A Dwarf Phenotype Identified in Breadfruit (Artocarpus altilis) Plants Growing on Marang (A. odoratissimus) Rootstocks


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Morphology Characterisation of Grafted Breadfruit Plants
2.3. Determination of Total Chlorophyll Content in Leaves
2.4. Determination of Total Soluble Sugar in Leaves
2.5. Measurement of Plasma Membrane H+-ATPase Activity in Stems
2.6. Statistical Analyses
3. Results
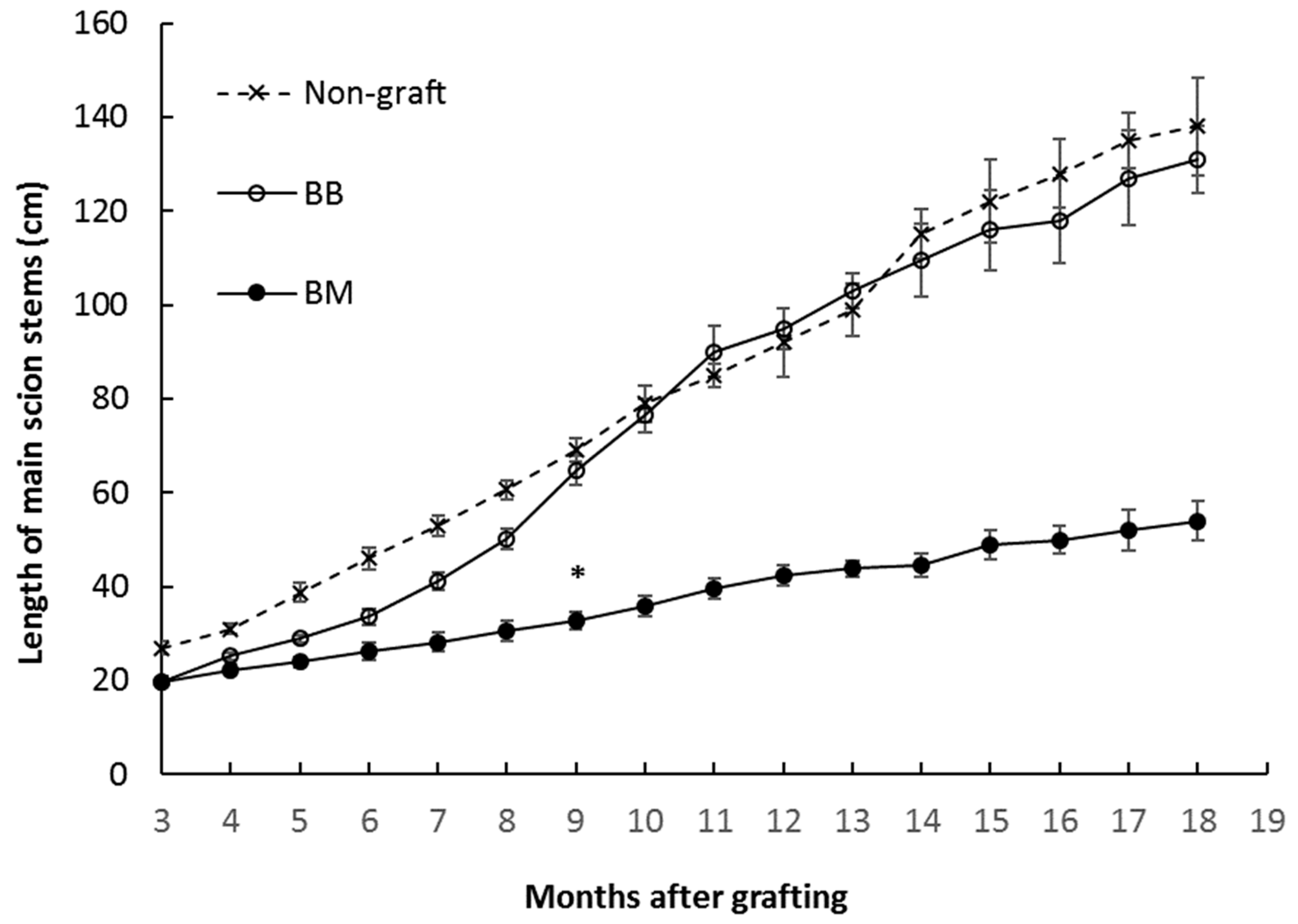
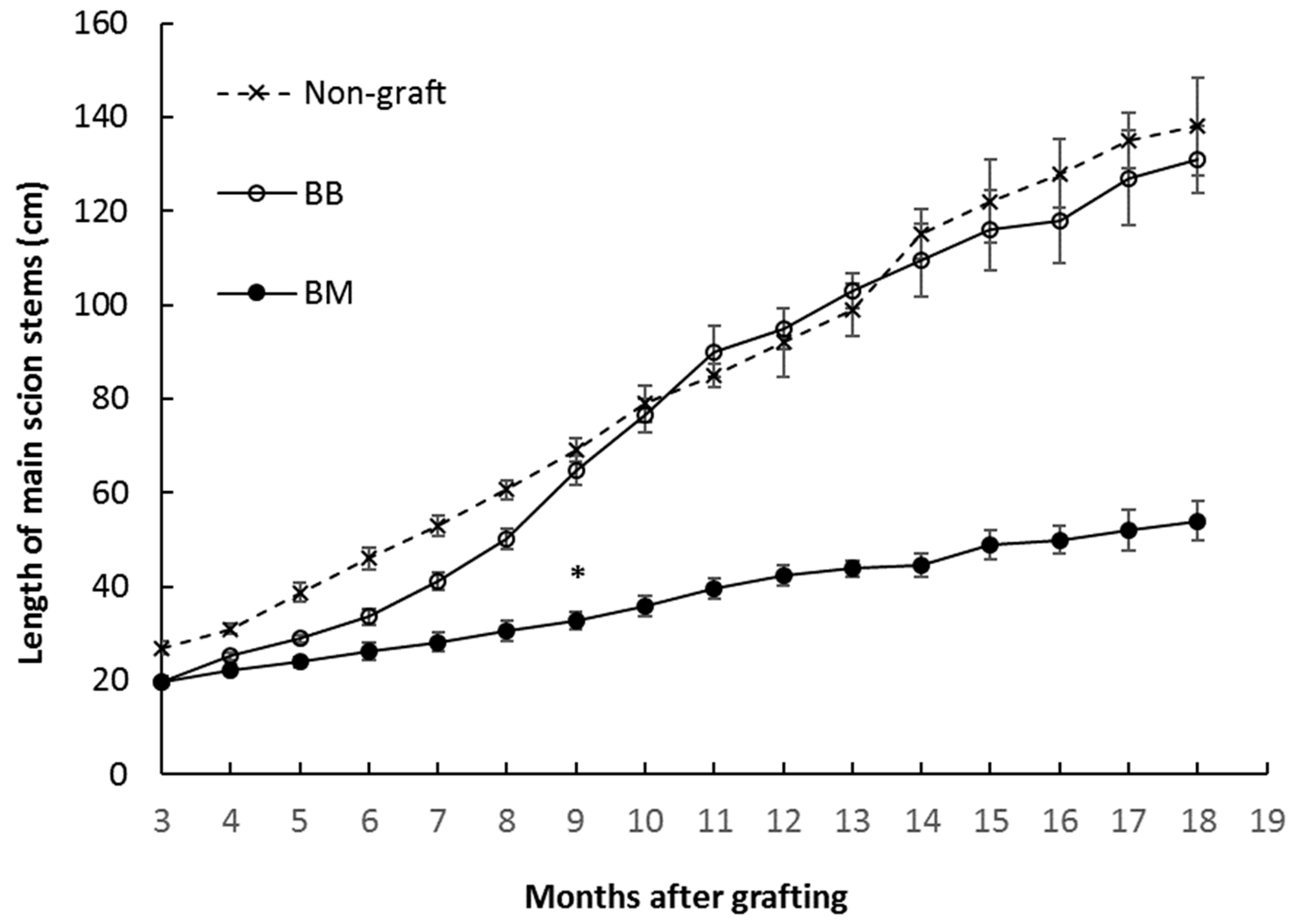
3.1. Growth Analysis of Breadfruit Plants Growing on Different Rootstocks
