Comparison of Secondary Metabolite Contents and Metabolic Profiles of Six Lycoris Species

 ,
, 
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials, Experimental Design, and Cultural Management
2.2. HPLC Analysis and Extraction of Galantamine
2.3. GC-TOFMS Analysis
2.4. Statistical Analysis
3. Results
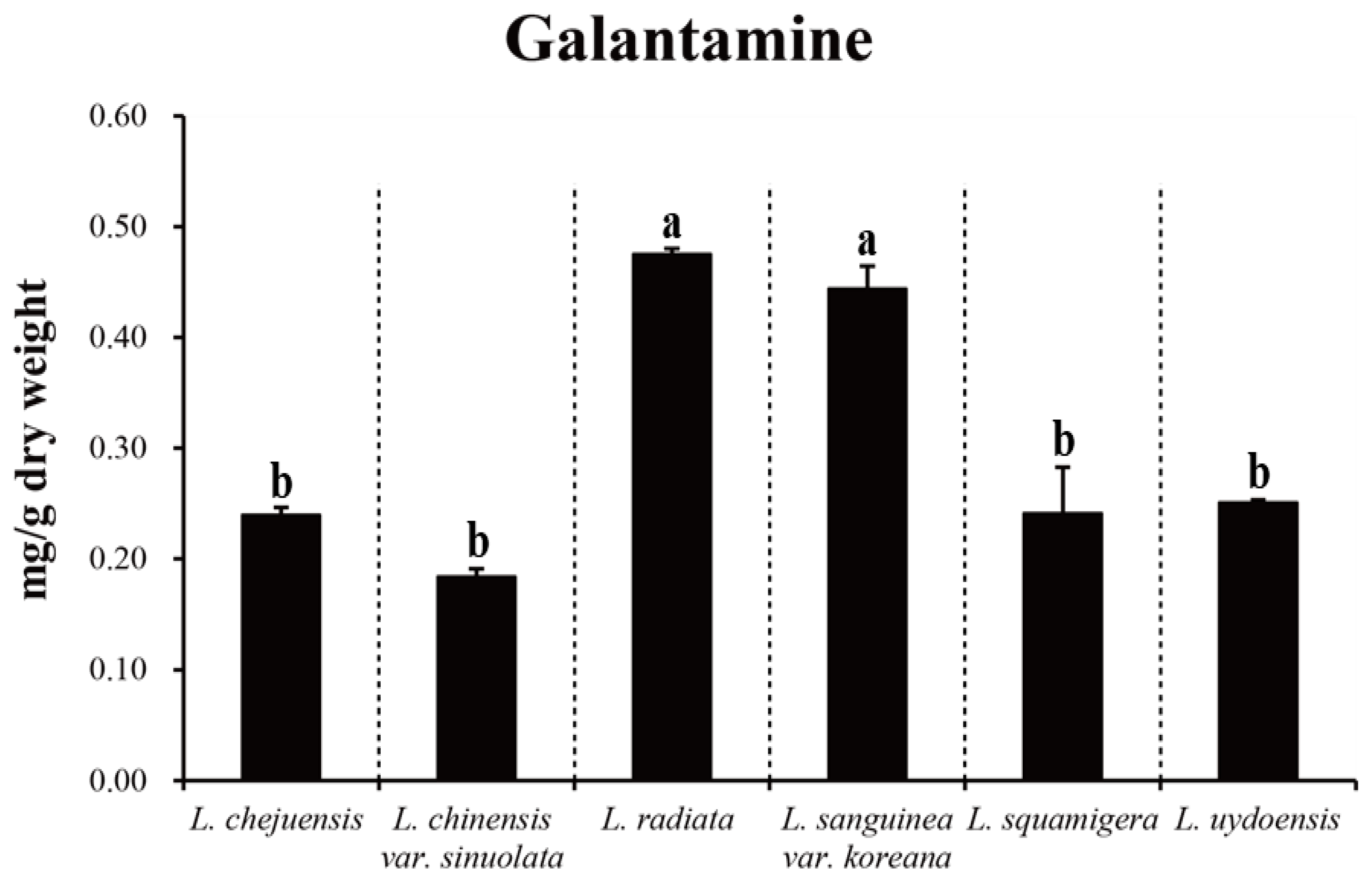
3.1. Subsection Variation of Galantamine Content Among the Lycoris spp.
3.2. Phenylpropanoid Content of Six Lycoris spp.
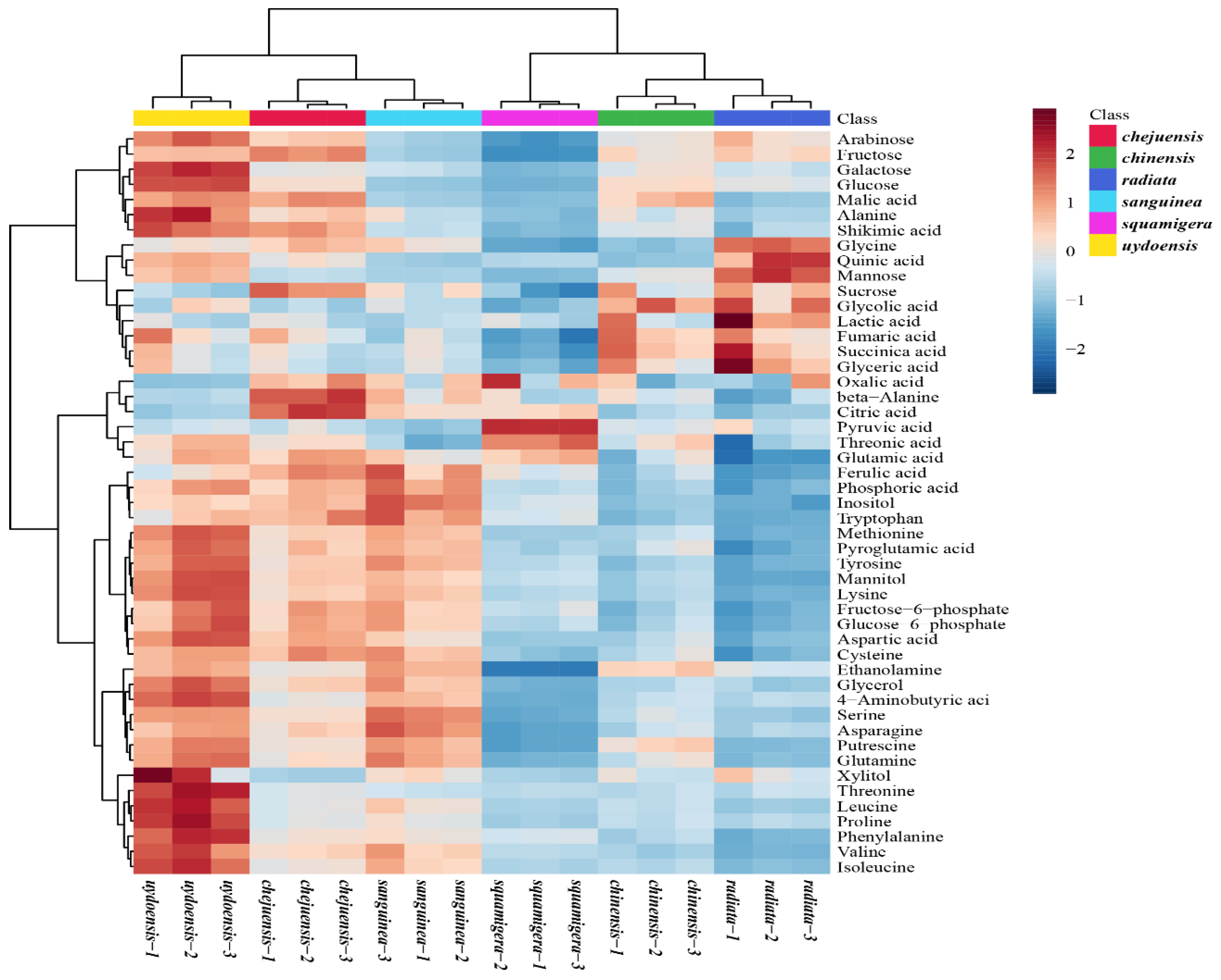
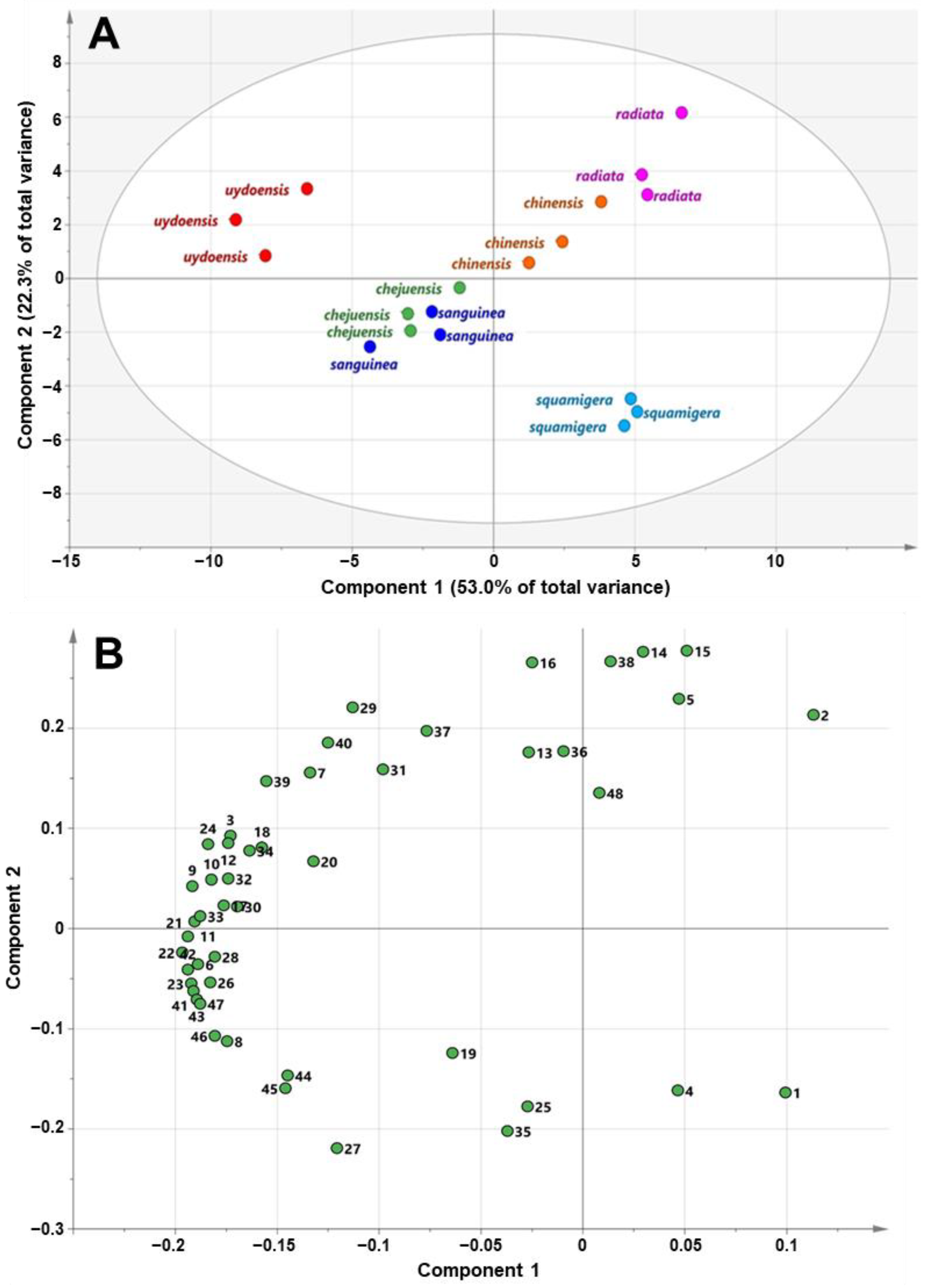
3.3. GC-TOFMS Metabolic Profiling of Six Lycoris spp.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jimenez, H.J.; Silva, A.D.F.; Martins, L.S.S.; Carvalho, R.D.; Moraes, F.R.M. Comparative genomics plastomes of the Amaryllidaceae family species. Scientia Plena 2020, 16, 1–11. [Google Scholar]
- Hayashi, A.; Saito, T.; Mukai, Y.; Kurita, S.; Hori, T.-A. Genetic variations in Lycoris radiata var. radiata in Japan. Genes Genet. Syst. 2005, 80, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Han, Y.; Park, K. Vegetation present and vascular plants of habitats Lycoris flavescens MY Kim et ST Lee and Iris koreana Nakai, Byeonsanbando. J. Agric. Life Sci. 2011, 42, 54–67. [Google Scholar]
- Lee, D.G.; Lee, A.Y.; Kim, S.-J.; Lee, S.; Cho, E.; Lee, S. Antibacterial phytosterols and alkaloids from Lycoris radiata. Nat. Prod. Sci. 2014, 20, 107–112. [Google Scholar]
- Arias, E.; Alés, E.; Gabilan, N.H.; Cano-Abad, M.F.; Villarroya, M.; García, A.G.; López, M.G. Galantamine prevents apoptosis induced by β-amyloid and thapsigargin: Involvement of nicotinic acetylcholine receptors. Neuropharmacology 2004, 46, 103–114. [Google Scholar] [CrossRef]
- Chang, K.L.; Ho, P.C. Gas chromatography time-of-flight mass spectrometry (GC-TOF-MS)-based metabolomics for comparison of caffeinated and decaffeinated coffee and its implications for Alzheimer’s disease. PLoS ONE 2014, 9, e104621. [Google Scholar] [CrossRef]
