Freezing Tolerance and Chilling Fulfillment Differences in Cold Climate Grape Cultivars




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Low Temperature Exotherms
2.3. Dormancy Status
2.4. Statistical Analysis
2.4.1. Freezing
2.4.2. Dormancy Status
3. Results
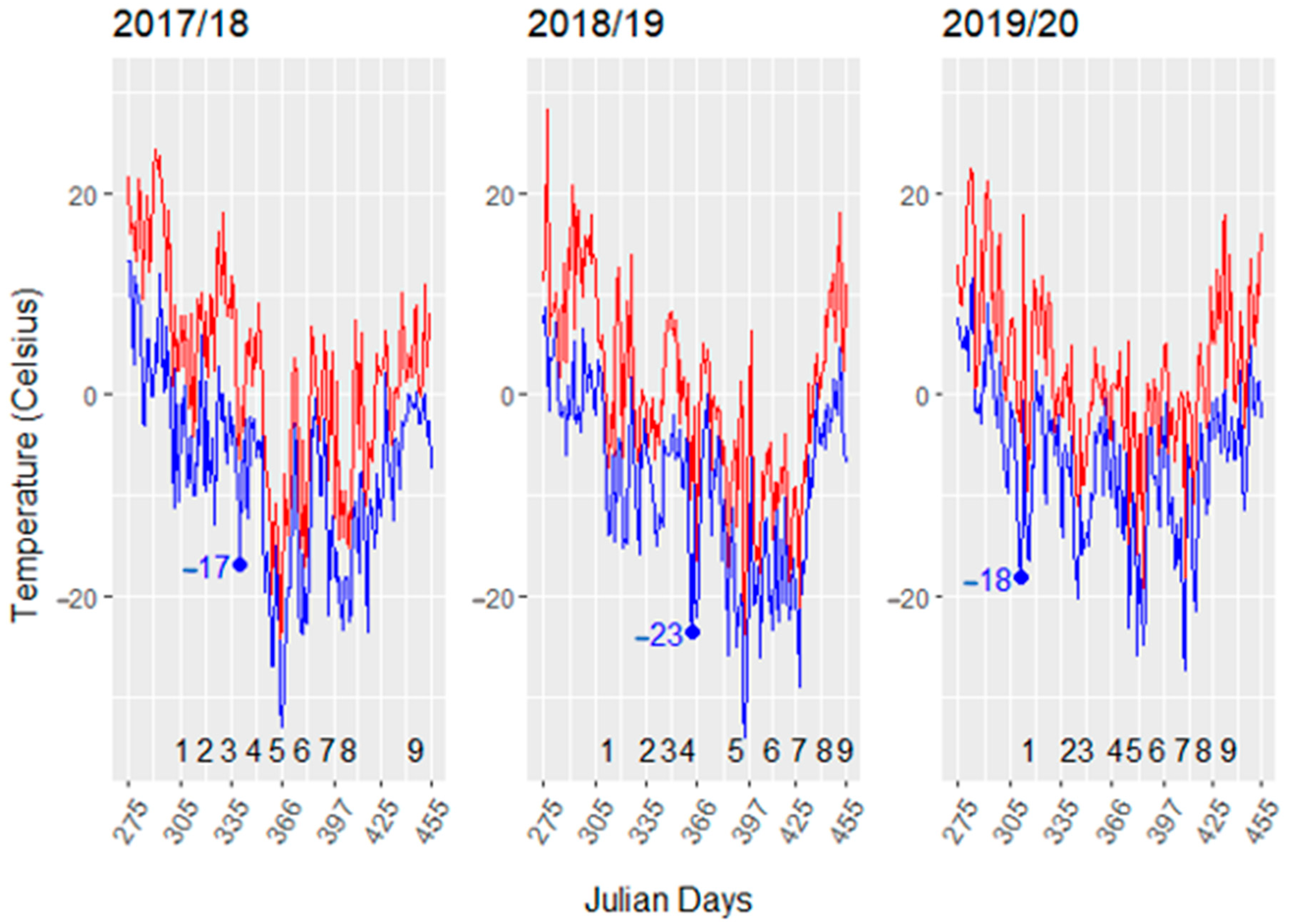
3.1. Dormant Season Temperature Variation 2017–2020
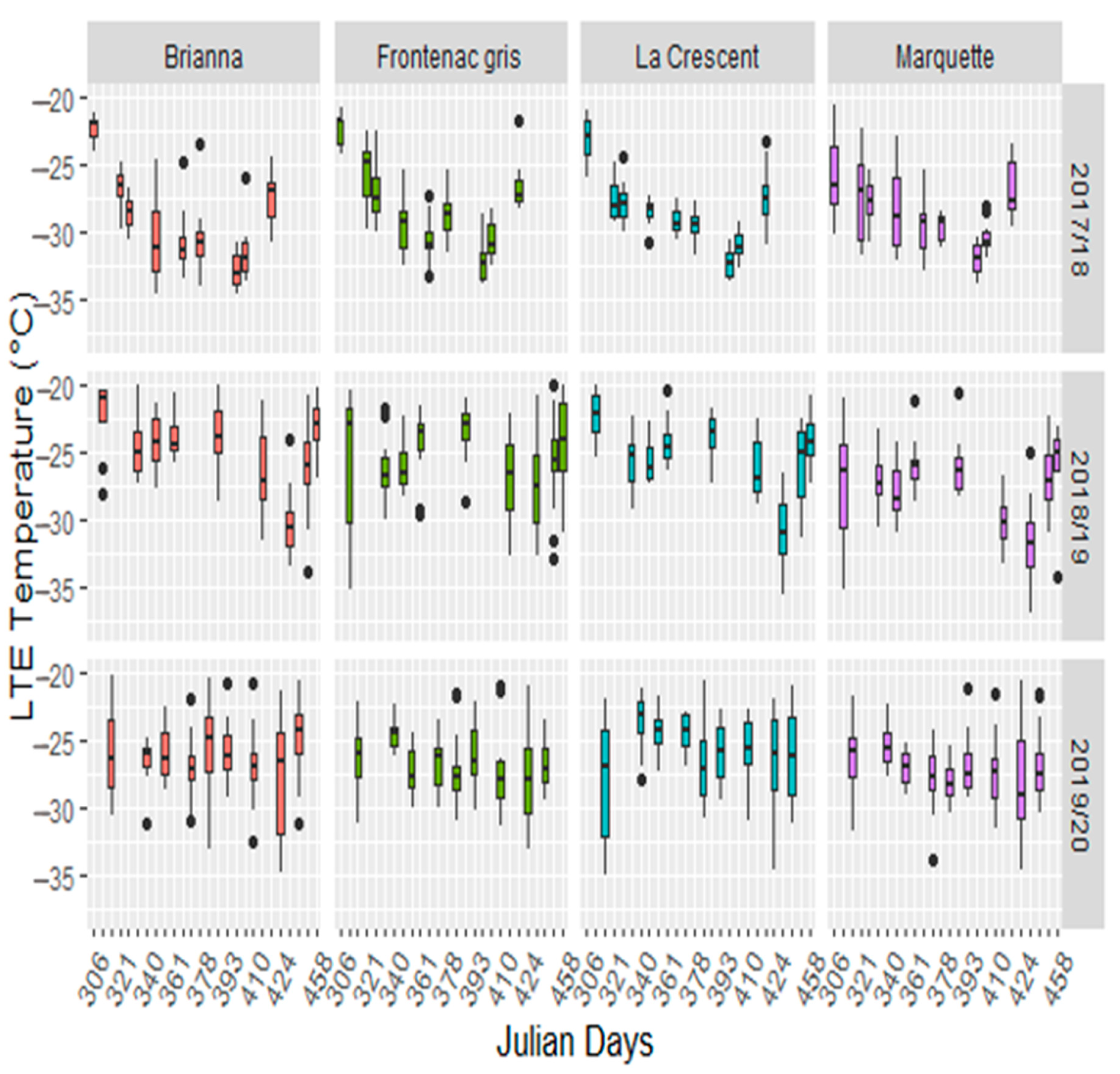
3.2. Bud Freezing Tolerance Differs between Seasons and Cultivars
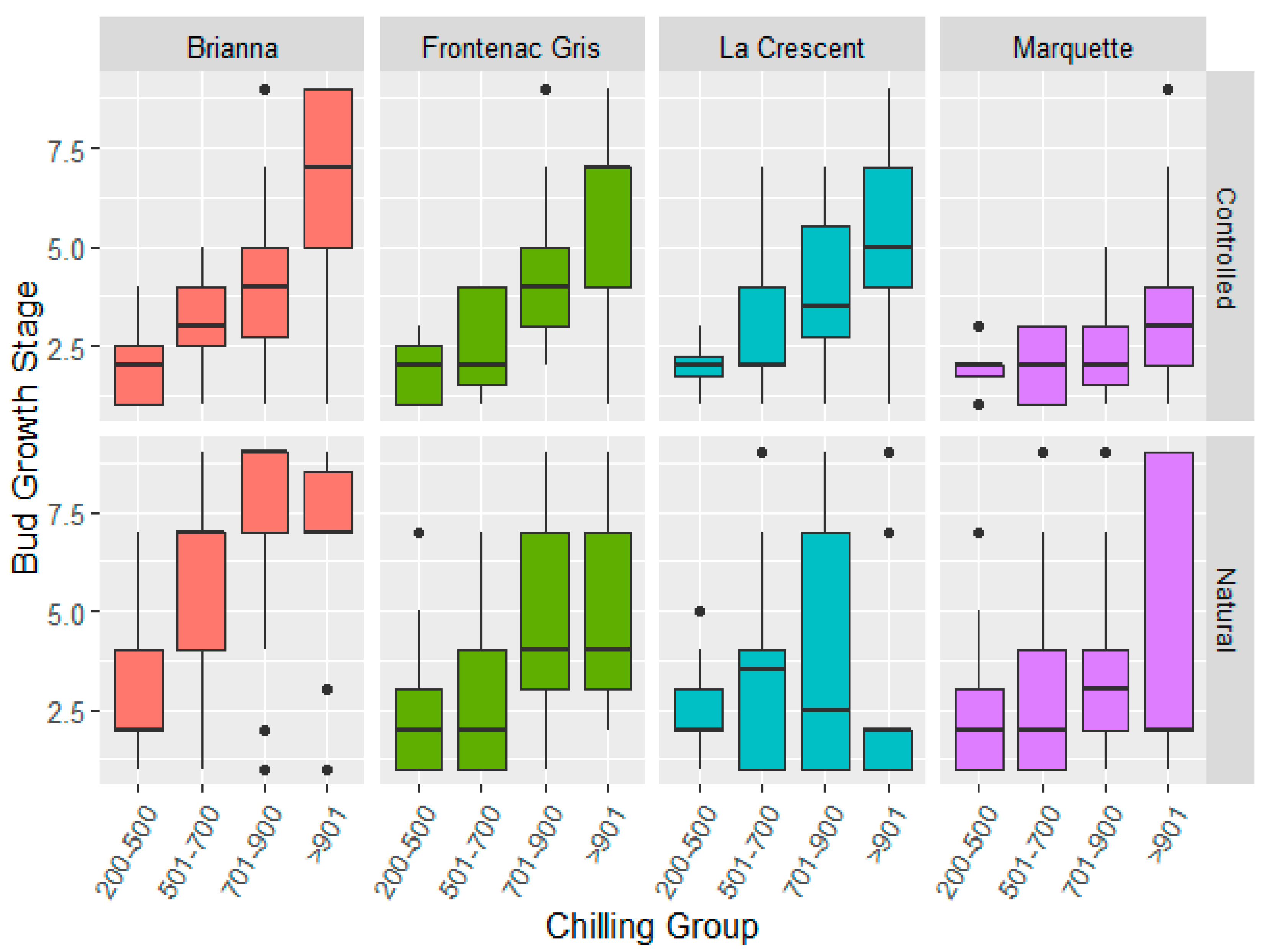
3.3. Bud Dormancy Release Showed Significant Cultivar by Treatment or Season Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fennell, A. Freezing tolerance and injury in grapevines. J. Crop. Improv. 2004, 10, 201–235. [Google Scholar] [CrossRef]
- Svyantek, A.; Köse, B.; Stenger, J.; Auwarter, C.; Hatterman-Valenti, H. Cold-Hardy Grape Cultivar Winter Injury and Trunk Re-Establishment Following Severe Weather Events in North Dakota. Horticulturae 2020, 6, 75. [Google Scholar] [CrossRef]
- Zabadal, T. Winter Injury to Grapevines and Methods of Protection (E2930); Michigan State University: East Lansing, MI, USA, 2015. [Google Scholar]
