
Evaluation of 130 Eggplant (Solanum melongena L.) Genotypes for Future Breeding Program Based on Qualitative and Quantitative Traits, and Various Genetic Parameters

 , ,
, ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Site
2.2. Experimental Materials, Treatments, Desing and Procedures
2.3. Data Recorded
Quantitative Traits Measuring
2.4. Statistical Analysis
3. Results
3.1. Qualitative Traits
3.2. Variability in Growth and Foliage
3.3. The Analysis of Variance and Frequency Distribution of Quantitative Traits
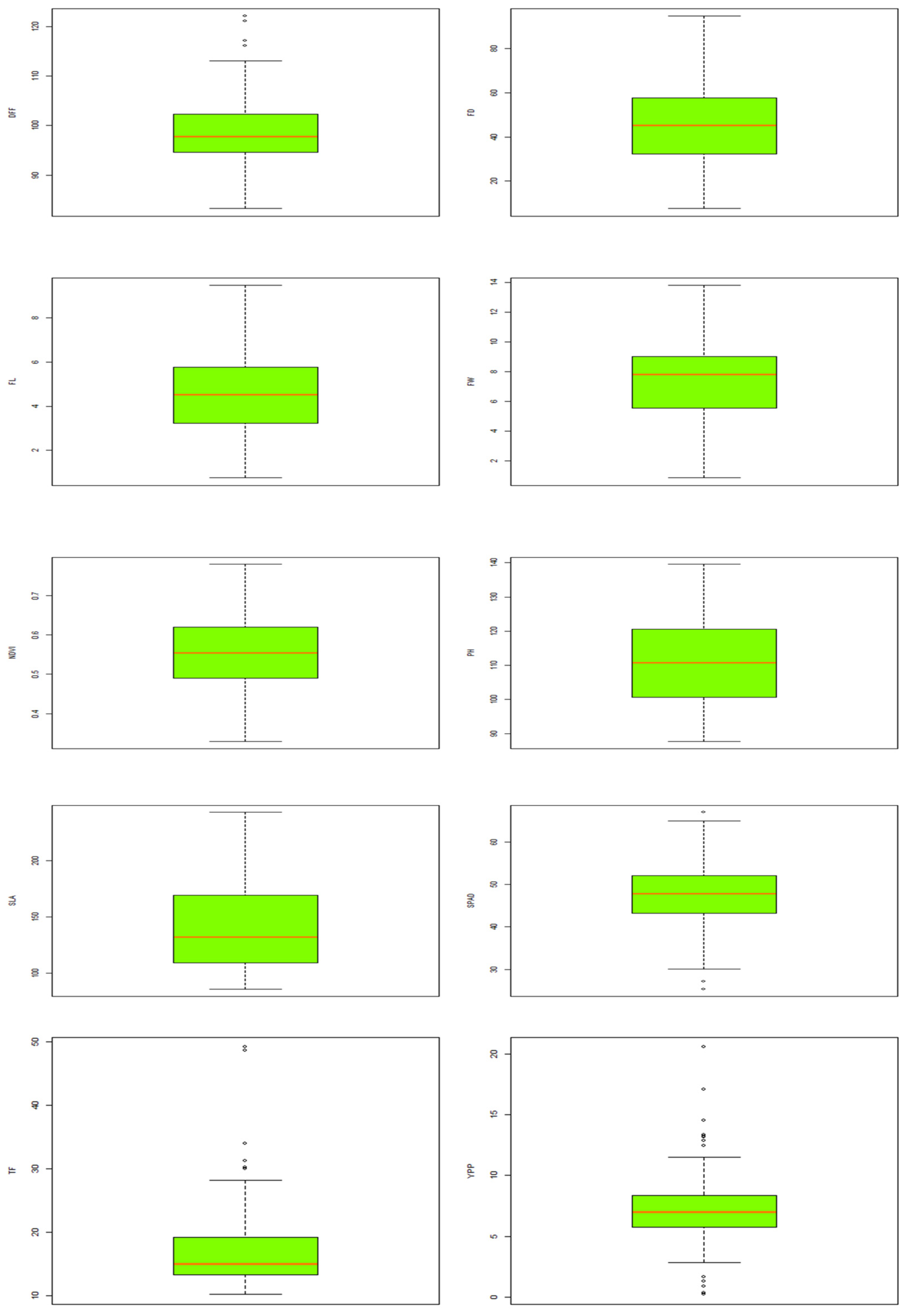
3.4. Descriptive Statistic of the Traits
3.5. Variability in Fruit Traits
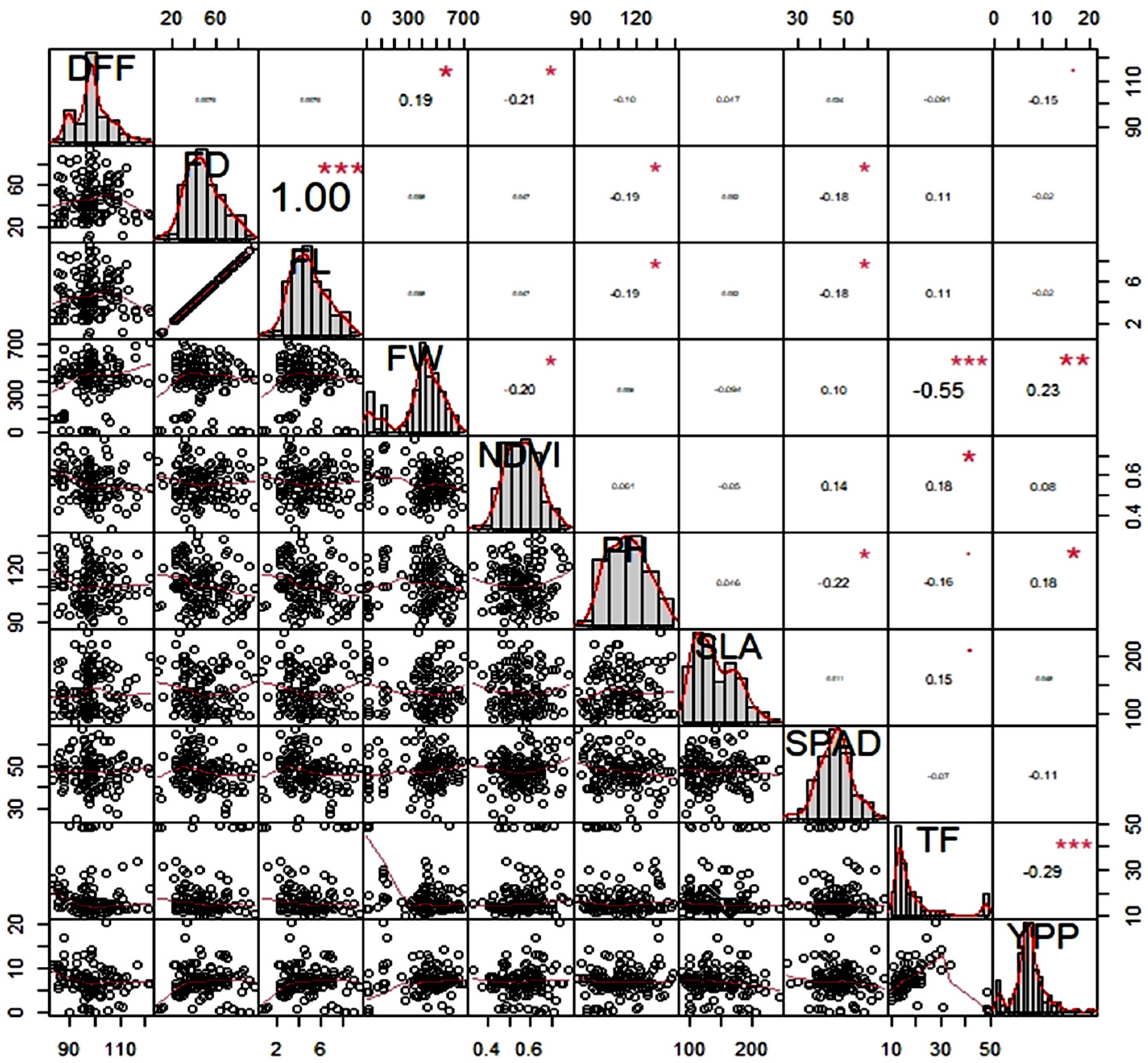
3.6. Analysis of Correlation Matrix
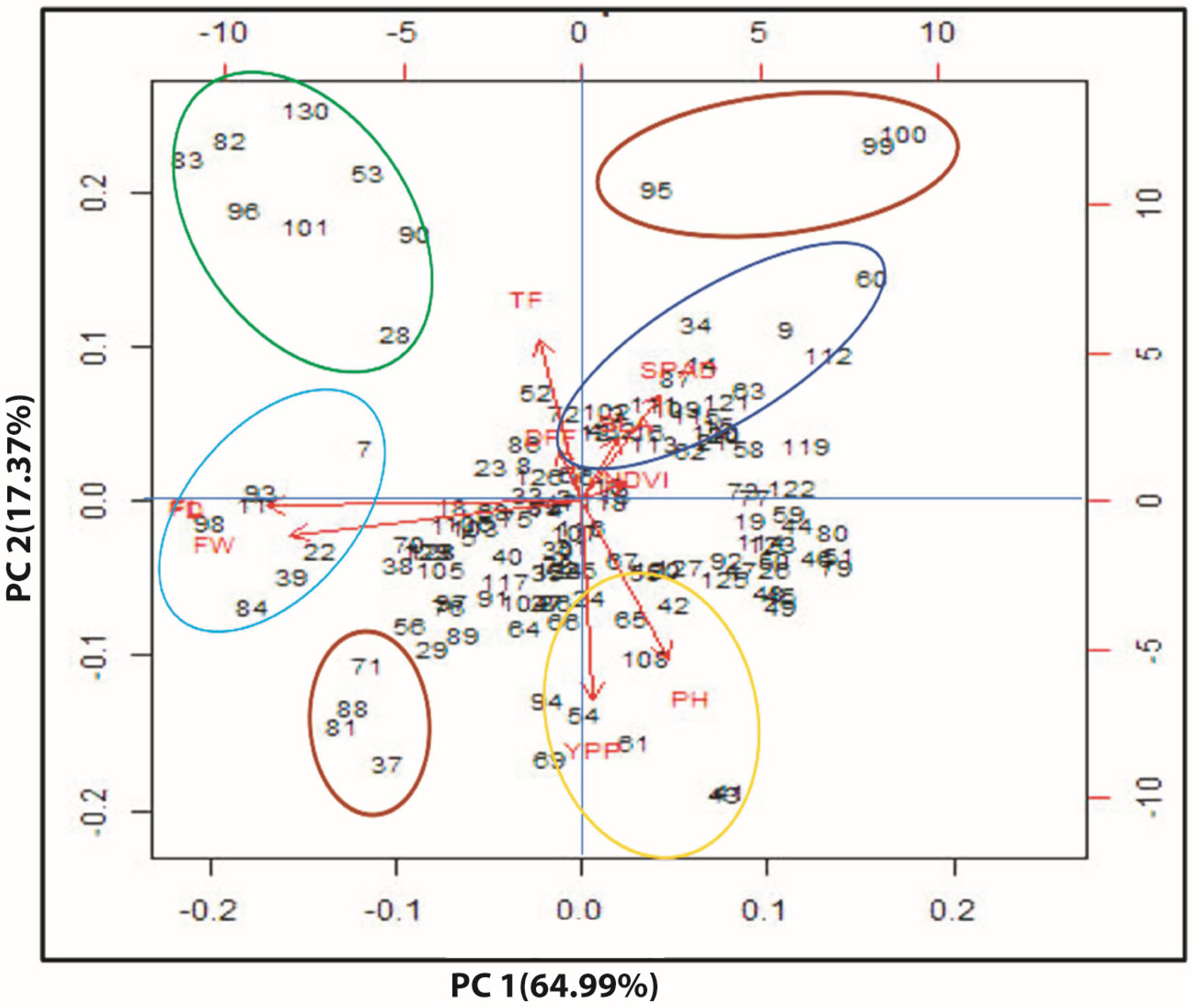
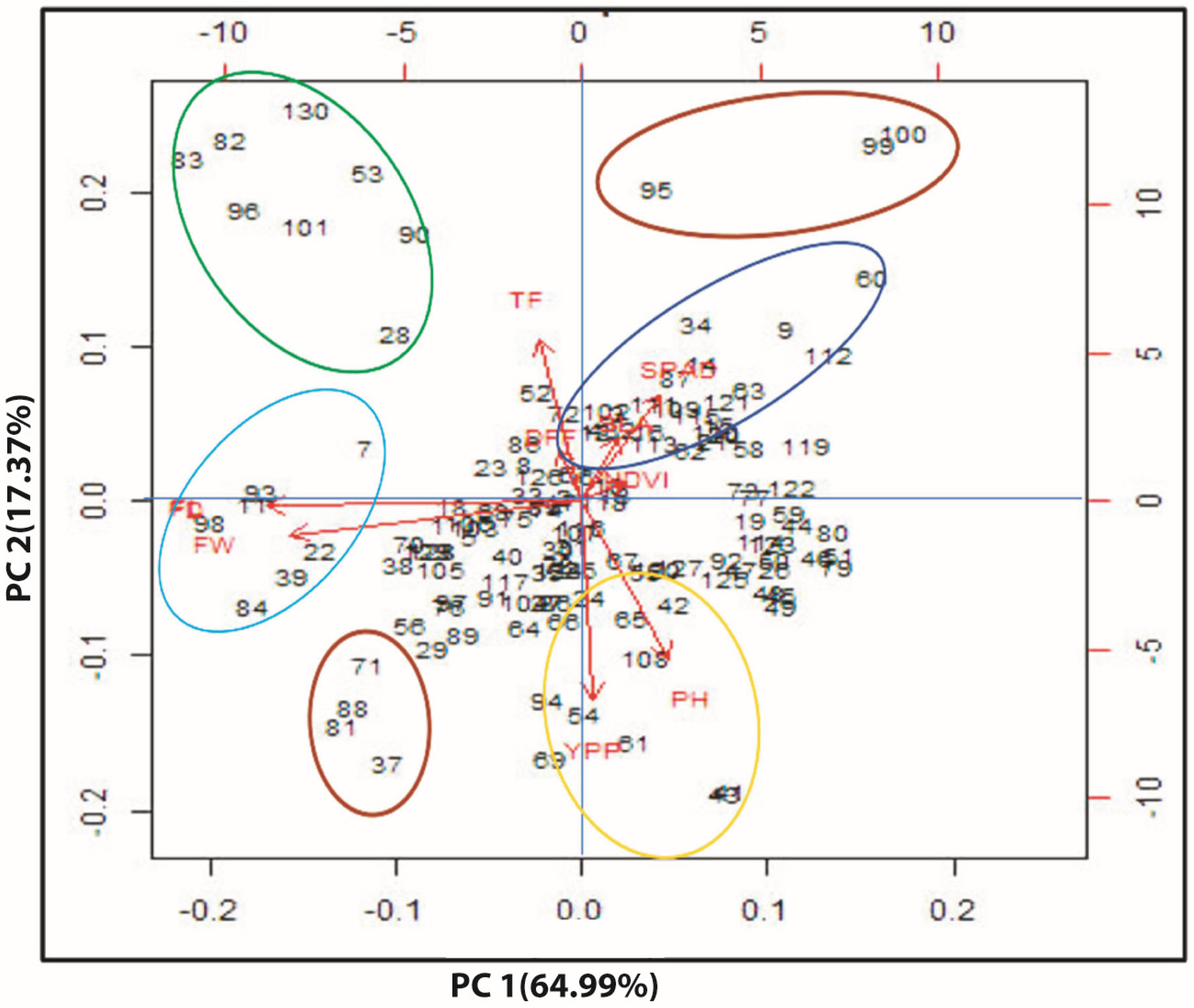
3.7. Multivariate Analysis
3.8. Heatmap Analysis
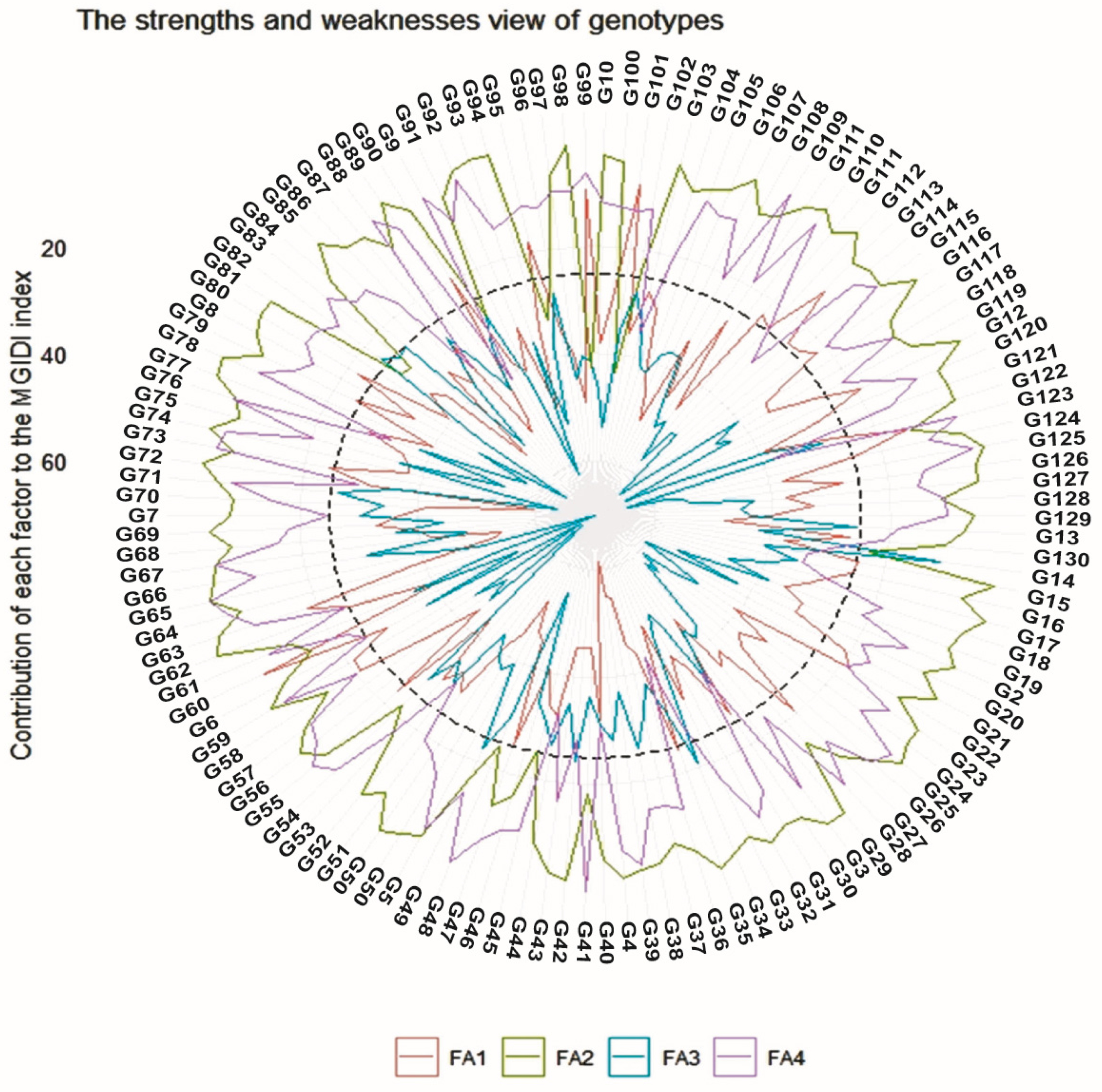
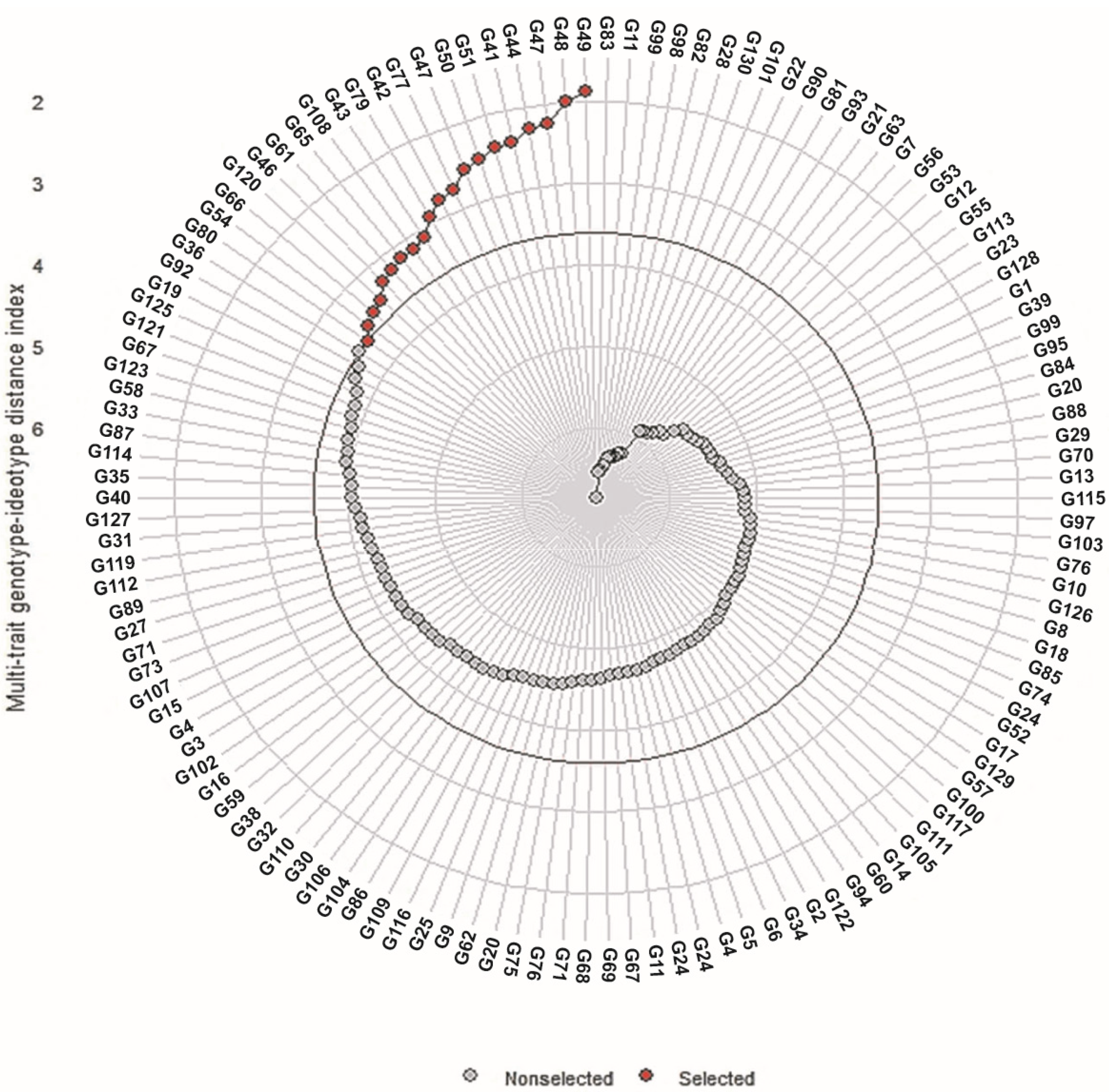
3.9. Multi-Trait Index Based on Factor Analysis and Genotype-Ideotype Distance (MGIDI)
4. Discussion
4.1. Qualitative Traits
4.2. Quantitative Traits
4.2.1. Genetic Components
4.2.2. Correlation Matrix
4.3. Multivariate Statistical