- Li, A.; Du, Z.; Liao, M.; Feng, Y.; Ruan, H.; Jiang, H. Discovery and characterisation of lycorine-type alkaloids in Lycoris spp. (Amaryllidaceae) using UHPLC-QTOF-MS. Phytochem Anal. 2019, 30, 268–277. [Google Scholar] [CrossRef]
- Bastida, A.J.; Berkov, S.; Torras, C.L.; Pigni, N.B.; Andrade, J.P.D.; Martínez, V.; Codina, M.C.; Meya, F. Chemical and biological aspects of Amaryllidaceae alkaloids. Recent Adv. Pharm. Sci. 2011, 3, 65–100. [Google Scholar]
- Wang, D.; Gao, C.; Liu, S.; Wei, L. Structure investigation of a new alkaloid from Zanthoxylum schinifolium Siebet zucc. Chem. Res. Chin. Univ. 1991, 7, 124–128. [Google Scholar]
- Liu, J.; Hu, W.; He, L.; Ye, M.; Li, Y. Effects of lycorine on HL-60 cells via arresting cell cycle and inducing apoptosis. FEBS Lett. 2004, 578, 245–250. [Google Scholar] [CrossRef]
- Jin, Z. Amaryllidaceae and Sceletium alkaloids. Nat. Prod. Rep. 2007, 24, 886–905. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xia, N.; Li, X.; Shen, W.; Liang, L.; Wang, C.; Wang, R.; Peng, F.; Xia, B. Molecular cloning and characterization of a phenylalanine ammonia-lyase gene (LrPAL) from Lycoris radiata. Mol. Biol. Rep. 2011, 38, 1935–1940. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Pan, D.-S.; Han, S.; Yu, C.-Y.; Zhao, Q.-J.; Song, Y.; Liang, Y. Alkaloids from Lycoris caldwellii and their particular cytotoxicities against the astrocytoma and glioma cell lines. Arch. Pharmacal Res. 2013, 36, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.; Shen, S.-F.; Zhao, Q.-J. Cytotoxic and antimalarial amaryllidaceae alkaloids from the bulbs of Lycoris radiata. Molecules 2013, 18, 2458–2468. [Google Scholar] [CrossRef]
- Song, J.-H.; Zhang, L.; Song, Y. Alkaloids from Lycoris aurea and their cytotoxicities against the head and neck squamous cell carcinoma. Fitoterapia 2014, 95, 121–126. [Google Scholar] [CrossRef]
