Changes in Acetylene Reduction Activities and nifH Genes Associated with Field-Grown Sweet Potatoes with Different Nursery Farmers and Cultivars


Abstract
1. Introduction
2. Materials and Methods
2.1. Cultivation and Sampling of Sweet Potatoes
2.2. ARA
2.3. Nitrogen Content and Natural 15N Abundance
2.4. Analysis of nifH Genes Prepared from Plant Samples
2.5. Statistical Analysis
3. Results
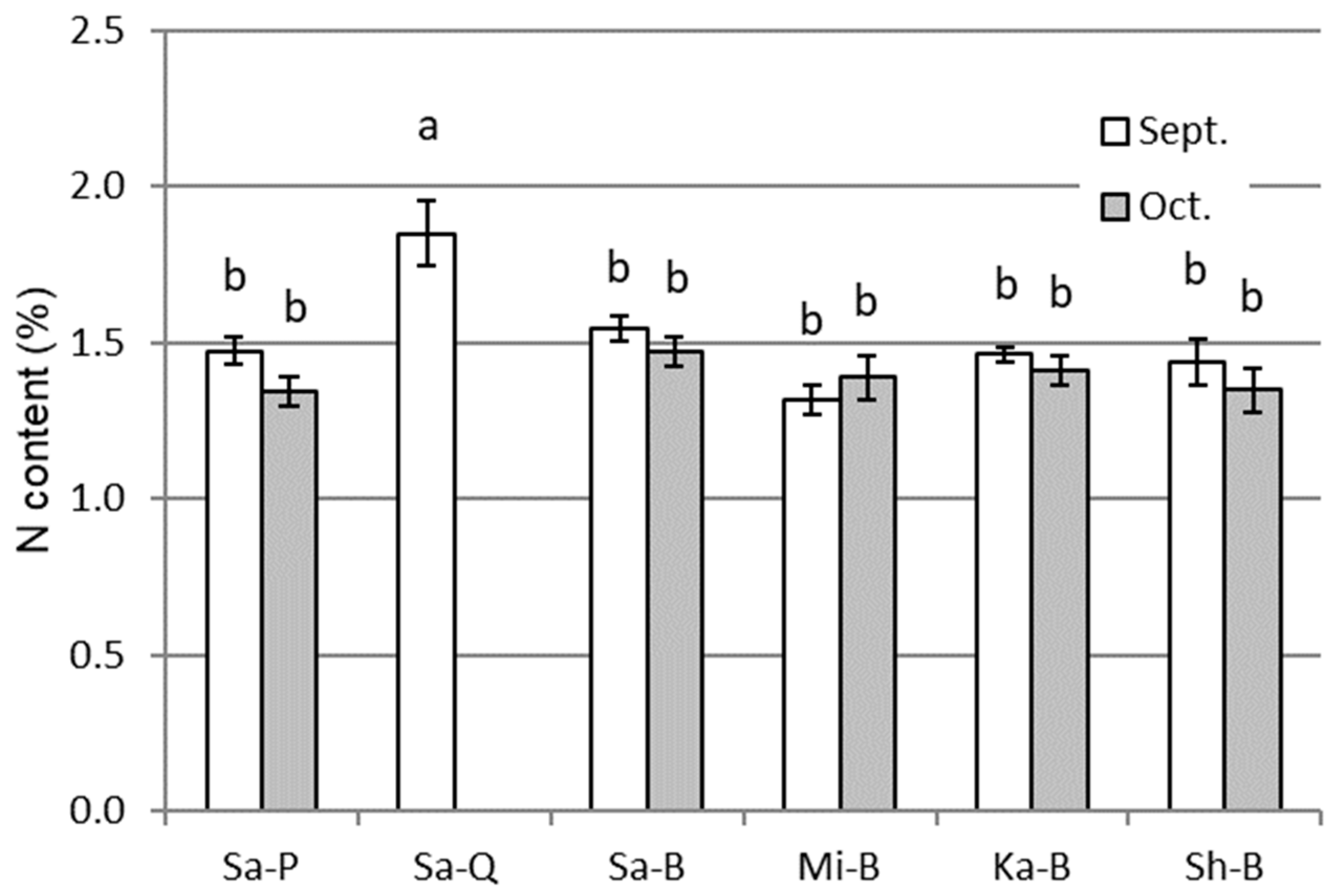
3.1. Plant Growth and Nitrogen Content
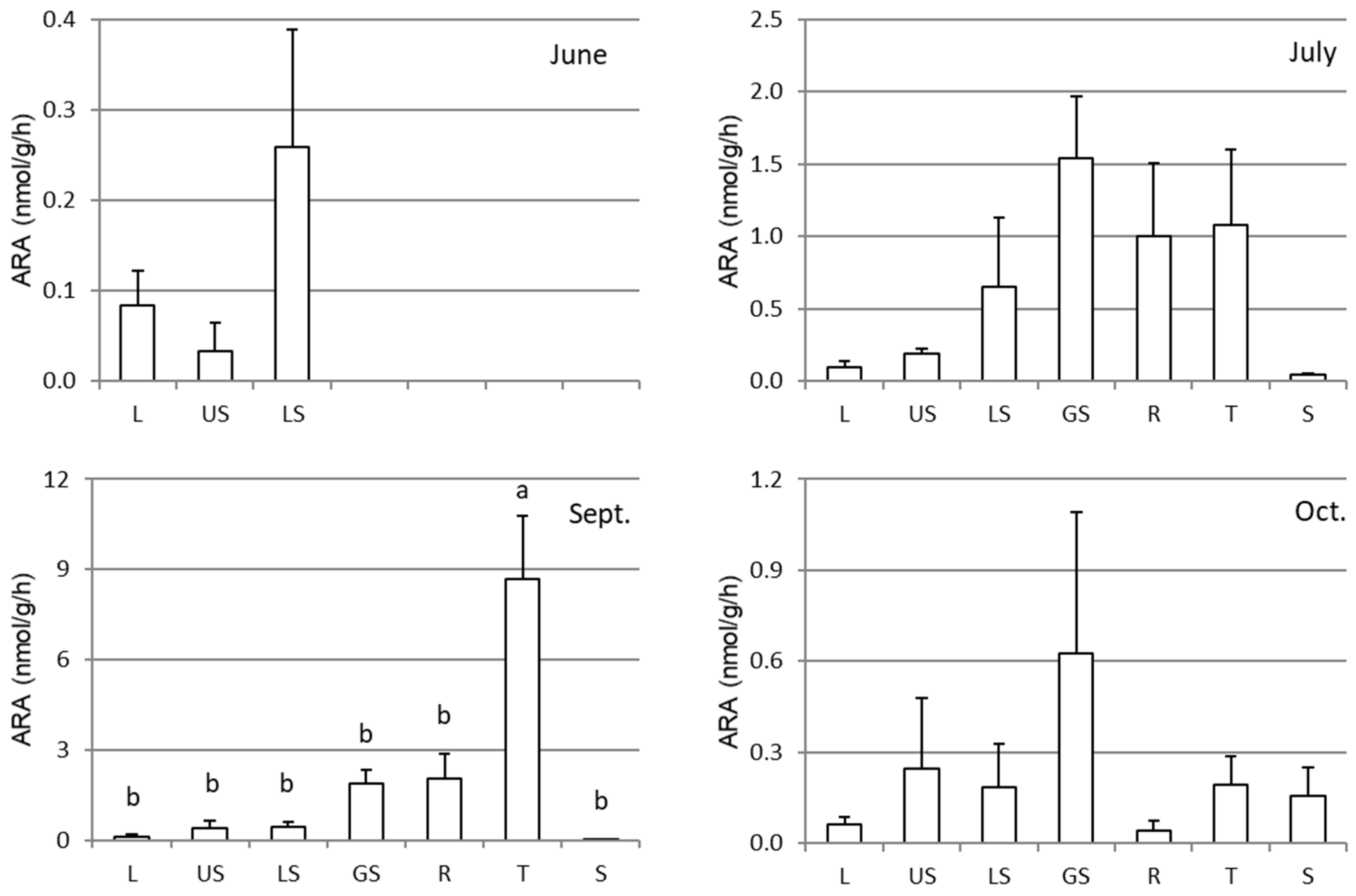
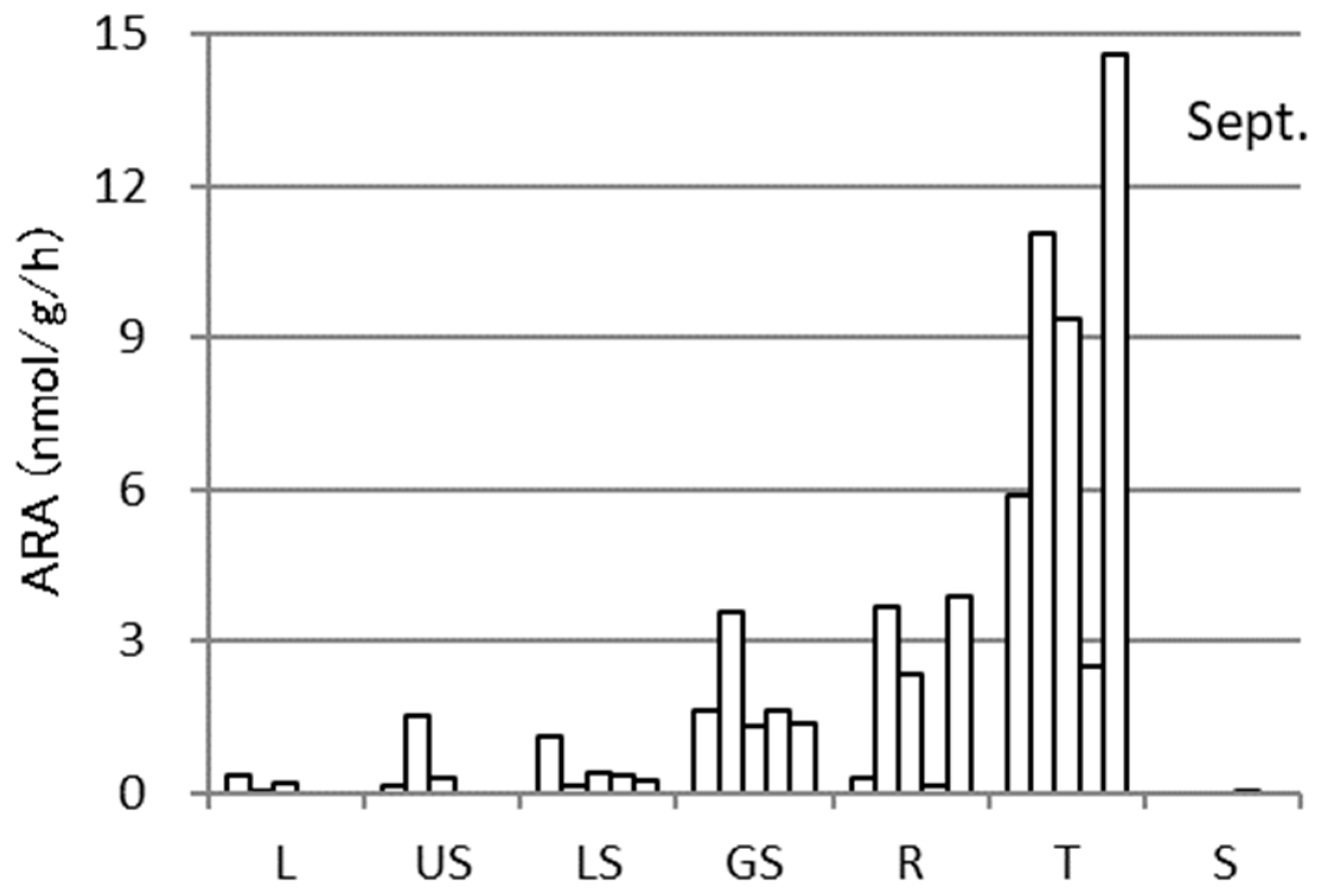
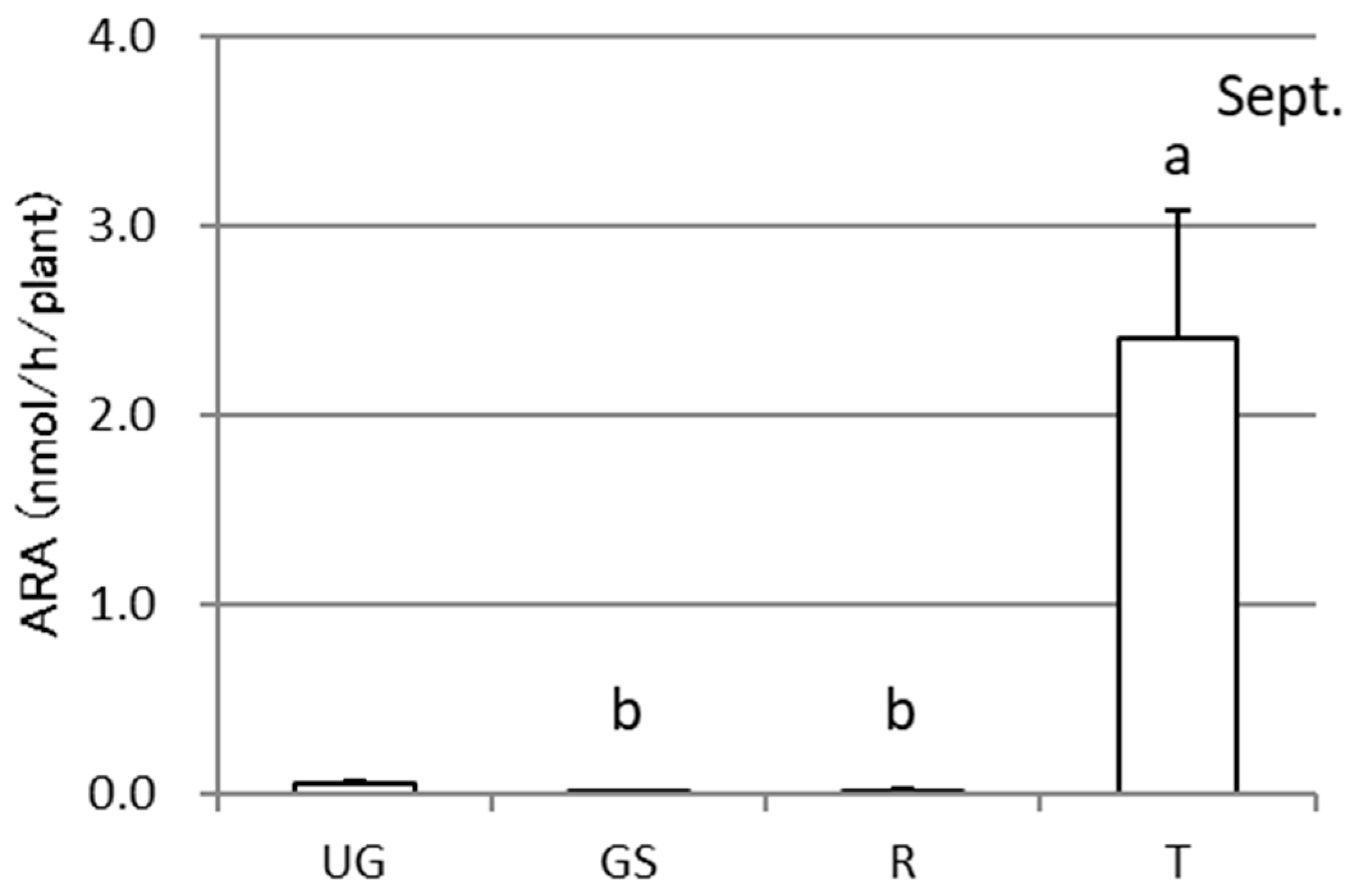
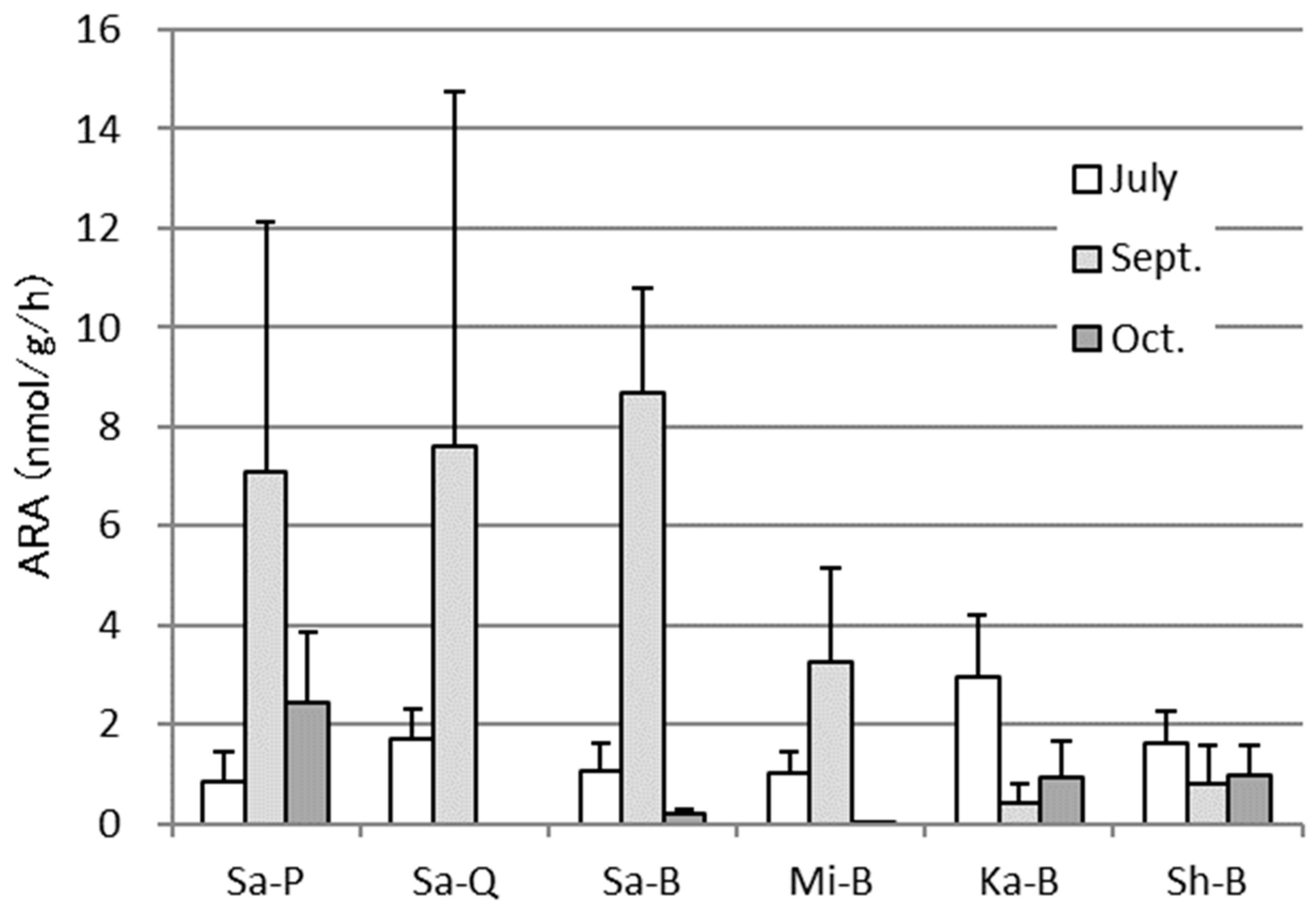
3.2. ARA
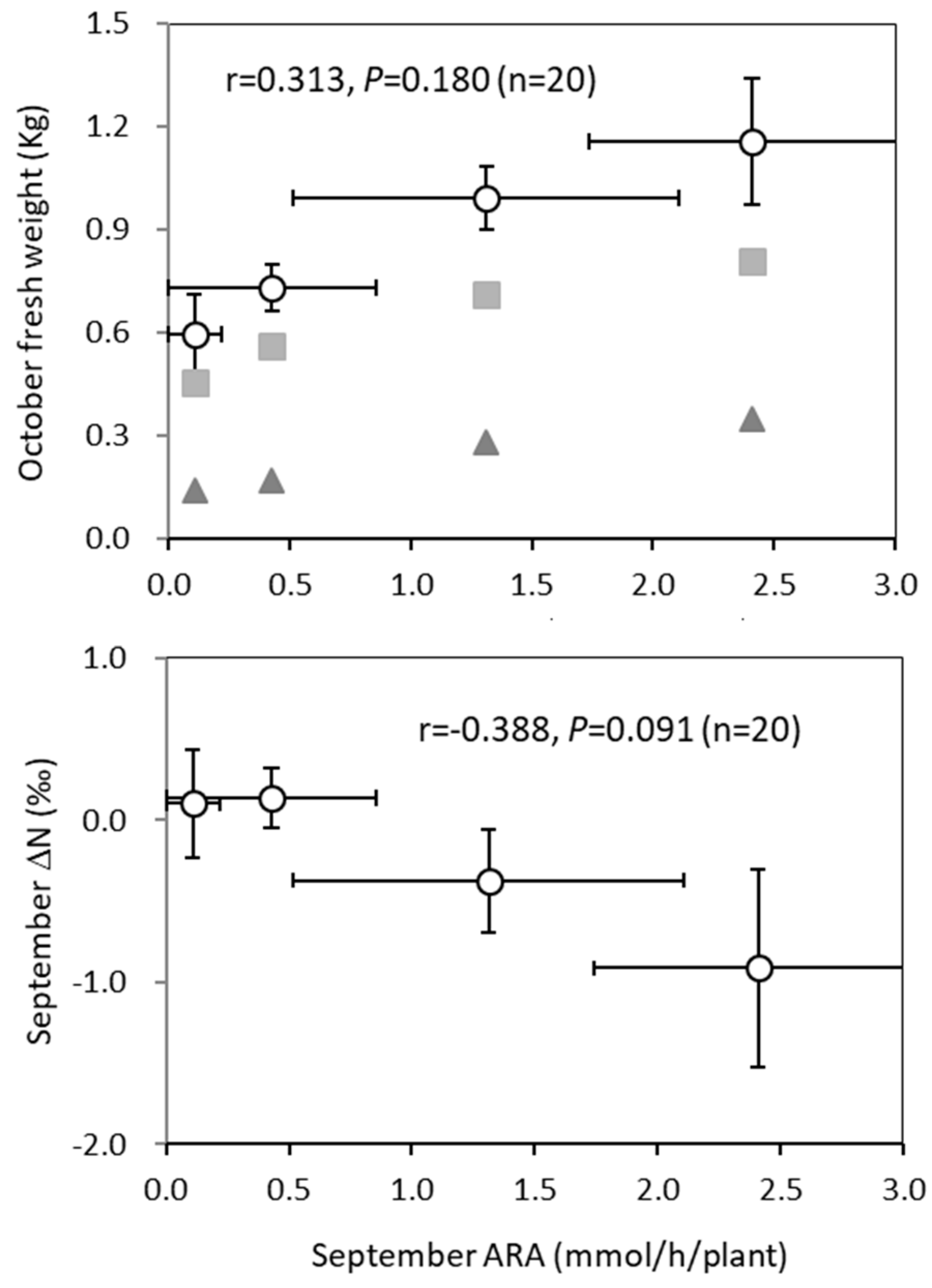
3.3. Relationship of ARA with Plant Growth and ∆15N Content
3.4. nifH Genes in Sweet Potato
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hill, W.A.; Dodo, H.; Hahn, S.K.; Mulongoy, K.; Adeyeye, S.O. Sweet potato root and biomass production with and without nitrogen fertilization. Agron. J. 1990, 82, 1120–1122. [Google Scholar] [CrossRef]
- Yonebayashi, K.; Katsumi, N.; Nishi, T.; Okazaki, M. Activation of nitrogen-fixing endophytes is associated with the tuber growth of sweet potato. Mass Spectrom. 2014, 3, A0032. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, T.; Terakado, J.; Masuda, T. Natural abundance of 15N in sweet potato, pumpkin, sorghum and castor bean: Possible input of N2-derived nitrogen in sweet potato. Biol. Fertil. Soils 1998, 26, 152–154. [Google Scholar] [CrossRef]
- Hill, W.A.; Bacon-Hill, P.; Crossman, S.M.; Stevens, C. Characterization of N2-fixing bacteria associated with sweet potato roots. Can. J. Microbiol. 1983, 29, 860–862. [Google Scholar] [CrossRef]
- Paula, M.A.; Reis, V.M.; Döbereiner, J. Interactions of Glomus clarum with Acetobacter diazotrophicus in infection of sweet potato (Ipomoea batatas), sugarcane (Saccharum spp.), and sweet sorghum (Sorghum vulgare). Biol. Fertil. Soils 1991, 11, 111–115. [Google Scholar] [CrossRef]
- Adachi, K.; Nakatani, M.; Mochida, H. Isolation of an endophytic diazotroph, Klebsiella oxytoca, from sweetpotato stems in Japan. Soil Sci. Plant Nutr. 2002, 48, 889–895. [Google Scholar] [CrossRef]
- Asis, C.A., Jr.; Adachi, K. Isolation of endophytic diazotroph Pantoea agglomerans and nondiazotroph Enterobacter asburiae from sweetpotato stem in Japan. Lett. Appl. Microbiol. 2003, 38, 19–23. [Google Scholar] [CrossRef]
- Terakado-Tonooka, J.; Fujihara, S.; Ohwaki, Y. Possible contribution of Bradyrhizobium on nitrogen fixation in sweet potatoes. Plant Soil 2013, 367, 639–650. [Google Scholar] [CrossRef]