4.4. Multi-Trait Index Based on Factor Analysis and Genotype-Ideotype Distance (MGIDI)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aubriot, X.; Daunay, M.-C. Eggplants and relatives: From exploring their diversity and phylogenetic relationships to conservation challenges. In The Eggplant Genome; Springer: Berlin/Heidelberg, Germany, 2019; pp. 91–134. [Google Scholar]
- Minardi, A. Food Security in Developing Countries: The Case of “Production of Appropriate Food: Sufficient, Safe and Sustainable” Project. Ph.D Thesis, (Ph.D. in Agro-Food System, Academic Year 2017/2018). Catholic University of Sacred Heart, Via Emilia Parmense, Piacenza, Italy, 2019. Available online: http://tesionline.unicatt.it/bitstream/10280/67851/1/tesiphd_completa_Minardi.pdf (accessed on 15 July 2021).
- Bhagirath, C.; Kadambini, G. The Development and Regulation of Bt Brinjal in India (Eggplant/Aubergine); International Service for the Acquisition of Agri-biotech Applications: Ithaca, NY, USA, 2009. [Google Scholar]
- BBS. Statistical Yearbook of Bangladesh. Bangladesh Bureau of Statistics; Ministry of Planning, Government of the Peoples’ Republic of Bangladesh: Dhaka, Bangladesh, 2020; p. 139.
