The Relationship between Fruit Size and Phenolic and Enzymatic Composition of Avocado Byproducts (Persea americana Mill.): The Importance for Biorefinery Applications

 , and
, and
Abstract
1. Introduction
2. Material and Methods
2.1. Chemicals
2.2. Sampling and Ripeness State Evaluation
2.3. Extraction Procedure
2.4. Phytochemical Characterization of APEs
2.4.1. TPC
2.4.2. TFC
2.4.3. DPPH Antioxidant Capacity
2.5. HPLC–UV/vis Individual Phenolics Determination
2.6. Enzymatic Activity Measurement
2.6.1. PAL Activity
2.6.2. CHS Activity
2.6.3. PPO Activity
2.7. Statistical Analysis
3. Results and Discussion
3.1. Size and Ripeness State Evaluation of Avocado Fruits
3.2. Phytochemicals and Antioxidant Characteristics of the APEs
3.3. HPLC–UV/vis of Individual Phenolic Content in S, M, and L APEs
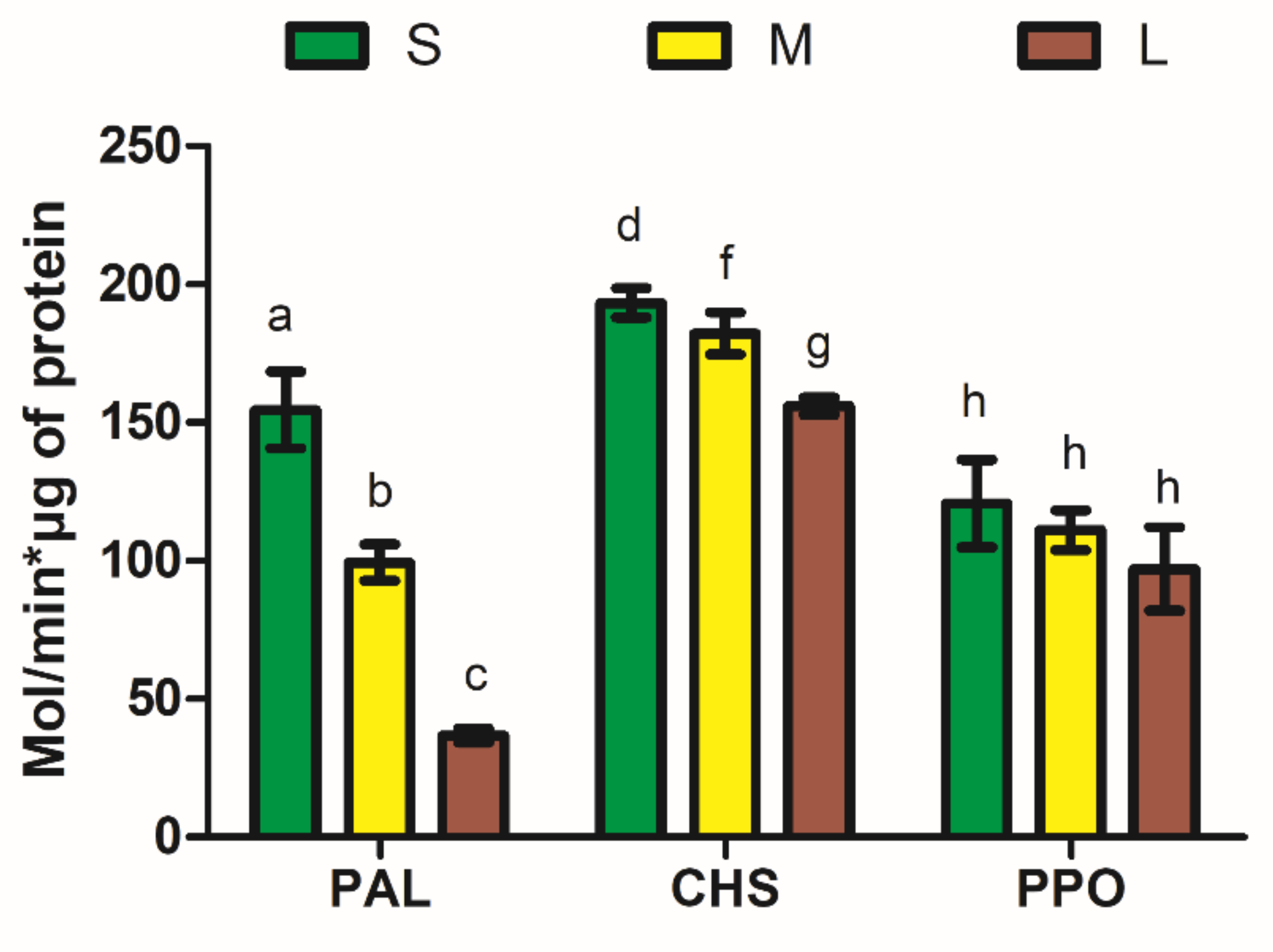
3.4. PAL, CHS, and PPO Activities in the Peel of Avocado Sorted by Size
3.5. Statistical Correlation between the Phytochemical Parameters and Avocado Physiology
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schaffer, B.; Wolstenholme, N.; Whiley, A. The Avocado, Botany, Production and Uses, 2nd ed.; Schaffer, B., Wolstenholme, B.N., Whiley, A., Eds.; CABI: Oxfordshire, UK, 2013; ISBN 9788578110796. [Google Scholar]
- Silber, A.; Naor, A.; Cohen, H.; Bar-Noy, Y.; Yechieli, N.; Levi, M.; Noy, M.; Peres, M.; Duari, D.; Narkis, K.; et al. Irrigation of ‘Hass’ avocado: Effects of constant vs. temporary water stress. Irrig. Sci. 2019, 37, 451–460. [Google Scholar] [CrossRef]
- Cowan, A.K.; Cripps, R.F.; Richings, E.W.; Taylor, N.J. Fruit size: Towards an understanding of the metabolic control of fruit growth using avocado as a model system. Physiol. Plant. 2001, 111, 127–136. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Celedón, J.M.; Gil, P.M.; Ferreyra, R.; Maldonado, P.; Barrera, C. Sensitivity and Variability of Two Plant Water Stress Indicators: Exploring Criteria for Choosing a plant Monitoring Method for Avocado Irrigation Management. Chil. J. Agric. Res. 2012, 72, 379–387. [Google Scholar] [CrossRef]
- Colombo, R.; Papetti, A. Avocado (Persea americana Mill.) by-products and their impact: From bioactive compounds to biomass energy and sorbent material for removing contaminants. A review. Int. J. Food Sci. Technol. 2019, 54, 943–951. [Google Scholar] [CrossRef]
- Christopoulos, M.V.; Tsantili, E. Participation of phenylalanine ammonia-lyase (PAL) in increased phenolic compounds in fresh cold stressed walnut (Juglans regia L.) kernels. Postharvest Biol. Technol. 2015, 104, 17–25. [Google Scholar] [CrossRef]