- Khan, Z.; Doty, S.L. Characterization of bacterial endophytes of sweet potato plants. Plant Soil 2009, 322, 197–207. [Google Scholar] [CrossRef]
- Marques, J.M.; da Silva, T.F.; Vollú, R.E.; de Lacerda, J.R.M.; Blank, A.F.; Smalla, K.; Seldin, L. Bacterial endophytes of sweet potato tuberous roots affected by the plant genotype and growth stage. Appl. Soil Ecol. 2015, 96, 273–281. [Google Scholar] [CrossRef]
- Reiter, B.; Bürgmann, H.; Burg, K.; Sessitsch, A. Endophytic nifH gene diversity in African sweet potato. Can. J. Microbiol. 2003, 49, 549–555. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Aizaki, M.; Sumita, A. Effect of hydrophytes on the control of water temperature in model wetland type green roof garden. Environ. Sci. 2005, 18, 535–540. [Google Scholar]
- Poly, F.; Monrozier, L.J.; Bally, R. Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Res. Microbiol. 2001, 152, 95–103. [Google Scholar] [CrossRef]
- Adhikari, D.; Kaneto, M.; Itoh, K.; Suyama, K.; Pokharel, B.B.; Gaihre, Y.K. Genetic diversity of soybean-nodulation rhizobia in Nepal in relation to climate and soil properties. Plant Soil 2012, 357, 131–145. [Google Scholar] [CrossRef]