- Prabakaran, R.K.-S.; Balakrishnan, S.; RameshKumar, S.; Arumugam, T.; Anandakumar, C. Genetic diversity, trait relationship and path analysis in eggplant landraces. Electron. J. Plant 2015, 6, 831–837. [Google Scholar]
- Van der Weerden, G.M.; Barendse, G.W. A web-based searchable database developed for the EGGNET project and applied to the Radboud University Solanaceae database. Acta Hortic. 2007, 745, 503–506. [Google Scholar] [CrossRef]
- Boyaci, H.F.; Topcu, V.; Akin, T.; Yildirim, I.K.; Mehmet, O.; Aktas, A. Morphological and molecular characterization and relationships of Turkish local eggplant heirlooms. Not. Bot. Horti Agrobot. Cluj Napoca 2015, 43, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Mat Sulaiman, N.N.; Rafii, M.Y.; Duangjit, J.; Ramlee, S.I.; Phumichai, C.; Oladosu, Y.; Datta, D.R.; Musa, I. Genetic variability of eggplant germplasm evaluated under open field and glasshouse cropping conditions. Agronomy 2020, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Falcón, J.; Prohens, J.; Vilanova, S.; Nuez, F. Diversity in commercial varieties and landraces of black eggplants and implications for broadening the breeders’ gene pool. Ann. Appl. Biol. 2009, 154, 453–465. [Google Scholar] [CrossRef]