- Pierquet, P.; Stushnoff, C. Relationship of low temperature exotherms to cold injury in Vitis riparia Michx. Am. J. Enol. Vitic. 1980, 31, 1–6. [Google Scholar]
- Pierquet, P.; Stushnoff, C.; Burke, M. Low temperature exotherms in stem and bud tissues of Vitis riparia Michx [North America]. J. Am. Soc. Hortic. Sci. 1977, 102, 54–55. [Google Scholar]
- Lipe, W.N.; Baumhardt, L.; Wendt, C.; Rayburn, D. Differential thermal analysis of deacclimating Chardonnay and Cabernet Sauvignon grape buds as affected by evaporative cooling. Am. J. Enol. Vitic. 1992, 43, 355–361. [Google Scholar]
- Mills, L.J.; Ferguson, J.C.; Keller, M. Cold-hardiness evaluation of grapevine buds and cane tissues. Am. J. Enol. Vitic. 2006, 57, 194–200. [Google Scholar]
- Londo, J.P.; Kovaleski, A.P. Characterization of wild North American grapevine cold hardiness using differential thermal analysis. Am. J. Enol. Vitic. 2017, 68, 203–212. [Google Scholar] [CrossRef]
- Reynolds, A.G. Grapevine Breeding Programs for the Wine Industry; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Hemstad, P.; Luby, J.J. Utilization of Vitis riparia for the development of new wine varieties with resistance to disease and extreme cold. In Proceedings of the VII International Symposium on Grapevine Genetics and Breeding, Montpellier, France, 6 July 1998; pp. 487–496. [Google Scholar]
- Mesonet at SDSTATE. Available online: https://www.sdstate.edu/mesonet (accessed on 20 November 2020).
- Ferguson, J.C.; Moyer, M.M.; Mills, L.J.; Hoogenboom, G.; Keller, M. Modeling dormant bud cold hardiness and budbreak in twenty-three Vitis genotypes reveals variation by region of origin. Am. J. Enol. Vitic. 2014, 65, 59–71. [Google Scholar] [CrossRef]
- Ferguson, J.C.; Tarara, J.M.; Mills, L.J.; Grove, G.G.; Keller, M. Dynamic thermal time model of cold hardiness for dormant grapevine buds. Ann. Bot. 2011, 107, 389–396. [Google Scholar] [CrossRef]
- Londo, J.P.; Johnson, L.M. Variation in the chilling requirement and budburst rate of wild Vitis species. Environ. Exp. Bot. 2014, 106, 138–147. [Google Scholar] [CrossRef]
- Lang, G.; Early, J.; Martin, G.; Darnell, R. Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar]