- Marques, J.M.; da Silva, T.F.; Vollu, R.E.; Blank, A.F.; Ding, G.C.; Seldin, L.; Smalla, K. Plant age and genotype affect the bacterial community composition in the tuber rhizosphere of field-grown sweet potato plants. FEMS Microbiol. Ecol. 2014, 88, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Terakado-Tonooka, J.; Ohwaki, Y.; Yamakawa, H.; Tanaka, F.; Yoneyama, T.; Fujihara, S. Expressed nifH genes of endophytic bacteria detected in field-grown sweet potatoes (Ipomoea batatas L.). Microbes Environ. 2008, 23, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Li, O.; Xiao, R.; Sun, L.; Guan, C.; Kong, D.; Hu, X. Bacterial and diazotrophic diversities of endophytes in Dendrobium catenatum determined through barcoded pyrosequencing. PLoS ONE 2017, 12, e0184717. [Google Scholar] [CrossRef] [PubMed]
- Soni, R.; Suyal, D.C.; Sai, S.; Goel, R. Exploration of nifH gene through soil metagenomes of the western Indian Himalayas. 3 Biotech 2016, 6, 25. [Google Scholar] [CrossRef]
- Samson, R.; Legendre, J.B.; Christen, R.; Fischer-Le, S.M.; Achouak, W.; Gardan, L. Transfer of Pectobacterium chrysanthemi (Burkholder et al. 1953) Brenner et al. 1973 and Brenneria paradisiaca to the genus Dickeya gen. nov. as Dickeya chrysanthemi comb. nov. and Dickeya paradisiaca comb. nov. and delineation of four novel species, Dickeya dadantii sp. nov., Dickeya dianthicola sp. nov., Dickeya dieffenbachiae sp. nov. and Dickeya zeae sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 1415–1427. [Google Scholar]