- Gramazio, P.; Prohens, J.; Plazas, M.; Mangino, G.; Herraiz, F.J.; Vilanova, S. Development and genetic characterization of advanced backcross materials and an introgression line population of Solanum incanum in a S. melongena background. Front. Plant Sci. 2017, 8, 1477. [Google Scholar] [CrossRef]
- Adeniji, O.; Aloyce, A. Farmer’s knowledge of horticultural traits and participatory selection of African eggplant varieties (Solanum aethiopicum) in Tanzania. Tropicultura 2012, 30, 185–191. [Google Scholar]
- Datta, D.R.; Rafii, M.Y.; Misran, A.; Jusoh, M.; Yusuff, O.; Sulaiman, N.M.; Momodu, J. Genetic diversity, heritability and genetic advance of Solanum melongena L. from three secondary centers of diversity. Bangladesh J. Plant Taxonom. 2021, 28, 155–169. [Google Scholar] [CrossRef]
- Osei, M.; Oluoch, M.; Osei, C.; Banful, B. Morphological characterisafion of African Eggplant (Solanum spp.) Germplasm in some African countries. Agric. Innov. Sustain. Dev. 2010. Available online: https://library.faraafrica.org/wp-content/uploads/2019/11/Agricultural-Innovations-for-Sustainable-Development.pdf#page=171 (accessed on 15 July 2021).