- Morais, D.R.; Rotta, E.M.; Sargi, S.C.; Schmidt, E.M.; Bonafe, E.G.; Eberlin, M.N.; Sawaya, A.C.H.F.; Visentainer, J.V. Antioxidant activity, phenolics and UPLC-ESI(-)-MS of extracts from different tropical fruits parts and processed peels. Food Res. Int. 2015, 77, 392–399. [Google Scholar] [CrossRef]
- Pathak, P.D.; Mandavgane, S.A.; Puranik, N.M.; Jambhulkar, S.J.; Kulkarni, B.D. Valorization of potato peel: A biorefinery approach. Crit. Rev. Biotechnol. 2018, 38, 218–230. [Google Scholar] [CrossRef]
- Trujillo-Mayol, I.; Alarcón-Enos, J.; Céspedes-Acuña, C.; Silva, F.L. Improvement of the polyphenol extraction from avocado peel by assisted ultrasound and microwaves. Food Process Eng. 2019, 42, 1–11. [Google Scholar] [CrossRef]
- Villa-Rodríguez, J.A.; Molina-Corral, F.J.; Ayala-Zavala, J.F.; Olivas, G.I.; González-Aguilar, G.A. Effect of maturity stage on the content of fatty acids and antioxidant activity of “Hass” avocado. Food Res. Int. 2011, 44, 1231–1237. [Google Scholar] [CrossRef]
- Herrera-González, J.A.; Salazar-García, S.; Martínez-Flores, H.E.; Ruiz-García, J.E. Indicadores preliminares de madurez fisiológica y comportamiento postcosecha del fruto de aguacate méndez. Rev. Fitotec. Mex. 2017, 40, 55–63. [Google Scholar] [CrossRef]
- Márquez, C.J.; Yepes, D.P.; Sanchez, L. Changes physical-chemical of avocado (Persea americana Mill. cv. “Hass”) in postharvest for two municipalities of antioquia. Temas Agrar. 2014, 19, 32–47. [Google Scholar] [CrossRef]
- Sahpazidou, D.; Geromichalos, G.D.; Stagos, D.; Apostolou, A.; Haroutounian, S.A.; Tsatsakis, A.M.; Tzanakakis, G.N.; Hayes, A.W.; Kouretas, D. Anticarcinogenic activity of polyphenolic extracts from grape stems against breast, colon, renal and thyroid cancer cells. Toxicol. Lett. 2014, 230, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Piechowiak, T.; Balawejder, M. Impact of ozonation process on the level of selected oxidative stress markers in raspberries stored at room temperature. Food Chem. 2019, 298, 125093. [Google Scholar] [CrossRef]
- Ardila, H.D.; Martínez, S.T.; Higuera, B.L. Levels of constitutive flavonoid biosynthetic enzymes in carnation (Dianthus caryophyllus L.) cultivars with differential response to Fusarium oxysporum f. sp. dianthi. Acta Physiol. Plant. 2013, 35, 1233–1245. [Google Scholar] [CrossRef]
- Kumar, V.B.A.; Mohan, T.C.K.; Murugan, K. Purification and kinetic characterization of polyphenol oxidase from Barbados cherry (Malpighia glabra L.). Food Chem. 2008, 110, 328–333. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Nasiri, A.; Jaafar, H.Z.E.; Baghdadi, A.; Ahmad, I. Changes in phytochemical synthesis, chalcone synthase activity and pharmaceutical qualities of sabah snake grass (Clinacanthus nutans L.) in relation to plant age. Molecules 2014, 19, 17632–17648. [Google Scholar] [CrossRef]