- Atanasova, M.; Stavrakov, G.; Philipova, I.; Zheleva, D.; Yordanov, N.; Doytchinova, I.A. Galantamine derivatives with indole moiety: Docking, design, synthesis and acetylcholinesterase inhibitory activity. Bioorg. Med. Chem. 2015, 23, 5382–5389. [Google Scholar] [CrossRef]
- Suffness, M.; Cordell, G.A. Antitumor alkaloids. In The Alkaloids: Chemistry and Pharmacology; Brossi, A., Ed.; Academic Press INC: Orlando, FL, USA, 1985; Volume 25, pp. 1–355. [Google Scholar]
- Kihara, M.; Konishi, K.; Xu, L.; Kobayashi, S. Alkaloidal constituents of the flowers of Lycoris radiata herb amaryllidaceae. Chem. Pharm. Bull. 1991, 39, 1849–1853. [Google Scholar] [CrossRef]
- Park, C.H.; Morgan, A.M.; Park, B.B.; Lee, S.Y.; Lee, S.; Kim, J.K.; Park, S. Metabolic analysis of four cultivars of Liriope platyphylla. Metabolites 2019, 9, 59. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Baskar, T.B.; Park, Y.E.; Park, J.; Lee, S.Y.; Park, S. In vitro antioxidant and antimicrobial properties of flower, leaf, and stem extracts of Korean mint. Antioxidants 2019, 8, 75. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Kim, J.B.; Eun, P.Y.; Kim, S.-J.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.-Y.; Kim, J.K.; Park, S. Metabolic profiling of pale green and purple kohlrabi (Brassica oleracea var. gongylodes). Appl. Biol. Chem. 2017, 60, 249–257. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Park, S.-Y.; Kim, J.K.; Park, S. Comparative phytochemical analyses and metabolic profiling of different phenotypes of Chinese cabbage (Brassica Rapa ssp. Pekinensis). Foods 2019, 8, 587. [Google Scholar] [CrossRef] [PubMed]
- Quan, M.; Ou, L.; She, C.; Wu, X.; Chen, D. rDNA internal transcribed spacer sequence analysis of Lycoris Hert. Afr. J. Biotechnol. 2012, 11, 29. [Google Scholar]