- Plazas, M.; Vilanova, S.; Gramazio, P.; Rodríguez-Burruezo, A.; Fita, A.; Herraiz, F.J.; Ranil, R.; Fonseka, R.; Niran, L.; Fonseka, H. Interspecific hybridization between eggplant and wild relatives from different genepools. J. Am. Soc. Hortic. Sci. 2016, 141, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Polignano, G.; Uggenti, P.; Bisignano, V.; Della Gatta, C. Genetic divergence analysis in eggplant (Solanum melongena L.) and allied species. Genet. Resour. Crop. Evol. 2010, 57, 171–181. [Google Scholar] [CrossRef]
- Cericola, F.; Portis, E.; Toppino, L.; Barchi, L.; Acciarri, N.; Ciriaci, T.; Sala, T.; Rotino, G.L.; Lanteri, S. The population structure and diversity of eggplant from Asia and the Mediterranean Basin. PloS ONE 2013, 8, e73702. [Google Scholar]
- Syfert, M.M.; Castañeda-Álvarez, N.P.; Khoury, C.K.; Särkinen, T.; Sosa, C.C.; Achicanoy, H.A.; Bernau, V.; Prohens, J.; Daunay, M.C.; Knapp, S. Crop wild relatives of the brinjal eggplant (Solanum melongena): Poorly represented in genebanks and many species at risk of extinction. Am. J. Bot. 2016, 103, 635–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taher, D.; Solberg, S.Ø.; Prohens, J.; Chou, Y.-y.; Rakha, M.; Wu, T.-h. World vegetable center eggplant collection: Origin, composition, seed dissemination and utilization in breeding. Front. Plant Sci. 2017, 8, 1484. [Google Scholar] [CrossRef] [PubMed]
- Collonnier, C.; Fock, I.; Kashyap, V.; Rotino, G.; Daunay, M.; Lian, Y.; Mariska, I.; Rajam, M.; Servaes, A.; Ducreux, G. Applications of biotechnology in eggplant. Plant Cell Tissue Organ Cult. 2001, 65, 91–107. [Google Scholar] [CrossRef]
- Daunay, M.-C. Eggplant. In Vegetables II; Springer: Berlin/Heidelberg, Germany, 2008; pp. 163–220. [Google Scholar]
- Mutegi, E.; Snow, A.A.; Rajkumar, M.; Pasquet, R.; Ponniah, H.; Daunay, M.C.; Davidar, P. Genetic diversity and population structure of wild/weedy eggplant (Solanum insanum, Solanaceae) in southern India: Implications for conservation. Am. J. Bot. 2015, 102, 140–148. [Google Scholar] [CrossRef]
- Praneetha, K.J.D.S. Evaluation of brinjal (Solanum melongena L.) local types for yield and its quality characters. IJCS 2018, 6, 292–297. [Google Scholar]
- Rahman, M.; Islam, K.; Jahan, M.; Uddin, M. Efficacy of some botanicals in controlling brinjal shoot and fruit borer, Leucinodes orbonalis. Progress. Agric. 2009, 20, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Sharmin, D.; Meah, M.; Moniruzzaman, M. Inheritance of resistance to phomopsis blight and fruit rot in brinjal. J. Agrofor. Environ. 2010, 3, 135–140. [Google Scholar]
- Rocha, J.R.D.A.S.D.C.; Machado, J.C.; Carneiro, P.C.S. Multitrait index based on factor analysis and ideotype-design: Proposal and application on elephant grass breeding for bioenergy. Gcb Bioenergy 2018, 10, 52–60. [Google Scholar] [CrossRef]
- Sellami, M.H.; Lavini, A.; Pulvento, C. Phenotypic and Quality Traits of Chickpea Genotypes under Rainfed Conditions in South Italy. Agronomy 2021, 11, 962. [Google Scholar] [CrossRef]
- Olivoto, T.; Nardino, M. MGIDI: Toward an effective multivariate selection in biological experiments. Bioinformatics 2021, 37, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Verma, A.; Bisht, Y.S.; Maurya, P.; Jamir, I.; Mondal, S.; Bhattacharjee, T.; Chattopadhyay, A. Genetic variability, correlation coefficient and path coefficient analysis in brinjal germplasm. Int. J. Chem. Stud. 2018, 6, 3069–3073. [Google Scholar]
- Guide, F.F.R. Bangladesh Agricultural Research Council (BARC); BARC: Dhaka, Bangladesh, 2012.
- Withers, L.A.; Wheelans, S.K.; Williams, J.J.E. In vitro conservation of crop germplasm and the IBPGR databases. Euphytica 1990, 45, 9–22. [Google Scholar] [CrossRef]
- Reynolds, M.; Balota, M.; Delgado, M.; Amani, I.; Fischer, R. Physiological and morphological traits associated with spring wheat yield under hot, irrigated conditions. Funct. Plant Biol. 1994, 21, 717–730. [Google Scholar] [CrossRef]
- Radford, P. Growth analysis formulae-their use and abuse 1. Crop. Sci. 1967, 7, 171–175. [Google Scholar] [CrossRef]
- Rocha, J.R.D.A.S.D.C.; Nunes, K.V.; Carneiro, A.L.N.; Marçal, T.D.S.; Salvador, F.V.; Carneiro, P.C.S.; Carneiro, J.E.S. Selection of superior inbred progenies toward the common bean ideotype. Agron. J. 2019, 111, 1181–1189. [Google Scholar]
- Pinherio, J.; Bates, D.; DebRoy, S.; Deepayan, S.; Team, R.C. Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-145. 2007, Volume 3, pp. 1–89. Available online: https://ftp.uni-bayreuth.de/math/statlib/R/CRAN/doc/packages/nlme.pdf (accessed on 15 July 2021).
- Aravind, J.; Mukesh, S.; Wankhede, D.; Kaur, V. Augmented RCBD: Analysis of Augmented Randomised Complete Block Designs. R Package Version 0.1. 2020, Volume 2. Available online: https://aravind-j.github.io/augmentedRCBD/ (accessed on 15 July 2021).