- Niphadkar, S.S.; Vetal, M.D.; Rathod, V.K. Purification and Characterization of Polyphenol Oxidase from Waste Potato Peel by Aqueous Two-Phase Extraction. Prep. Biochem. Biotechnol. 2015, 45, 632–649. [Google Scholar] [CrossRef]
- Cox, K.A.; McGhie, T.K.; White, A.; Woolf, A.B. Skin colour and pigment changes during ripening of “Hass” avocado fruit. Postharvest Biol. Technol. 2004, 31, 287–294. [Google Scholar] [CrossRef]
- López-Cobo, A.; Gómez-Caravaca, A.M.; Pasini, F.; Caboni, M.F.; Segura-Carretero, A.; Fernández-Gutiérrez, A. HPLC-DAD-ESI-QTOF-MS and HPLC-FLD-MS as valuable tools for the determination of phenolic and other polar compounds in the edible part and by-products of avocado. Food Sci. Technol. 2016, 73, 505–513. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Y.; Khuong, T.; Lovatt, C.J. Developmental differences in antioxidant compounds and systems in normal and small-phenotype fruit of “Hass” avocado (Persea americana Mill.). Sci. Hortic. 2016, 206, 15–23. [Google Scholar] [CrossRef]
- Villangó, S.; Szekeres, A.; Bencsik, O.; Láposi, R.; Pálfi, Z.; Zsófi, Z. The effect of postveraison water deficit on the phenolic composition and concentration of the Kékfrankos (Vitis vinifera L.) berry. Sci. Hortic. 2016, 209, 113–116. [Google Scholar] [CrossRef]
- Roby, G.; Harbertson, J.F.; Adams, D.A.; Matthews, M.A. Berry size and vine water deficits as factors in Winegrape composition: Anthocyanins and tannins. Aust. J. Grape Wine Res. 2004, 10, 100–107. [Google Scholar] [CrossRef]
- Quan, N.; Anh, L.; Khang, D.; Tuyen, P.; Toan, N.; Minh, T.; Minh, L.; Bach, D.; Ha, P.; Elzaawely, A.; et al. Involvement of Secondary Metabolites in Response to Drought Stress of Rice (Oryza sativa L.). Agriculture 2016, 6, 23. [Google Scholar] [CrossRef]
- Bautista, I.; Boscaiu, M.; Lidón, A.; Llinares, J.V.; Lull, C.; Donat, M.P.; Mayoral, O.; Vicente, O. Environmentally induced changes in antioxidant phenolic compounds levels in wild plants. Acta Physiol. Plant. 2016, 38, 1–15. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Li, Y.; Guo, T. Expression of flavonoid biosynthesis genes and accumulation of flavonoid in wheat leaves in response to drought stress. Plant Physiol. Biochem. 2014, 80, 60–66. [Google Scholar] [CrossRef]
- Keller, M.; Rogiers, S.Y.; Schultz, R. Nitrogen and ultraviolet radiation modify grapewines′ susceptibility to powdery midew. Vitis 2003, 42, 87–94. [Google Scholar]
- Haminiuk, C.W.I.; Maciel, G.M.; Plata-Oviedo, M.S.V.; Peralta, R.M. Phenolic compounds in fruits—An overview. Int. J. Food Sci. Technol. 2012, 47, 2023–2044. [Google Scholar] [CrossRef]
