at the two sites (Table S1): it significantly increased at Yongchuan but decreased at Xingyang. The Ndff% of mature leaves and their ratios to fibrous roots corresponded to these changes, with a more remarkable increase at Yongchuan than at Xingyang (Figure 3b, Figure 4b and Table 2). The changes of leaf total N and 15N amounts were estimated based on Ndff% values assuming constant biomass between DFH and DPH. We found that about 65% (60–81%) of the increase of leaf N content could be accounted for by the change of 15N amount at Yongchuan. Thus, the concomitant root 15N uptake had contributed mainly to the increase of leaf N from DFH to DPH, indicating that 15N had been allocated to both young shoots and mature leaves. However, at Xingyang, the N content of mature leaves decreased, suggesting a net N export to young shoots during this period. Meanwhile, leaf 15N content increased only minimum (by 0.24-fold), suggesting that 15N had been predominantly allocated to young shoots. The differing 15N allocation to mature leaves and young shoots between the two sites might be attributed to different inherent properties of the two varieties which deserves future work.
Young shoot Ndff% was markedly affected by fertilization dates (Figure 5). Young shoot 15N was contributed from the remobilization of N reserves before bud breaking and the current uptake during the spring growth. Application in October improved N absorption throughout winter and early spring, which was stored in the tea plants and contributed to greater accumulation of N reserves in roots and mature leaves than other applications (Figure 3 and Figure 4). The largest difference of Ndff% among dates of applications occurred in the first harvest (March 16 and April 8) at both sites, showing that N fertilizer applied in late October makes a relatively greater contribution to the early spring tea (Figure 5). This finding is in line with previous works demonstrating that the N taken up in the late phenological stages (i.e., in autumn before winter dormancy or after the harvest of fruits) is preferentially remobilized to support early spring growth [22,23,24,25,43]. The decreasing total N concentrations of young shoots from the first to the latest harvest, in contrast with the increasing Ndff% at both sites reflected weakening support of N reserves, which progressively depleted (Table 3). The difference of shoot Ndff% between application dates became smaller (Figure 5 and Table 3), likely suggesting the increasing ability of concurrent uptake in the late spring, and contribution to new spring growth with rising temperature [7,44]. The difference of young shoot Ndff% among fertilization dates was closely associated with the GDD values between dates of fertilization and the first harvest (Figure 2 and Figure 6). At Yongchuan where there was large difference in GDD, there were significant different Ndff% of young shoots among different fertilization dates. At Xingyang where there was different GDD and shoot Ndff% only among October and other applications. Our finding confirms a few earlier works showing that air or soil GDD depicts the combining effect of temperature and residence time and thereby well predicted 15N uptake by citrus, apple and tea plants [30,45,46].
There were clear differences of Ndff% in fibrous roots, mature leaves, and young spring shoots between the single October application and split applications in October and January. The split application supplied only half of the 15N in October, likely weakened the N reserves and subsequently, the amount remobilized to young spring shoots. On the other hand, splitting another half to January application has shortened the residence time of this N part in the root zone and cumulative heat condition estimated from GDD [30,45,46]. Our work suggests that a decrease or withdrawal of autumn fertilization cannot be compensated by an later application, an observation has been previously shown in young peach trees [17].
5. Conclusions
In summary, application of N fertilizer in October relative to other later months most significantly improves the N absorption and accumulation of plant N reserves throughout the winter dormancy, and consequently contributes more significantly to the early spring tea. Such timing effect was closely associated with the growing degree days between the dates of fertilization and harvest of young shoots, which represents the combining effect of the temperature and the residence time of N fertilizer in the soil. The result showed the determining effect of thermal condition on the efficiency of fertilizer N applied in the autumn–winter period and provides practical information to assist the optimization of N nutrition management in green tea production. Due to the relatively short experimental duration, future works at more sites and for a longer period would further deepen our understanding about the interacting effect of thermal condition and N utilization by tea plants.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/horticulturae7120544/s1, Figure S1: Cumulative rainfall from October 1 to May 30 at Yongchuan, Chongqing (CQ) and Xingyang, Henan (HN) province, China, Table S1: Total N concentration of fibrous roots and mature leaves measured 15 days after 15N application (DAA15), on the days of the first harvest (DFH) and post-harvest (DPH) of young spring shoots at Yongchuan (March 16 and May 4) and Xingyang (April 8 and May 12), respectively.