- Londo, J.; Kovaleski, A.P. Deconstructing cold hardiness: Variation in supercooling ability and chilling requirements in the wild grapevine Vitis riparia. Aust. J. Grape Wine Res. 2019, 25, 276–285. [Google Scholar] [CrossRef]
- Fuchigami, L.; Weiser, C.; Kobayashi, K.; Timmis, R.; Gusta, L. A degree growth stage (GS) model and cold acclimation in temperate woody plants. Plant cold hardiness and freezing stress. Mech. Crop. Implic. 1982, 2, 93–116. [Google Scholar]
- Meier, M.; Fuhrer, J.; Holzkämper, A. Changing risk of spring frost damage in grapevines due to climate change? A case study in the Swiss Rhone Valley. Int. J. Biometeorol. 2018, 62, 991–1002. [Google Scholar] [CrossRef]
- Maul, E.; Sudharma, K.N.; Ganesch, A.; Brühl, U.; Hundemer, M.; Kecke, S.; Mahler-Ries, A.; Marx, G.; Schreiber, T.; Walk, M.; et al. 30 years VIVC—Vitis International Variety Catalogue. In Proceedings of the XI International Conference on Grapevine Breeding and Genetics, Beijing, China, 28 July–2 August 2014; Available online: www.vivc.de (accessed on 12 December 2020).