- Azmir, J.; Zaidul, I.S.M.; Rahman, M.M.; Sharif, K.M.; Mohamed, A.; Sahena, F.; Jahurul, M.H.A.; Ghafoor, K.; Norulaini, N.A.N.; Omar, A.K.M. Techniques for extraction of bioactive compounds from plant materials: A review. J. Food Eng. 2013, 117, 426–436. [Google Scholar] [CrossRef]
- Tiwari, B.K. Ultrasound: A clean, green extraction technology. Trends Anal. Chem. 2015, 71, 100–109. [Google Scholar] [CrossRef]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and agri-industrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Melgar, B.; Dias, M.I.; Ciric, A.; Sokovic, M.; Garcia-Castello, E.M.; Rodriguez-Lopez, A.D.; Barros, L.; Ferreira, I.C.R.F. Bioactive characterization of Persea americana Mill. by-products: A rich source of inherent antioxidants. Ind. Crops Prod. 2018, 111, 212–218. [Google Scholar] [CrossRef]
- Neilson, A.P.; O’Keefe, S.F.; Bolling, B.W. High-Molecular-Weight Proanthocyanidins in Foods: Overcoming Analytical Challenges in Pursuit of Novel Dietary Bioactive Components. Annu. Rev. Food Sci. Technol. 2016, 7, 43–64. [Google Scholar] [CrossRef]
- Mayer, R.; Stecher, G.; Wuerzner, R.; Silva, R.C.; Sultana, T.; Trojer, L.; Feuerstein, I.; Krieg, C.; Abel, G.; Popp, M.; et al. Proanthocyanidins: Target compounds as antibacterial agents. J. Agric. Food Chem. 2008, 56, 6959–6966. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Xiao, K.; Li, B.; Jiang, W.; Peng, W.; Zheng, J.; Zhou, H. The combination of catechin and epicatechin gallate from Fructus crataegi potentiates β-lactam antibiotics against Methicillin-Resistant Staphylococcus aureus (MRSA) in vitro and in vivo. Int. J. Mol. Sci. 2013, 14, 1802–1821. [Google Scholar] [CrossRef] [PubMed]
- Miklasińska, M.; Kȩpa, M.; Wojtyczka, R.D.; Idzik, D.; Dziedzic, A.; Wąsik, T.J. Catechin hydrate augments the antibacterial action of selected antibiotics against Staphylococcus aureus clinical strains. Molecules 2016, 21, 244. [Google Scholar] [CrossRef] [PubMed]
- Torres, E.; Garcia, A.; Aranda, M.; Saéz, V.; Zúñiga, F.; Alarcón, J.; Avello, M.; Pastene, E. One-step purification of two semi-synthetic epicatechin adducts prepared from avocado peels procyanidins by centrifugal partition chromatography and evaluation of their anti-inflammatory effects on adenocarcinoma gastric cells infected with Helicobacter pylori. J. Chil. Chem. Soc. 2018, 4, 4222–4228. [Google Scholar] [CrossRef]
- Wong, X.; Carrasco-Pozo, C.; Escobar, E.; Navarrete, P.; Blachier, F.; Andriamihaja, M.; Lan, A.; Tomé, D.; Cires, M.J.; Pastene, E.; et al. Deleterious Effect of p-Cresol on Human Colonic Epithelial Cells Prevented by Proanthocyanidin-Containing Polyphenol Extracts from Fruits and Proanthocyanidin Bacterial Metabolites. J. Agric. Food Chem. 2016, 64, 3574–3583. [Google Scholar] [CrossRef]