Author Contributions
Conceptualization and experimental design, J.R. and L.M.; field investigation, sample collection and preparation, S.J., M.D., L.L. and Z.X. sample analysis, L.M., data curation, J.R., L.M., S.J. and M.D. writing of manuscript, J.R., L.M, Z.X. and L.L.; funding acquisition, J.R., L.L. and Z.X. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Science Foundation of China under Grant 32172634, the Earmarked Fund for China Agriculture Research System under Grant CARS 19, the Agricultural Science and Technology Innovation Program under Grant CAAS-ASTIP-TRICAAS, and Yunnan provincial special fund for construction of science and technology innovation base (202102AE090038).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, Q.; Ruan, J.Y. Tea: Analysis and Tasting. In The Encyclopedia of Food and Health; Caballero, B., Finglas, P., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; Volume 5, pp. 256–267. [Google Scholar]
- Xu, W.; Song, Q.; Li, D.; Wan, X. Discrimination of the production season of Chinese green tea by chemical analysis in combination with supervised pattern recognition. J. Agric. Food Chem. 2012, 60, 7064–7070. [Google Scholar] [CrossRef]
- Dai, W.; Qi, D.; Yang, T.; Lv, H.; Guo, L.; Zhang, Y.; Zhu, Y.; Peng, Q.; Xie, D.; Tan, J.; et al. Nontargeted analysis using ultraperformance liquid chromatography–quadrupole time-of-flight mass spectrometry uncovers the effects of harvest season on the metabolites and taste quality of tea (Camellia sinensis L.). J. Agric. Food Chem. 2015, 63, 9869–9878. [Google Scholar] [CrossRef]
- Liu, J.W.; Zhang, Q.F.; Liu, M.Y.; Ma, L.F.; Shi, Y.Z.; Ruan, J.Y. Metabolomic analyses reveal distinct change of metabolites and quality of green tea during the short duration of a single spring season. J. Agric. Food Chem. 2016, 64, 3302–3309. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Yang, J.; Xie, X.; Lin, C. Over-supply of tea in China—A marketing analysis. Acta Tea Sin 2017, 58, 75–79. [Google Scholar]
- Millard, P.; Grelet, G.A. Nitrogen storage and remobilization by trees: Ecophysiological relevance in a changing world. Tree Physiol. 2010, 30, 1083–1095. [Google Scholar] [CrossRef] [Green Version]
- Bazot, S.; Fresneau, C.; Damesin, C.; Barthes, L. Contribution of previous year’s leaf N and soil N uptake to current year’s leaf growth in sessile oak. Biogeosciences 2016, 13, 3475–3484. [Google Scholar] [CrossRef] [Green Version]
- Ueda, M.U.; Mizumachi, E.; Tokuchi, N. Foliage nitrogen turnover: Differences among nitrogen absorbed at different times by Quercus serrata saplings. Ann. Bot. 2011, 108, 169–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Sun, M.; Qi, Y.; Liu, S. Remobilization of storage nitrogen in young pear trees grafted onto vigorous rootstocks (Pyrus betulifolia). Horticulturae 2021, 7, 148. [Google Scholar] [CrossRef]
- Ueda, M.; Tokuchi, N.; Hiura, T. Soil nitrogen pools and plant uptake at sub-zero soil temperature in a cool temperate forest soil: A field experiment using 15N labeling. Plant Soil 2015, 392, 205–214. [Google Scholar] [CrossRef]
- Fife, D.N.; Nambiar, E.K.S.; Saur, E. Retranslocation of foliar nutrients in evergreen tree species planted in a Mediterranean environment. Tree Physiol. 2008, 28, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legaz, F.; Serna, M.D.; Primo-Millo, E. Mobilization of the reserve N in citrus. Plant Soil 1995, 173, 205–210. [Google Scholar] [CrossRef]
- Uscola, M.; Villar-Salvador, P.; Gross, P.; Maillard, P. Fast growth involves high dependence on stored resources in seedlings of Mediterranean evergreen trees. Ann. Bot. 2015, 115, 1001–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.-H.; Li, K.-T. Dormant season fertigation promotes photosynthesis, growth, and flowering of ‘Blueshower’ rabbiteye blueberry in warm climates. Hortic. Environ. Biotechnol. 2015, 56, 756–761. [Google Scholar] [CrossRef]