- Czajkowski, R.; Pérombelon, M.; Jafra, S.; Lojkowska, E.; Potrykus, M.; van der Wolf, J.; Sledz, W. Detection, identification and differentiation of Pectobacterium and Dickeya species causing potato blackleg and tuber soft rot: A review. Ann. Appl. Biol. 2015, 166, 18–38. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef]
- Gaby, J.C.; Buckley, D.H. A comprehensive evaluation of PCR primers to amplify the nifH gene of nitrogenase. PLoS ONE 2012, 7, e42149. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; Elsas, J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo-and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Latitude (°N) | Longitude (°E) | Temperature (°C) | Rainfall (mm) | Soil Type | Total-N (g/Kg) | P2O5 (mg/Kg) | K2O (mg/Kg) |
|---|---|---|---|---|---|---|---|---|
| Saitama | 36.04 | 139.35 | 12.5 a | 748 a | Andisol b | 10.6 | 2427 | 2142 |
| Shimane | 35.33 | 132.67 | 12.5 | 470 | Entisol | na c | na | na |
| Miyazaki | 32.12 | 131.48 | 15.8 | 1635 | Andisol | 4.16 | 189 | 167 |
| Kagoshima | 30.42 | 130.89 | 15.5 | 1306 | Andisol | 6.55 | 3078 | 1096 |
| PCR | SEQ | Dominant SEQ | Enterobacteriaceae | Dickeya | Klebsiella | Phytobacter | |