- Okie, W. Register of new fruit and nut varieties. HortScience 2004, 39, 1509–1523. [Google Scholar] [CrossRef]
- Luby, J.; Hemstad, P. Grape plant named ‘Frontenac gris’. U.S. Patent Application No. 10/775,435, 25 April 2006. [Google Scholar]
- Okie, W. Register of new fruit and nut varieties list 41. HortScience 2002, 37, 251–272. [Google Scholar] [CrossRef]
- Hemstad, P.; Luby, J. Grapevine plant named ‘Marquette’. U.S. Patent Application No. 11/580,356, 16 December 2008. [Google Scholar]
- USDA. United States Department of Agriculture. USDA Plant Hardiness Zone Map. Available online: https://mauroseed.com/pages/usda-planting-zone-map?gclid=Cj0KCQiAlZH_BRCgARIsAAZHSBlLbpQSJzHpikz5SGDi1FCbvorASxtrspMSoGrDJjBSCyaiENEtRSUaAgawEALw_wcB (accessed on 28 December 2020).
- Coombe, B.G. Growth stages of the grapevine: Adoption of a system for identifying grapevine growth stages. Aust. J. Grape Wine Res. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Dokoozlian, N. Chilling Temperature and Duration Interact on the Budbreak of ‘Perlette’ Grapevine Cuttings. HortScience 1999, 34, 1–3. [Google Scholar] [CrossRef]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; Northwestern University: Evanston, IL, USA, 2020. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 28 December 2020).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wolf, T.K.; Cook, M.K. Seasonal deacclimation patterns of three grape cultivars at constant, warm temperature. Am. J. Enol. Vitic. 1992, 43, 171–179. [Google Scholar]
- Bourne, T.; Moore, J. Cold hardiness in grape cultivar development. Fruit Var. J. 1991, 45, 26–28. [Google Scholar]
- Bourne, T.F.; Moore, J.; George, M.F. Primary bud hardiness of four genotypes of grapes in Arkansas. J. Am. Soc. Hortic. Sci. 1991, 116, 835–837. [Google Scholar] [CrossRef]
- Wolf, T.K.; Cook, M.K. Cold hardiness of dormant buds of grape cultivars: Comparison of thermal analysis and field survival. HortScience 1994, 29, 1453–1455. [Google Scholar] [CrossRef]
- Clark, J.R.; Wolf, T.K.; Warren, M.K. Thermal Analysis of Dormant Buds of Two Muscadine Grape Cultivars and of Vitis labrusca L.‘Mars’. HortScience 1996, 31, 79–81. [Google Scholar] [CrossRef]
- Sanliang Gu, S.; Shufu Dong, S.; Jianqiang Li, J.; Susanne Howard, S. Acclimation and deacclimation of primary bud cold hardiness in ‘Norton’, ‘Vignoles’ and ‘St. Vincent’grapevines. J. Hortic. Sci. Biotechnol. 2001, 76, 655–660. [Google Scholar] [CrossRef]
- Cragin, J.; Serpe, M.; Keller, M.; Shellie, K. Dormancy and cold hardiness transitions in winegrape cultivars Chardonnay and Cabernet Sauvignon. Am. J. Enol. Vitic. 2017, 68, 195–202. [Google Scholar] [CrossRef]
- Atucha, A.; Hedtcke, J.; Workmaster, B.A. Evaluation of cold-climate interspecific hybrid wine grape cultivars for the upper Midwest. J. Am. Pomol. Soc 2018, 72, 80–93. [Google Scholar]
- Berkett, L.P.; Bradshaw, T.L.; Kingsley-Richards, S.L.; Cromwell, M.L. 2008 Grape Bud Survival on Eight Winegrape Cultivars in Vermont. In Proceedings of the 2nd Annual National Viticulture Research Conference, University of California-Davis, Davis, CA, USA, 9–11 July 2008; p. 11. [Google Scholar]