- Cires, M.J.; Navarrete, P.; Pastene, E.; Carrasco-Pozo, C.; Valenzuela, R.; Medina, D.A.; Andriamihaja, M.; Beaumont, M.; Blachier, F.; Gotteland, M. Protective Effect of an Avocado Peel Polyphenolic Extract Rich in Proanthocyanidins on the Alterations of Colonic Homeostasis Induced by a High-Protein Diet. J. Agric. Food Chem. 2019, 67, 11616–11626. [Google Scholar] [CrossRef]
- Sawai-Kuroda, R.; Kikuchi, S.; Shimizu, Y.K.; Sasaki, Y.; Kuroda, K.; Tanaka, T.; Yamamoto, T.; Sakurai, K.; Shimizu, K. A polyphenol-rich extract from Chaenomeles sinensis (Chinese quince) inhibits influenza A virus infection by preventing primary transcription in vitro. J. Ethnopharmacol. 2013, 146, 866–872. [Google Scholar] [CrossRef]
- Russo, M.; Moccia, S.; Spagnuolo, C.; Tedesco, I.; Russo, G.L. Roles of flavonoids against coronavirus infection. Chem. Biol. Interact. 2020, 328, 109211. [Google Scholar] [CrossRef] [PubMed]
- Quiles, J.L.; Rivas-García, L.; Varela-López, A.; Llopis, J.; Battino, M.; Sánchez-González, C. Do nutrients and other bioactive molecules from foods have anything to say in the treatment against COVID-19? Environ. Res. 2020, 191, 110053. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Silván, J.M.; Assar, S.H.; Srey, C.; Dolores, M.; Ames, J.M. Control of the Maillard reaction by ferulic acid. Food Chem. 2011, 128, 208–213. [Google Scholar] [CrossRef]
- Kuppusamy, S.; Thavamani, P.; Megharaj, M.; Naidu, R. Bioremediation potential of natural polyphenol rich green wastes: A review of current research and recommendations for future directions. Environ. Technol. Innov. 2015, 4, 17–28. [Google Scholar] [CrossRef]
- Araújo, R.G.; Rodriguez-Jasso, R.M.; Ruiz, H.A.; Pintado, M.M.E.; Aguilar, C.N. Avocado by-products: Nutritional and functional properties. Trends Food Sci. Technol. 2018, 80, 51–60. [Google Scholar] [CrossRef]
- Geerkens, C.H.; Schweiggert, R.M.; Steingass, H.; Boguhn, J.; Rodehutscord, M.; Carle, R. Influence of apple and citrus pectins, processed mango peels, a phenolic mango peel extract, and gallic acid as potential feed supplements on in vitro total gas production and rumen methanogenesis. J. Agric. Food Chem. 2013, 61, 5727–5737. [Google Scholar] [CrossRef]
- Rubin, I.; Oliveira, D.; Rodrigues, G.; Severo, J.; Renard, C.M.G.C.; Clasen, F.; Valmor, C. Plant Physiology and Biochemistry Preharvest UV-C radiation influences physiological, biochemical, and transcriptional changes in strawberry cv. Camarosa. Plant Physiol. Biochem. 2016, 108, 391–399. [Google Scholar] [CrossRef]