|---|---|---|---|---|---|---|---|
| Sa-P | 75 | 60 (80) z | 60 (100) y | 60 (100) x | 55 (92) w | 0 (0) w | 0 (0) w |
| Sa-Q | 27 | 27 (100) | 27 (100) | 20 (75) | 20 (100) | 0 (0) | 0 (0) |
| Sa-B | 80 | 70 (88) | 70 (100) | 70 (100) | 50 (71) | 5 (7) | 10 (14) |
| Ka-B | 50 | 45 (90) | 40 (89) | 35 (88) | 15 (43) | 10 (29) | 5 (14) |
| Sh-B | 75 | 65 (87) | 50 (77) | 35 (70) | 10 (29) | 15 (43) | 10 (29) |
| Mi-B | 50 | 35 (70) | 15 (43) | 5 (33) | 0 (0) | 0 (0) | 5 (100) |
| June | 77 | 67 (87) | 57 (85) | 47 (82) | 33 (71) | 0 (0) | 10 (21) |
| July | 53 | 47 (88) | 37 (79) | 37 (100) | 23 (64) | 7 (18) | 3 (9) |
| September | 63 | 57 (89) | 50 (88) | 37 (73) | 23 (64) | 10 (27) | 3 (9) |
| October | 48 | 32 (67) | 32 (100) | 32 (100) | 20 (63) | 4 (13) | 4 (13) |
| SUM | 61 | 51 (84) | 44 (86) | 38 (86) | 25 (66) | 5 (14) | 5 (14) |
| Most Similar nifH Sequence | Accession | No. of Sample | Similarity (%) | Class | Family |
|---|---|---|---|---|---|
| Dickeya dadantii 3937 | CP001654 | 29 (57) z | 92–99 | γ-Proteobacteria | Enterobacteriaceae |
| Klebsiella sp. CRLS069a | FJ593757 | 6 (12) | 92–100 | γ-Proteobacteria | Enterobacteriaceae |
| Phytobacter ursingii CAV1151 | CP011602 | 6 (12) | 91–98 | γ-Proteobacteria | Enterobacteriaceae |