- Domoto, P.A.; Nonnecke, G.R.; Hannan, J.M.; Portz, D.N.; Riesselman, L.B.; Havlovic, B.J.; Howell, N.P.; Pecinovsky, K.T.; Van Dee, K. 2003 Wine Grape Cultivar Trial Performance; RFR A1039; Iowa State University: Ames, IA, USA, 2011. [Google Scholar]
- Domoto, P.A.; Nonnecke, G.R.; Tabor, P.; Riesselman, L.B. Cold Hardy Wine Grape Cultivar Trial; RFR A1214; Iowa State University: Ames, IA, USA, 2013. [Google Scholar]
- Kovaleski, A.P.; Reisch, B.I.; Londo, J.P. Deacclimation kinetics as a quantitative phenotype for delineating the dormancy transition and thermal efficiency for budbreak in Vitis species. AoB Plants 2018, 10, ply066. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cultivar | 2017/18 | 2018/19 | 2019/2020 | All Seasons |
|---|---|---|---|---|
| Mean across cultivars | 0.46 * | 0/28 * | 0.05 | 0.23 * |
| Brianna | 0.45 * | 0.45 * | 0.06 | 0.27 * |
| Frontenac gris | 0.52 * | 0.08 * | 0..11 | 0.18 * |
| La Crescent | 0.45 * | 0.40 * | 0.01 | 0.25 * |
| Marquette | 0.43 * | 0.28 * | 0.05 | 0.25 * |
| Season(s) | Factors in ANOVA | ||||
|---|---|---|---|---|---|
| Season | Cultivar | Sampling Time | Cultivar × Sampling Time | Cultivar × Season | |
| 2017/18 | NA | * | * | * | NA |
| 2018/19 | NA | * | * | * | NA |
| 2019/20 | NA | NS | * | * | NA |
| 2017/18 + 2018/19 | * | * | * | * | NA |
| 2017/18 + 2018/19 | * | * | * | * | * |
| 2017/18 + 2018/19 + 2019/20 | * | * | * | * | NA |
| 2017/18 + 2018/19 + 2019/20 | * | NS | * | * | * |
| Cultivar | Condition | Mean Bud Phenology Stage (Standard Deviation) in Chilling Groups | |||
|---|---|---|---|---|---|
| 200–500 | 501–700 | 701–900 | >901 | ||
| Brianna (a) | Natural | 2.8 ± 1.6 | 5.9 ± 2.4 | 7.7 ± 2.3 | 6.6 ± 2.5 |
| Controlled | 2.0 ± 1.2 | 3.3 ± 0.7 | 4.2 ± 2.2 | 7.2 ± 2.2 | |
| Frontenac gris (b) | Natural | 2.3 ± 1.6 | 3.0 ± 2.2 | 5.1 ± 2.8 | 4.9 ± 2.6 |
| Controlled | 2.1 ± 0.9 | 2.2 ± 1.3 | 4.4 ± 2.5 | 6.0 ± 2.5 | |
| La Crescent (b) | Natural | 2.4 ± 1.1 | 3.5 ± 2.4 | 3.9 ± 3.1 | 2.7 ± 2.7 |
| Controlled | 2.1 ± 0.8 | 2.8 ± 1.9 | 4.2 ± 2.2 | 5.6 ± 2.1 | |
| Marquette (c) | Natural | 2.6 ± 1.6 | 2.7 ± 2.0 | 3.4 ± 2.4 | 4.6 ± 3.6 |
| Controlled | 2.0 ± 0.7 | 1.9 ± 1.0 | 2.2 ± 1.2 | 3.8 ± 2.3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yilmaz, T.; Alahakoon, D.; Fennell, A. Freezing Tolerance and Chilling Fulfillment Differences in Cold Climate Grape Cultivars. Horticulturae 2021, 7, 4. https://doi.org/10.3390/horticulturae7010004
Yilmaz T, Alahakoon D, Fennell A. Freezing Tolerance and Chilling Fulfillment Differences in Cold Climate Grape Cultivars. Horticulturae. 2021; 7(1):4. https://doi.org/10.3390/horticulturae7010004
Chicago/Turabian StyleYilmaz, Turhan, Dilmini Alahakoon, and Anne Fennell. 2021. "Freezing Tolerance and Chilling Fulfillment Differences in Cold Climate Grape Cultivars" Horticulturae 7, no. 1: 4. https://doi.org/10.3390/horticulturae7010004
APA StyleYilmaz, T., Alahakoon, D., & Fennell, A. (2021). Freezing Tolerance and Chilling Fulfillment Differences in Cold Climate Grape Cultivars. Horticulturae, 7(1), 4. https://doi.org/10.3390/horticulturae7010004