- Falconer, D.S. Problems on Quantitative Genetics; Longman: London, UK, 1983. [Google Scholar]
- Burton, G.W. Quantitative inheritance in grasses. 6th Intl. Grassland Cong. Proc. 1952, 1, 227–238. [Google Scholar]
- Olivoto, T.; Nardino, M. MGIDI: A novel multi-trait index for genotype selection in plant breeding. Bioinformatics 2020, 1–22. [Google Scholar] [CrossRef]
- Wickham, H.; Chang, W.; Wickham, M.H. Package ‘ggplot2’. Create elegant data visualisations using the grammar of graphics. R Package Version 2016, 2, 1–189. [Google Scholar]
- Sarkar, M.; Kundagrami, S. Multivariate analysis in some genotypes of mungbean [Vigna radiata (L.) Wilczek] on the basis of agronomic traits of two consecutive growing cycles. Legume Res. Int. J. 2016, 39, 523–527. [Google Scholar] [CrossRef] [Green Version]
- Carrillo-Perdomo, E.; Vidal, A.; Kreplak, J.; Duborjal, H.; Leveugle, M.; Duarte, J.; Desmetz, C.; Deulvot, C.; Raffiot, B.; Marget, P. Development of new genetic resources for faba bean (Vicia faba L.) breeding through the discovery of gene-based SNP markers and the construction of a high-density consensus map. Sci. Rep. 2020, 10, 1–14. [Google Scholar]
- Islam, M.; Chhanda, R.; Pervin, N.; Hossain, M.; Chowdhury, R. Characterization and genetic diversity of brinjal germplasm. Bangladesh J. Agric. Res. 2018, 43, 499–512. [Google Scholar] [CrossRef]
- Begum, F.; Islam, A.A.; Rasul, M.G.; Mian, M.K.; Hossain, M.M. Morphological diversity of eggplant (Solanum melongena) in Bangladesh. Emir. J. Food Agric. 2013, 25, 45–51. [Google Scholar] [CrossRef]
- Solaiman, A.H.M.; Nishizawa, T.; Khatun, M.; Ahmad, S. Morphological characterization and genetic diversity studies of promising brinjal genotypes for hybridization program in Bangladesh. J. Adv. Agric. 2014, 3, 218–228. [Google Scholar] [CrossRef]
- Parida, H.; Mandal, J.; Mohanta, S. A note on morphological characterization of brinjal (Solanum melongena L.) genotypes. J. Crop. Weed 2020, 16, 250–255. [Google Scholar] [CrossRef]
- Shekar, K.C.; Ashok, P.; Sasikala, K. Characterization, character association, and path coefficient analyses in eggplant. Int. J. Veg. Sci. 2013, 19, 45–57. [Google Scholar] [CrossRef]
- Kumar, G.; Meena, B.; Kar, R.; Tiwari, S.K.; Gangopadhyay, K.; Bisht, I.; Mahajan, R. Morphological diversity in brinjal (Solanum melongena L.) germplasm accessions. Plant Genet. Resour. 2008, 6, 232–236. [Google Scholar] [CrossRef]
- Sunseri, F.; Polignano, G.B.; Alba, V.; Lotti, C.; Bisignano, V.; Mennella, G.; Drsquo, A.; Bacchi, M.; Riccardi, P.; Fiore, M.C.; et al. Genetic diversity and characterization of African eggplant germplasm collection. Afr. J. Plant Sci. 2010, 4, 231–241. [Google Scholar]
- Dash, S.P.; Singh, J.; Sharma, D. Morphological characterization of brinjal (Solanum melongena L.) germplasm. J. Pharmacogn. Phytochem. 2019, 8, 1574–1578. [Google Scholar]
- Tiwari, S.K.; Bisht, I.S.; Kumar, G.; Karihaloo, J.L. Diversity in brinjal (Solanum melongena L.) landraces for morphological traits of evolutionary significance. Veg. Sci. 2016, 43, 106–111. [Google Scholar]
- Solaimana, A.H.M.; Nishizawa, T.; Khatun, M.; Ahmad, S. Physio-morphological characterization genetic variability and correlation studies in brinjal genotypes of Bangladesh. Comput. Math. Biol. 2015, 4, 1–36. [Google Scholar]
- Begum, M.N.S.; Shirazy, B.J.; Mahbub, M.M.; Siddikee, M.A. Performance of Brinjal (Solanum Melongena) Genotypes through Genetic Variability Analysis. Am. J. Plant Biol. 2017, 43, 499–512. [Google Scholar]
- Shinde, K.; Birajdar, U.; Bhalekar, M.; Patil, B. Genetic divergence in brinjal (Solanum melongena L.). Veg. Sci. 2012, 39, 103–104. [Google Scholar]
- Singh, S.; Khemariya, P.; Rai, A.; Rai, A.C.; Koley, T.K.; Singh, B. Carnauba wax-based edible coating enhances shelf-life and retain quality of eggplant (Solanum melongena) fruits. LWT 2016, 74, 420–426. [Google Scholar] [CrossRef]
- Hassan, I.; Jatoi, S.A.; Arif, M.; Siddiqui, S.U. Genetic Variability in Eggplant for Agro-Morphological Traits; World Vegetable Center: Tainan, Taiwan, 2015. [Google Scholar]
- Premabati Devi, C.; Munshi, A.D.; Behera, T.K.; Choudhary, H.; Saha, P. Characterisation of cultivated breeding lines of eggplant (Solanum melongena L.) and related wild Solanum species from India. J. Hortic. Sci. Biotechnol. 2016, 91, 87–92. [Google Scholar] [CrossRef]
- Koundinya, A.; Das, A.; Layek, S.; Chowdhury, R.; Pandit, M.e. Genetic variability, characters association and path analysis for yield and fruit quality components in Brinjal. J. Appl. Nat. Sci. 2017, 9, 1343–1349. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.R.; Arumugam, T.; An, C.; Premalakshmi, V. Genetic variability for quantitative and qualitative characters in Brinjal (Solanum melongena L.). Afr. J. Agric. Res. 2013, 8, 4956–4959. [Google Scholar]
- Sherly, J.; Shanthi, A. Variability, heritability and genetic advance in brinjal (Solanum melongena L.). Res. Crop. 2009, 10, 105–108. [Google Scholar]
- Singh, O.; Kumar, J. Variability, heritability and genetic advance in brinjal. Indian J. Hortic. 2005, 62, 265–267. [Google Scholar]