| Paenibacillus stellifer DSM 14472 | CP009286 | 3 (6) | 88–90 | Bacilli | Paenibacillaceae |
| Enterobactor sp. YL34S | HQ204265 | 2 (4) | 84–86 | γ-Proteobacteria | Enterobacteriaceae |
| Pantoea sp. P0352 | FJ593780 | 1 (2) | 91 | γ-Proteobacteria | Enterobacteriaceae |
| Azotobacter chroococcum CGMCC 1.178 | EU693338 | 1 (2) | 94 | γ-Proteobacteria | Pseudomonadaceae |
| Bradyrhizobium sp. SEMIA 5067 | KY247016 | 1 (2) | 95 | α-Proteobacteria | Bradyrhizobiaceae |
| Azohydromonas lata IAM 12665 | AB201627 | 1 (2) | 97 | β-Proteobacteria | Alcaligenaceae |
| Variovorax sp. HW608 | LT607803 | 1 (2) | 96 | β-Proteobacteria | Comamonadaceae |
| June | July | September | October | |
|---|---|---|---|---|
| Sa-P | Dickeya (3) z | Dickeya (2) | Dickeya (2) | Dickeya (4) |
| Pantoea (1) | ||||
| Sa-Q | Dickeya (1) | Dickeya (1) | Dickeya (1) | |
| Bradyrhizobium (1) | ||||
| Sa-B | Dickeya (3) | Dickeya (3) | Dickeya (3) | Dickeya (1) |
| Phytobacter (1) | Klebsiella (1) | Phytobacter (1) | Enterobactor (1) | |
| Ka-B | Dickeya (3) | Klebsiella (2) | Phytobacter (1) | |
| Enterobactor (1) | ||||
| Variovorax (1) | ||||
| Sh-B | Phytobacter (1) | Phytobacter (1) | Paenibacillus (2) | Klebsiella (1) |
| Azohydromonas (1) | Dickeya (1) | Dickeya (1) | ||
| Klebsiella (1) | Klebsiella (1) | |||