- Park, C.H.; Yeo, H.J.; Park, Y.E.; Baek, S.-A.; Kim, J.K.; Park, S. Transcriptome analysis and metabolic profiling of Lycoris radiata. Biology 2019, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Yue, Y.; Shi, M.; Chen, M.; Yang, X.; Wang, L. Exploration of floral volatile organic compounds in six typical Lycoris taxa by GC-MS. Plants 2019, 8, 422. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.B.; Kim, Y.J.; Kim, H.J.; Choi, J.; Kim, J.K. Phytochemical profiles of Brassicaceae vegetables and their multivariate characterization using chemometrics. Appl. Biol. Chem. 2018, 61, 131–144. [Google Scholar] [CrossRef]
- Kim, T.J.; Hyeon, H.; Park, N.-I.; Yi, T.G.; Lim, S.-H.; Park, S.-Y.; Ha, S.-H.; Kim, J.K. A high-throughput platform for interpretation of metabolite profile data from pepper (Capsicum) fruits of 13 phenotypes associated with different fruit maturity states. Food Chem. 2020, 331, 127286. [Google Scholar] [CrossRef]
- De Paiva, J.; Souza, A.; Pereira, R.; Ribeiro, P.; Zocolo, G.J.; De Brito, E.S.; Pessoa, O.D.L.; Canuto, K.M. Development and validation of a UPLC-ESI-MS method for quantitation of the anti-alzheimer drug galantamine and other amaryllidaceae alkaloids in plants. J. Braz. Chem. Soc. 2020, 31, 265–272. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, C.-Y.; Guo, M. Comparative analysis of Amaryllidaceae alkaloids from three Lycoris species. Molecules 2015, 20, 21854–21869. [Google Scholar] [CrossRef]
- Janssen, B.; Schäfer, B. Galantamine. Chemtexts 2017, 3, 7. [Google Scholar] [CrossRef]
- Nikolova, M.; Gevrenova, R. Determination of phenolic acids in Amaryllidaceae species by high performance liquid chromatography. Pharm. Biol. 2005, 43, 289–291. [Google Scholar] [CrossRef]
- Park, C.H.; Park, S.-Y.; Lee, S.Y.; Kim, J.K.; Park, S. Analysis of metabolites in white flowers of Magnolia Denudata Desr. and violet flowers of Magnolia Liliiflora Desr. Molecules 2018, 23, 1558. [Google Scholar] [CrossRef] [PubMed]
- Francisco, M.; Velasco, P.; Moreno, D.A.; García-Viguera, C.; Cartea, M.E. Cooking methods of Brassica rapa affect the preservation of glucosinolates, phenolics and vitamin C. Food Res. Int. 2010, 43, 1455–1463. [Google Scholar] [CrossRef]