- Sheng, K.; Zheng, H.; Shui, S.S.; Yan, L.; Liu, C.; Zheng, L. Comparison of postharvest UV-B and UV-C treatments on table grape: Changes in phenolic compounds and their transcription of biosynthetic genes during storage. Postharvest Biol. Technol. 2018, 138, 74–81. [Google Scholar] [CrossRef]
- Cirilli, M.; Caruso, G.; Gennai, C.; Urbani, S.; Frioni, E.; Ruzzi, M.; Servili, M.; Gucci, R.; Poerio, E.; Muleo, R. The role of polyphenoloxidase, peroxidase, and β-glucosidase in phenolics accumulation in Olea europaea L. Fruits under different water regimes. Front. Plant Sci. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Toledo, L.; Aguirre, C. Enzymatic browning in avocado (Persea americana) revisited: History, advances, and future perspectives. Crit. Rev. Food Sci. Nutr. 2017, 57, 3860–3872. [Google Scholar] [CrossRef] [PubMed]
- Schovánková, J.; Opatová, H. Changes in phenols composition and activity of phenylalanine-ammonia lyase in apples after fungal infections. Hortic. Sci. 2011, 38, 1–10. [Google Scholar] [CrossRef]
- Koca, N.; Karaman, Ş. The effects of plant growth regulators and L-phenylalanine on phenolic compounds of sweet basil. Food Chem. 2015, 166, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Matharu, A.S.; de Melo, E.M.; Houghton, J.A. Opportunity for high value-added chemicals from food supply chain wastes. Bioresour. Technol. 2016, 215, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Schieber, A. Side Streams of Plant Food Processing as a Source of Valuable Compounds: Selected Examples. Annu. Rev. Food Sci. Technol. 2017, 8, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Chemat, F.; Rombaut, N.; Meullemiestre, A.; Turk, M.; Perino, S.; Fabiano-Tixier, A.S.; Abert-Vian, M. Review of Green Food Processing techniques. Preservation, transformation, and extraction. Innov. Food Sci. Emerg. Technol. 2017, 41, 357–377. [Google Scholar] [CrossRef]
- Ciriminna, R.; Carnaroglio, D.; Delisi, R.; Arvati, S.; Tamburino, A.; Pagliaro, M. Industrial Feasibility of Natural Products Extraction with Microwave Technology. ChemistrySelect 2016, 1, 549–555. [Google Scholar] [CrossRef]
- López, J.A.S.; Li, Q.; Thompson, I.P. Biorefinery of waste orange peel. Crit. Rev. Biotechnol. 2010, 30, 63–69. [Google Scholar] [CrossRef]
- Cheok, C.Y.; Mohd Adzahan, N.; Abdul Rahman, R.; Zainal Abedin, N.H.; Hussain, N.; Sulaiman, R.; Chong, G.H. Current trends of tropical fruit waste utilization. Crit. Rev. Food Sci. Nutr. 2018, 58, 335–361. [Google Scholar] [CrossRef]
- Dávila, J.A.; Rosenberg, M.; Castro, E.; Cardona, C.A. A model biorefinery for avocado (Persea americana Mill.) processing. Bioresour. Technol. 2017, 243, 17–29. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Size z | Weight kg | Volume m3 | Firmness N | Color y | ||