- Pescie, M.A.; Borda, M.P.; Ortiz, D.P.; Landriscini, M.R.; Lavado, R.S. Absorption, distribution and accumulation of nitrogen applied at different phenological stages in southern highbush blueberry (Vaccinium corymbosum interspecific hybrid). Sci. Hortic. 2018, 230, 11–17. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, V.; Sánchez, E.; Tognetti, J. Timing of nitrogen fertilization influences color and anthocyanin content of apple (Malus domestica Borkh. cv ‘Royal Gala’) fruits. Int. J. Fruit Sci. 2011, 11, 364–375. [Google Scholar] [CrossRef]
- Jordan, M.O.; Wendler, R.; Millard, P. Autumnal N storage determines the spring growth, N uptake and N internal cycling of young peach trees. Trees 2012, 26, 393–404. [Google Scholar] [CrossRef]
- Kato, T. Nitrogen metabolism and utilization in citrus. In Horticultural Reviews Volume 8; Conover, C.A., Van Wann, E., Zimmerman, R.H., Eds.; AVI Publishing Company INC.: Westport, CT, USA, 2011; pp. 181–216. [Google Scholar]
- Dong, S.; Cheng, L.; Scagel, C.F.; Fuchigami, L.H. Timing of urea application affects leaf and root N uptake in young Fuji/M.9 apple trees. J. Hortic. Sci. Biotechnol. 2005, 80, 116–120. [Google Scholar] [CrossRef]
- Martinez-Alcantara, B.; Quinones, A.; Primo-Millo, E.; Legaz, F. Seasonal changes in nitrate uptake efficiency in young potted citrus trees. J. Agric. Sci. 2012, 4, 11–19. [Google Scholar] [CrossRef]
- Khemira, H.; Righetti, T.L.; Azarenko, A.N. Nitrogen partitioning in apple as affected by timing and tree growth habit. J. Hortic. Sci. Biotechnol. 1998, 73, 217–223. [Google Scholar] [CrossRef]
- Martínez-Alcántara, B.; Quiñones, A.; Legaz, F.; Primo-Millo, E. Nitrogen-use efficiency of young citrus trees as influenced by the timing of fertilizer application. J. Plant Nutr. Soil Sci. 2012, 175, 282–292. [Google Scholar] [CrossRef]
- San-Martino, L.; Sozzi, G.O.; San-Martino, S.; Lavado, R.S. Isotopically-labelled nitrogen uptake and partitioning in sweet cherry as influenced by timing of fertilizer application. Sci. Hortic. 2010, 126, 42–49. [Google Scholar] [CrossRef]
- Rivera, R.; Bañados, P.; Ayala, M. Distribution of 15N applied to the soil in the ‘Bing’/‘Gisela®6′ sweet cherry (Prunus avium L.) combination. Sci. Hortic. 2016, 210, 242–249. [Google Scholar] [CrossRef]
- Quartieri, M.; Millard, P.; Tagliavini, M. Storage and remobilisation of nitrogen by pear (Pyrus communis L.) trees as affected by timing of N supply. Eur. J. Agron. 2002, 17, 105–110. [Google Scholar] [CrossRef]
- Niederholzer, F.J.A.; DeJong, T.M.; Saenz, J.L.; Muraoka, T.T.; Weinbaum, S.A. Effectiveness of fall versus spring soil fertilization of field-grown peach trees. J. Am. Soc. Hortic. Sci. 2001, 126, 644–648. [Google Scholar] [CrossRef]
- Roccuzzo, G.; Scandellari, F.; Allegra, M.; Torrisi, B.; Stagno, F.; Mimmo, T.; Zanotelli, D.; Gioacchini, P.; Millard, P.; Tagliavini, M. Seasonal dynamics of root uptake and spring remobilisation of nitrogen in field grown orange trees. Sci. Hortic. 2017, 226, 223–230. [Google Scholar] [CrossRef]
- Okano, K.; Matsuo, K. Seasonal changes in uptake, distribution and redistribution of 15N-nitrogen in young tea (Camellia sinensis L.) plants. Jpn. J. Crop Sci. 1996, 65, 707–713. [Google Scholar] [CrossRef]
- Watanabe, I. Effect of nitrogen fertilizer application at different stages on the quality of green tea. Soil Sci. Plant Nutr. 1995, 41, 763–768. [Google Scholar] [CrossRef]
- Ma, L.F.; Shi, Y.Z.; Ruan, J.Y. Nitrogen absorption by field-grown tea plants (Camellia sinensis) in winter dormancy and utilization in spring shoots. Plant Soil 2019, 442, 127–140. [Google Scholar] [CrossRef]