| Mi-B | Phytobacter (1) | Paenibacillus (1) | ||
| Azotobacter (1) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Itoh, K.; Ohashi, K.; Yakai, N.; Adachi, F.; Hayashi, S. Changes in Acetylene Reduction Activities and nifH Genes Associated with Field-Grown Sweet Potatoes with Different Nursery Farmers and Cultivars. Horticulturae 2019, 5, 53. https://doi.org/10.3390/horticulturae5030053
Itoh K, Ohashi K, Yakai N, Adachi F, Hayashi S. Changes in Acetylene Reduction Activities and nifH Genes Associated with Field-Grown Sweet Potatoes with Different Nursery Farmers and Cultivars. Horticulturae. 2019; 5(3):53. https://doi.org/10.3390/horticulturae5030053
Chicago/Turabian StyleItoh, Kazuhito, Keisuke Ohashi, Nao Yakai, Fumihiko Adachi, and Shohei Hayashi. 2019. "Changes in Acetylene Reduction Activities and nifH Genes Associated with Field-Grown Sweet Potatoes with Different Nursery Farmers and Cultivars" Horticulturae 5, no. 3: 53. https://doi.org/10.3390/horticulturae5030053
APA StyleItoh, K., Ohashi, K., Yakai, N., Adachi, F., & Hayashi, S. (2019). Changes in Acetylene Reduction Activities and nifH Genes Associated with Field-Grown Sweet Potatoes with Different Nursery Farmers and Cultivars. Horticulturae, 5(3), 53. https://doi.org/10.3390/horticulturae5030053