- Verma, P.; Kushwaha, M.; Panchbhaiya, A. Studies on variability, heritability and genetic advance for yield attributing traits in brinjal (Solanum melongena L.) for two different seasons. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1543–1552. [Google Scholar] [CrossRef]
- Chattopadhyay, A.; Dutta, S.; Hazra, P. Characterization of genetic resources and identification of selection indices of brinjal (Solanum melongena L.) grown in eastern India. Veg. Crop. Res. Bull. 2011, 74, 39. [Google Scholar] [CrossRef] [Green Version]
- Shende, R.; Desai, S.; Lachyan, T. Genetic variability and response to selection in brinjal (Solanum melangena L.). Int. J. Curr. Res. 2015, 7, 21545–21547. [Google Scholar]
- Calus, M.P.; Veerkamp, R.F. Accuracy of multi-trait genomic selection using different methods. Genet. Sel. Evol. 2011, 43, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Konyak, W.; Kanaujia, S.; Jha, A.; Chaturvedi, H.; Ananda, A. Genetic variability, correlation and path coefficient analysis of brinjal. SAARC J. Agric. 2020, 18, 13–21. [Google Scholar] [CrossRef]
- Saha, S.; Haq, M.E.; Parveen, S.; Mahmud, F.; Chowdhury, S.R.; Harun-Ur-Rashid, M. Variability, correlation and path coefficient analysis: Principle tools to explore genotypes of brinjal (Solanum melongena L.). Asian J. Biotechnol. Genet. Eng. 2019, 2, 1–9. [Google Scholar]
- Dugard, P.; Todman, J.; Staines, H. Approaching Multivariate Analysis: A Practical Introduction; Routledge/Taylor & Francis Group: Abingdon, UK, 2010. [Google Scholar]
- Otieno, O.V.; Owuor, O.N. Multivariate genotype and genotype by environment interaction biplot analysis of sugarcane breeding data using R. Int. J. Stat. Distrib. Appl. 2019, 5, 22. [Google Scholar]
- Yan, W.; Frégeau-Reid, J. Breeding line selection based on multiple traits. Crop. Sci. 2008, 48, 417–423. [Google Scholar] [CrossRef]
- Khan, M.M.H.; Rafii, M.Y.; Ramlee, S.I.; Jusoh, M.; Al Mamun, M. Genetic analysis and selection of Bambara groundnut (Vigna Subterranea [L.] Verdc.) Landraces for high yield revealed by qualitative and quantitative traits. Sci. Rep. 2021, 11, 1–21. [Google Scholar]
- Meyer, R.S.; Bamshad, M.; Fuller, D.Q.; Litt, A. Comparing medicinal uses of eggplant and related Solanaceae in China, India, and the Philippines suggests the independent development of uses, cultural diffusion, and recent species substitutions. Econ. Bot. 2014, 68, 137–152. [Google Scholar] [CrossRef]
- Virga, G.; Licata, M.; Consentino, B.B.; Tuttolomondo, T.; Sabatino, L.; Leto, C.; La Bella, S. Agro-morphological characterization of sicilian chili pepper accessions for ornamental purposes. Plants 2020, 9, 1400. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, H.; Bhanu, A.; Srivastava, K.; Singh, M.; Shreya, H.A. Assessment of genetic diversity in crop plants-an overview. Adv. Plants Agric. Res. 2017, 7, 00255. [Google Scholar]
- Olivoto, T.; Diel, M.I.; Schmidt, D.; Lúcio, A.D.C. Multivariate analysis of strawberry experiments: Where are we now and where can we go? BioRxiv 2021, 1–10. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No. | Trait | Code | Sl. No. | Trait | Code |
|---|---|---|---|---|---|
| Qualitative Traits | Quantitative Traits | ||||
| 1. | Plant Growth Habit | PGH | 15. | Days to First Flowering | DFF (Day) |
| 2. | Leaf Blade Lobing | LBL | 16. | Plant Height | PH (cm) |
| 3. | Leaf Blade Tip Angle | LBTA | 17. | Fruit Diameter | FD (cm) |
| 4. | Leaf Prickles | LP | 18. | Fruit Length | FL (cm) |
| 5. | Leaf Hairs | LH | 19. | Fruit Weight | FW (g) |
| 6. | Corolla Color | CC | 20. | Normalized Difference Vegetation Index | NDVI |
| 7. | Fruit Calyx Prickles | FCP | 21. | Single Leaf Area | SLA (cm2) |
| 8. | Fruit Color Distribution | FCD | 22. | Soil Plant Analyses Development | SPAD |
| 9. | Fruit Curvature | FC | 23. | Total Number of Fruits | TF |
| 10. | Fruit Apex Shape | FAS | 24. | Yield Per Plant | YPP (kg) |
| 11. | Fruit Cross Section | FCS | |||
| 12. | Fruit Color at Ripening | FCR | |||
| 13. | Fruit Flesh Density | FFD | |||
| 14. | Fruit Position | FP | |||
| Trait Names | Descriptor State | No. of Germplasm | Germplasm (%) |
|---|---|---|---|
| Plant Growth Habit | Upright | 23 | 17.69 |
| Intermediate | 64 | 49.23 | |
| Prostrate | 43 | 33.08 | |
| Leaf Blade Lobing | Weak | 59 | 45.38 |
| Intermediate | 43 | 33.08 | |
| Strong | 23 | 17.69 | |
| Very strong | 5 | 3.85 | |
| Leaf Blade Tip Angle | Very acute | 6 | 4.62 |
| Acute | 28 | 21.54 | |
| Intermediate | 64 | 49.23 | |
| Obtuse | 32 | 24.62 | |
| Leaf Prickles | Very few | 13 | 10.00 |
| Few | 38 | 29.23 | |
| Intermediate | 72 | 55.38 | |
| Many | 5 | 3.85 | |
| Very many | 2 | 1.54 | |
| Leaf Hairs | Very many | 128 | 98.46 |
| Few | 2 | 1.54 | |
| CorollaColor | Light Violet | 41 | 31.54 |
| Pale Violet | 84 | 64.62 | |
| White | 5 | 3.85 |
| Descriptor Name | Descriptor State | No. of Germplasm | Germplasm (%) |
|---|---|---|---|
| Fruit Calyx Prickles | Very few | 3 | 2.31 |
| Few | 31 | 23.85 | |
| Intermediate | 49 | 37.69 | |
| Many | 32 | 24.62 | |
| Very many | 15 | 11.54 | |
| Fruit Color Distribution | Uniform | 18 | 13.85 |
| Mottled | 21 | 16.15 | |
| Netted | 18 | 13.85 | |
| Striped | 73 | 56.15 | |