- Park, C.H.; Bong, S.J.; Lim, C.J.; Kim, J.K.; Park, S.U. Transcriptome analysis and metabolic profiling of green and red mizuna (Brassica rapa L. var. japonica). Foods 2020, 9, 1079. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.R.; Li, X.; Park, W.T.; Kim, Y.B.; Kim, S.J.; Kim, Y.S.; Lee, M.Y.; Park, C.H.; Park, S.U. Phenolic compound content in different organs of Korean common buckwheat cultivars. Asian J. Chem. 2013, 25, 424. [Google Scholar] [CrossRef]
- Kim, Y.K.; Park, W.T.; Uddin, M.R.; Kim, Y.B.; Bae, H.; Kim, H.H.; Park, K.W.; Park, S.U. Variation of charantin content in different bitter melon cultivars. Asian J. Chem. 2014, 26, 309. [Google Scholar] [CrossRef]
- Park, W.T.; Kim, J.K.; Park, S.; Lee, S.-W.; Li, X.; Kim, Y.B.; Uddin, M.R.; Park, N.I.; Kim, S.-J.; Park, S.U. Metabolic profiling of glucosinolates, anthocyanins, carotenoids, and other secondary metabolites in kohlrabi (Brassica oleracea var. gongylodes). J. Agric. Food Chem. 2012, 60, 8111–8116. [Google Scholar] [CrossRef]
- Kim, Y.K.; Li, X.; Xu, H.; Park, N.I.; Uddin, M.R.; Pyon, J.Y.; Park, S.U. Production of phenolic compounds in hairy root culture of Tartary buckwheat (Fagopyrum tataricum Gaertn). J. Crop Sci. Biotechnol. 2009, 12, 53–57. [Google Scholar] [CrossRef]
- Thwe, A.A.; Kim, J.K.; Li, X.; Kim, Y.B.; Uddin, M.R.; Kim, S.J.; Suzuki, T.; Park, N.I.; Park, S.U. Metabolomic analysis and phenylpropanoid biosynthesis in hairy root culture of Tartary buckwheat cultivars. PLoS ONE 2013, 8, e65349. [Google Scholar] [CrossRef]
- Park, N.I.; Xiaohua, L.; Uddin, R.M.; Park, S.U. Phenolic compound production by different morphological phenotypes in hairy root cultures of Fagopyrum tataricum Gaertn. Arch. Biol. Sci. 2011, 63, 193–198. [Google Scholar] [CrossRef]
- Colque, R.; Viladomat, F.; Bastida, J.; Codina, C. Improved production of galanthamine and related alkaloids by methyl jasmonate in Narcissus confusus shoot-clumps. Planta Medica 2004, 70, 1180–1188. [Google Scholar] [CrossRef]