|---|---|---|---|---|---|---|
| L* | a* | b* | ||||
| S | 9.4 × 10−4 ± 3.7 × 10−5 a x | 8.5 × 10−4 ± 6.6 × 10−4 a | 9.6 ± 0.5 a | 25.1 ± 0.7 a | 0.1 ± 0.4 a | 4.0 ± 0.3 a |
| M | 2.4 × 10−4 ± 1.5 × 10−5 b | 1.9 × 10−3 ± 2.0 × 10−4 b | 9.8 ± 0.5 a | 25.6 ± 0.4 a | −0.1± 0.2 a | 3.7 ± 0.3 a |
| L | 3.4 × 10−4 ± 2.6 × 10−5 c | 3.3 × 10−3 ± 1.8 × 10−4 c | 10.2 ± 0.4 a | 24.7± 0.6 a | 0.2 ± 0.1 a | 0.9 ± 0.2 a |
| Size | TPC mg GAE/g DM z | TFC µg QuE/g DM y | DPPH Inhibition µg TE/g DM x |
|---|---|---|---|
| S | 93.7 ± 0.4 a | 803.1 ± 89.3 b | 160.9 ± 1.6 a |
| M | 92.5 ± 3.2 a | 600.7 ± 31.1 a | 160.6 ± 2.8 a |
| L | 33.3 ± 2.9 b | 580.2 ± 34.9 a | 148.2 ± 5.2 b |
| TFC z | DPPH | PAL | CHS | PPO | Weight | Volume | |
|---|---|---|---|---|---|---|---|
| TPC | 0.5514 y | 0.9753 | 0.8847 | 0.9131 | 0.6220 | −0.8118 | −0.8943 |
| NS x | ** | ** | ** | NS | * | ** | |
| TFC | 0.5663 | 0.8286 | 0.6938 | 0.5330 | −0.9002 | −0.8282 | |
| NS | * | * | NS | ** | * | ||
| DPPH | 0.8441 | 0.9088 | 0.5144 | −0.8129 | −0.8607 | ||
| * | ** | NS | * | * | |||
| PAL | 0.8986 | 0.7202 | −0.9626 | −0.9859 | |||
| * | * | ** | ** | ||||
| CHS | 0.5194 | −0.9157 | −0.9334 | ||||
| ** | ** | ||||||
| PPO | −0.5863 | −0.7319 | |||||
| * | |||||||
| Weight | 0.9596 | ||||||
| ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trujillo-Mayol, I.; Badillo-Muñoz, G.; Céspedes-Acuña, C.; Alarcón-Enos, J. The Relationship between Fruit Size and Phenolic and Enzymatic Composition of Avocado Byproducts (Persea americana Mill.): The Importance for Biorefinery Applications. Horticulturae 2020, 6, 91. https://doi.org/10.3390/horticulturae6040091
Trujillo-Mayol I, Badillo-Muñoz G, Céspedes-Acuña C, Alarcón-Enos J. The Relationship between Fruit Size and Phenolic and Enzymatic Composition of Avocado Byproducts (Persea americana Mill.): The Importance for Biorefinery Applications. Horticulturae. 2020; 6(4):91. https://doi.org/10.3390/horticulturae6040091
Chicago/Turabian StyleTrujillo-Mayol, Igor, Guillermo Badillo-Muñoz, Carlos Céspedes-Acuña, and Julio Alarcón-Enos. 2020. "The Relationship between Fruit Size and Phenolic and Enzymatic Composition of Avocado Byproducts (Persea americana Mill.): The Importance for Biorefinery Applications" Horticulturae 6, no. 4: 91. https://doi.org/10.3390/horticulturae6040091
APA StyleTrujillo-Mayol, I., Badillo-Muñoz, G., Céspedes-Acuña, C., & Alarcón-Enos, J. (2020). The Relationship between Fruit Size and Phenolic and Enzymatic Composition of Avocado Byproducts (Persea americana Mill.): The Importance for Biorefinery Applications. Horticulturae, 6(4), 91. https://doi.org/10.3390/horticulturae6040091