| Fruit Curvature | None (fruit straight) | 68 | 52.31 |
| Slightly curved | 37 | 28.46 | |
| Curved | 25 | 19.23 | |
| Fruit Apex Shape | Rounded | 39 | 30.00 |
| Depressed | 91 | 70.00 | |
| Fruit Cross Section | Circular (no grooves) | 130 | 100.00 |
| Fruit Color at Ripening | Milky white | 2 | 1.54 |
| Lilac grey | 7 | 5.38 | |
| Purple | 32 | 24.62 | |
| Green with mottled at the distal end | 51 | 39.23 | |
| Green with a yellowish stripe | 37 | 28.46 | |
| Purple with light green at the distal end | 3 | 2.31 | |
| Fruit Flesh Density | Very loose (spongy) | 74 | 56.92 |
| Loose (crumbly) | 24 | 18.46 | |
| Average density | 21 | 16.15 | |
| Dense | 7 | 5.38 | |
| Very dense | 4 | 3.08 | |
| Fruit Position | Pendant | 130 | 100.00 |
| Traits | Source of Variation | |||||
|---|---|---|---|---|---|---|
| Accession (G) with C (df = 129) | Check (C) (df = 3) | Accession (G) vs. C (df = 1) | Accession (G) (df = 125) | Adjusted Block (B) (df = 6) | Residuals (df = 18) | |
| DFF | 59.61 ** | 161.62 ** | 0.7 ns | 57.63 ** | 20.31 ns | 16.67 |
| FD | 390.72 ** | 586.56 ** | 766.74 ** | 383.01 ** | 31.44 ns | 27.71 |
| FL | 3.91 ** | 5.87 ** | 7.67 ** | 3.83 ** | 0.31 ns | 0.28 |
| FW | 8.63 ** | 49.04 ** | 0.36 ns | 7.72 ** | 1.67 ns | 1.27 |
| NDVI | 0.01 ** | 0.01 ns | 0.07 ** | 0.01 ** | 0.0023 ns | 0.0028 |
| PH | 169.94 ** | 18.57 ns | 4014.75 ** | 142.81 ** | 3.14 ns | 7.57 |
| SLA | 1572.19 ** | 71.58 ns | 40431.19 ** | 1297.33 ** | 8.38 ns | 22.74 |
| SPAD | 59.63 ** | 235.78 ** | 637.78 ** | 50.78 ** | 17.9 ns | 9.96 |
| TF | 104.55 ** | 128.42 ** | 68.89 ** | 104.26 ** | 2.54 ns | 4.25 |
| YPP | 9.59 ** | 0.9 ns | 3.89 ns | 9.84 ** | 0.97 ns | 1.12 |
| Trait | Mean | Std.Error | CV | Min | Max | Skewness | Kurtosis |
|---|---|---|---|---|---|---|---|
| PH | 111.4 | 1.05 | 2.52 | 87.82 | 139.57 | 0.21 ns | 2.25 * |
| NDVI | 0.56 | 0.01 | 9.55 | 0.33 | 0.78 | 0.11 ns | 2.7 ns |
| SPAD | 47.5 | 0.66 | 6.54 | 25.37 | 66.97 | −0.07 ns | 3.37 ns |
| SLA | 140.53 | 3.18 | 3.55 | 86.12 | 242.85 | 0.56 ** | 2.37 ns |
| DFF | 98.4 | 0.7 | 4.15 | 83.32 | 122.07 | 0.46 * | 3.27 ns |
| FL | 47.25 | 1.6 | 11.35 | 3.71 | 24.73 | 0.47 * | 2.71 ns |
| FD | 4.72 | 0.16 | 11.35 | 0.77 | 9.47 | 0.47 * | 2.71 ns |
| FW | 7.55 | 0.23 | 39.52 | 10.00 | 650.40 | 0.14 ns | 2.69 ns |
| TF | 18.65 | 0.86 | 16.27 | 10.21 | 49.21 | 2.25 ** | 7.16 ** |
| YPP | 7.12 | 0.28 | 46.14 | 0.24 | 10.57 | 0.54 * | 5.41 ** |
| Traits | PV | GV | GCV | GCV | PCV | PCV | hBS | hBS | GA | GAM | GAM |
|---|---|---|---|---|---|---|---|---|---|---|---|
| PH | 142.81 | 135.24 | 10.44 | Medium | 10.73 | Medium | 94.7 | High | 23.35 | 20.96 | High |
| NDVI | 0.01 | 0.0047 | 12.26 | Medium | 15.46 | Medium | 62.92 | High | 0.11 | 20.07 | High |
| SPAD | 50.78 | 40.82 | 13.45 | Medium | 15 | Medium | 80.38 | High | 11.82 | 24.88 | High |
| SLA | 1297.3 | 1274.6 | 25.4 | High | 25.63 | High | 98.25 | High | 73 | 51.95 | High |
| DFF | 57.63 | 40.96 | 6.5 | Low | 7.71 | Low | 71.07 | High | 11.13 | 11.31 | Medium |
| FD | 383.01 | 355.3 | 39.9 | High | 41.42 | High | 92.77 | High | 37.45 | 79.27 | High |
| FL | 3.83 | 3.55 | 39.9 | High | 41.42 | High | 92.77 | High | 3.75 | 79.27 | High |
| FW | 7.72 | 6.46 | 33.65 | High | 36.8 | High | 83.61 | High | 4.79 | 63.47 | High |
| TF | 104.26 | 100.01 | 53.64 | High | 54.76 | High | 95.92 | High | 20.21 | 108.37 | High |
| YPP | 9.84 | 8.72 | 41.49 | High | 44.07 | High | 88.63 | High | 5.74 | 80.58 | High |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uddin, M.S.; Billah, M.; Afroz, R.; Rahman, S.; Jahan, N.; Hossain, M.G.; Bagum, S.A.; Uddin, M.S.; Khaldun, A.B.M.; Azam, M.G.; et al. Evaluation of 130 Eggplant (Solanum melongena L.) Genotypes for Future Breeding Program Based on Qualitative and Quantitative Traits, and Various Genetic Parameters. Horticulturae 2021, 7, 376. https://doi.org/10.3390/horticulturae7100376
Uddin MS, Billah M, Afroz R, Rahman S, Jahan N, Hossain MG, Bagum SA, Uddin MS, Khaldun ABM, Azam MG, et al. Evaluation of 130 Eggplant (Solanum melongena L.) Genotypes for Future Breeding Program Based on Qualitative and Quantitative Traits, and Various Genetic Parameters. Horticulturae. 2021; 7(10):376. https://doi.org/10.3390/horticulturae7100376
Chicago/Turabian StyleUddin, Md. Shalim, Masum Billah, Rozina Afroz, Sajia Rahman, Nasrin Jahan, Md. Golam Hossain, Shamim Ara Bagum, Md. Sorof Uddin, Abul Bashar Mohammad Khaldun, Md. Golam Azam, and et al. 2021. "Evaluation of 130 Eggplant (Solanum melongena L.) Genotypes for Future Breeding Program Based on Qualitative and Quantitative Traits, and Various Genetic Parameters" Horticulturae 7, no. 10: 376. https://doi.org/10.3390/horticulturae7100376
APA StyleUddin, M. S., Billah, M., Afroz, R., Rahman, S., Jahan, N., Hossain, M. G., Bagum, S. A., Uddin, M. S., Khaldun, A. B. M., Azam, M. G., Hossain, N., Akanda, M. A. L., Alhomrani, M., Gaber, A., & Hossain, A. (2021). Evaluation of 130 Eggplant (Solanum melongena L.) Genotypes for Future Breeding Program Based on Qualitative and Quantitative Traits, and Various Genetic Parameters. Horticulturae, 7(10), 376. https://doi.org/10.3390/horticulturae7100376