- Hu, X.; Neill, S.J.; Cai, W.; Tang, Z. Nitric oxide mediates elicitor-induced saponin synthesis in cell cultures of Panax ginseng. Funct. Plant Biol. 2003, 30, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.P.; Guo, Y.T.; Wang, J.W.; Tan, R.X. Nitric oxide potentiates oligosaccharide-induced artemisinin production in Artemisia annua hairy roots. J. Integr. Plant Biol. 2008, 50, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.-M.; Wang, R.; Li, X.-D.; Jiang, Y.-M.; Wang, C.-Y.; Quan, J.-P.; Peng, F.; Xia, B. Effect of abiotic and biotic elicitors on growth and alkaloid accumulation of Lycoris chinensis seedlings. Zeitschrift Naturforschung C 2009, 64, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Kim, Y.S.; Kim, Y.; Uddin, M.R.; Kim, Y.B.; Kim, H.H.; Park, S.Y.; Lee, M.Y.; Chung, S.O.; Park, S.U. Comparative analysis of flavonoids and polar metabolites from hairy roots of Scutellaria baicalensis and Scutellaria lateriflora. World J. Microbiol. Biotechnol. 2014, 30, 887–892. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| L. uydoensis | L. squamigera | L. sanguinea | L. radiata | L. chinensis | L. chejuensis | Compounds |
|---|---|---|---|---|---|---|
| 0.058 ± 0.004 b | 0.063 ± 0.005 a | 0.047 ± 0.004 c | 0.010 ± 0.000 d | 0.011 ± 0.000 d | 0.009 ± 0.000 d z | Gallic acid |
| 0.315 ± 0.024 a | 0.346 ± 0.025 a | 0.348 ± 0.024 a | 0.053 ± 0.002 c | 0.135 ± 0.003 b | 0.125 ± 0.003 b | Catechin |
| ND | ND | 0.020 ± 0.001 a | 0.002 ± 0.000 b | <0.000 b | ND y | 4-hydroxybenzoic acid |
| 0.304 ± 0.024 a | 0.289 ± 0.015 a | 0.287 ± 0.015 a | 0.055 ± 0.001 b | 0.064 ± 0.000 b | 0.048 ± 0.000 b | Chlorogenic acid |
| 0.093 ± 0.006 a | 0.093 ± 0.006 a | 0.093 ± 0.006 a | 0.016 ± 0.000 b | 0.022 ± 0.000 b | 0.017 ± 0.000 b | Caffeic acid |
| 0.071 ± 0.001 b | 0.073 ± 0.001 b | 0.100 ± 0.001 a | 0.010 ± 0.000 e | 0.019 ± 0.002 c | 0.013 ± 0.001 d | (−)-Epicatechin |