- Ruan, J.Y.; Haerdter, R.; Gerendas, J. Impact of nitrogen supply on carbon/nitrogen allocation: A case study on amino acids and catechins in green tea [Camellia sinensis (L.) O. Kuntze] plants. Plant Biol. 2010, 12, 724–734. [Google Scholar] [CrossRef]
- Ni, K.; Liao, W.; Yi, X.; Niu, S.; Ma, L.F.; Shi, Y.Z.; Zhang, Q.; Liu, M.Y.; Ruan, J.Y. Fertilization status and reduction potential in tea gardens of China. J. Plant Nutr. Fert. 2019, 25, 421–432. [Google Scholar]
- Ma, L.; Yang, X.; Shi, Y.; Yi, X.; Ji, L.; Cheng, Y.; Ni, K.; Ruan, J. Response of tea yield, quality and soil bacterial characteristics to long-term nitrogen fertilization in an eleven-year field experiment. Appl. Soil Ecol. 2021, 166, 103976. [Google Scholar] [CrossRef]
- Liu, M.Y.; Burgos, A.; Ma, L.; Zhang, Q.; Tang, D.; Ruan, J. Lipidomics analysis unravels the effect of nitrogen fertilization on lipid metabolism in tea plant (Camellia sinensis L.). BMC Plant Biol. 2017, 17, 165. [Google Scholar] [CrossRef] [Green Version]
- Carranca, C.; Brunetto, G.; Tagliavini, M. Nitrogen nutrition of fruit trees to reconcile productivity and environmental concerns. Plants 2018, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Sulas, L.; Mercenaro, L.; Campesi, G.; Nieddu, G. Different cover crops affect nitrogen fluxes in Mediterranean vineyard. Agron. J. 2017, 109, 2579–2585. [Google Scholar] [CrossRef]
- Johnson, I.R.; Thornley, J.H.M. Temperature dependence of plant and crop process. Ann. Bot. 1985, 55, 1–24. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; King, J.S. Effects of Soil Temperature on Nutrient Uptake. In Nutrient Acquisition by Plants. An Ecological Perspective; BassiriRad, H., Ed.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2005; pp. 290–323. [Google Scholar]
- Lloyd, D.T.; Soldat, D.J.; Stier, J.C. Low-temperature nitrogen uptake and use of three cool-season turfgrasses under controlled environments. HortScience 2011, 46, 1545–1549. [Google Scholar] [CrossRef] [Green Version]
- Wan, X.; Landhäusser, S.M.; Zwiazek, J.J.; Lieffers, V.J. Root water flow and growth of aspen (Populus tremuloides) at low root temperatures. Tree Physiol. 1999, 19, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Engels, C.; Marschner, H. Effect of root zone temperature and shoot demand on nitrogen translocation from the roots to the shoot in maize supplied with nitrate or ammonium. Plant Physiol. Biochem. 1996, 34, 144–157. [Google Scholar]
- Lamaze, T.; Pasche, F.; Pornon, A. Uncoupling nitrogen requirements for spring growth from root uptake in a young evergreen shrub (Rhododendron ferrugineum). New Phytol. 2003, 159, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Lim, C.S.; Kang, S.M.; Cho, J.L. Root storage of nitrogen applied in autumn and its remobilization to new growth in spring of persimmon trees (Diospyros kaki cv. Fuyu). Sci. Hortic. 2009, 119, 193–196. [Google Scholar] [CrossRef]
- Dong, S.; Scagel, C.F.; Cheng, L.; Fuchigami, L.H.; Rygiewicz, P.T. Soil temperature and plant growth stage influence nitrogen uptake and amino acid concentration of apple during early spring growth. Tree Physiol. 2001, 21, 541–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholberg, J.M.S.; Parsons, L.R.; Wheaton, T.A.; McNeal, B.L.; Morgan, K.T. Soil temperature, nitrogen concentration, and residence time affect nitrogen uptake efficiency in citrus. J. Environ. Qual. 2002, 31, 759–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neilsen, D.; Millard, P.; Herbert, L.C.; Neilsen, G.H.; Hogue, E.J.; Parchomchuk, P.; Zebarth, B.J. Remobilization and uptake of N by newly planted apple (Malus domestica) trees in response to irrigation method and timing of N application. Tree Physiol. 2001, 21, 513–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).