| 0.315 ± 0.024 a | 0.279 ± 0.015 b | 0.278 ± 0.015 b | 0.050 ± 0.001 c | 0.066 ± 0.000 c | 0.049 ± 0.000 c | Epicatechin gallate |
| ND | ND | ND | 0.002 ± 0.000 a | ND | <0.000 b | Sinapic acid |
| 0.282 ± 0.015 a | 0.284 ± 0.015 a | 0.282 ± 0.015 a | 0.051 ± 0.000 b | 0.067 ± 0.001 b | 0.051 ± 0.001 b | benzoic acid |
| 0.821 ± 0.051 a | 0.814 ± 0.051 a | 0.832 ± 0.051 a | 0.159 ± 0.002 b | 0.187 ± 0.001 b | 0.148 ± 0.001 b | Rutin |
| 0.602 ± 0.036 a | 0.605 ± 0.048 a | 0.605 ± 0.036 a | 0.114 ± 0.009 b | 0.143 ± 0.001 b | 0.108 ± 0.001 b | Quercetin |
| 0.215 ± 0.013 a | 0.217 ± 0.013 a | 0.218 ± 0.013 a | 0.039 ± 0.002 b | 0.051 ± 0.000 b | 0.038 ± 0.000 b | Kaempferol |
| 3.079 ± 0.208 a | 3.062 ± 0.205 a | 3.109 ± 0.209 a | 0.561 ± 0.010 b | 0.765 ± 0.004 b | 0.607 ± 0.001 b | Total |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeo, H.J.; Kim, Y.J.; Nguyen, B.V.; Park, Y.E.; Park, C.H.; Kim, H.H.; Kim, J.K.; Park, S.U. Comparison of Secondary Metabolite Contents and Metabolic Profiles of Six Lycoris Species. Horticulturae 2021, 7, 5. https://doi.org/10.3390/horticulturae7010005
Yeo HJ, Kim YJ, Nguyen BV, Park YE, Park CH, Kim HH, Kim JK, Park SU. Comparison of Secondary Metabolite Contents and Metabolic Profiles of Six Lycoris Species. Horticulturae. 2021; 7(1):5. https://doi.org/10.3390/horticulturae7010005
Chicago/Turabian StyleYeo, Hyeon Ji, Ye Jin Kim, Bao Van Nguyen, Ye Eun Park, Chang Ha Park, Haeng Hoon Kim, Jae Kwang Kim, and Sang Un Park. 2021. "Comparison of Secondary Metabolite Contents and Metabolic Profiles of Six Lycoris Species" Horticulturae 7, no. 1: 5. https://doi.org/10.3390/horticulturae7010005
APA StyleYeo, H. J., Kim, Y. J., Nguyen, B. V., Park, Y. E., Park, C. H., Kim, H. H., Kim, J. K., & Park, S. U. (2021). Comparison of Secondary Metabolite Contents and Metabolic Profiles of Six Lycoris Species. Horticulturae, 7(1), 5. https://doi.org/10.3390/horticulturae7010005