3.2. Effect of Rootstocks on the Biochemical Properties of Scion Leaves
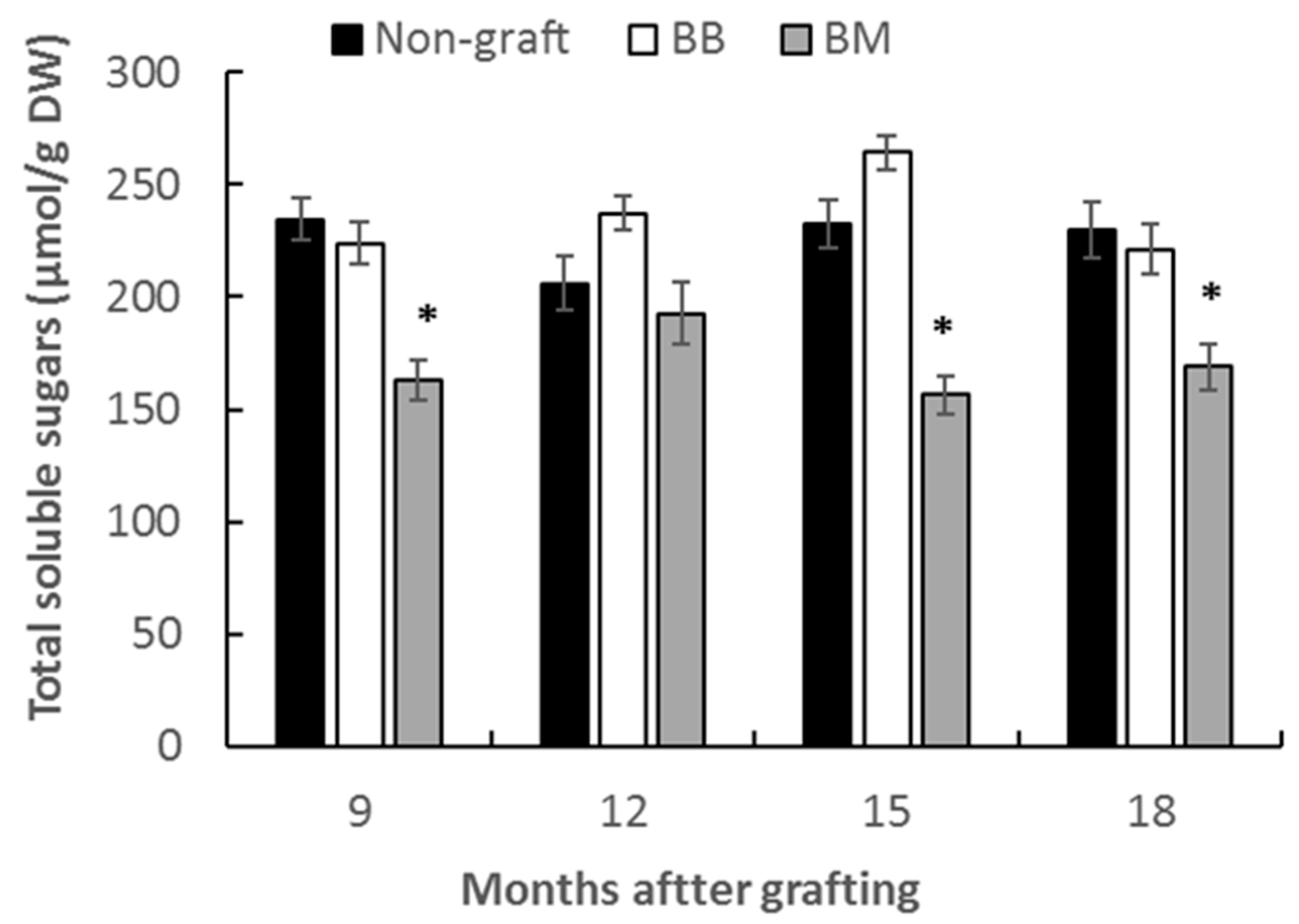
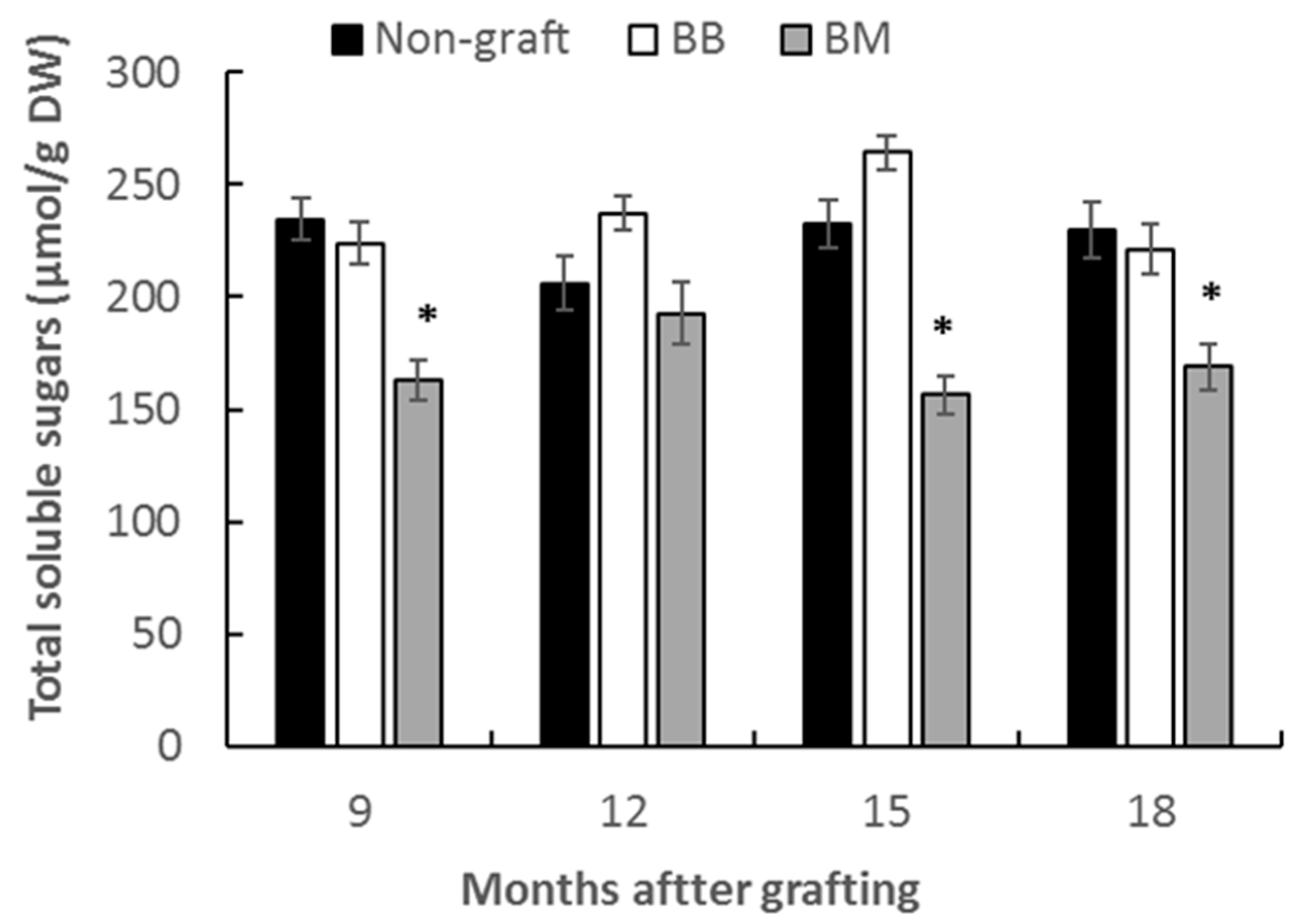
3.2.1. Total Soluble Sugar Content
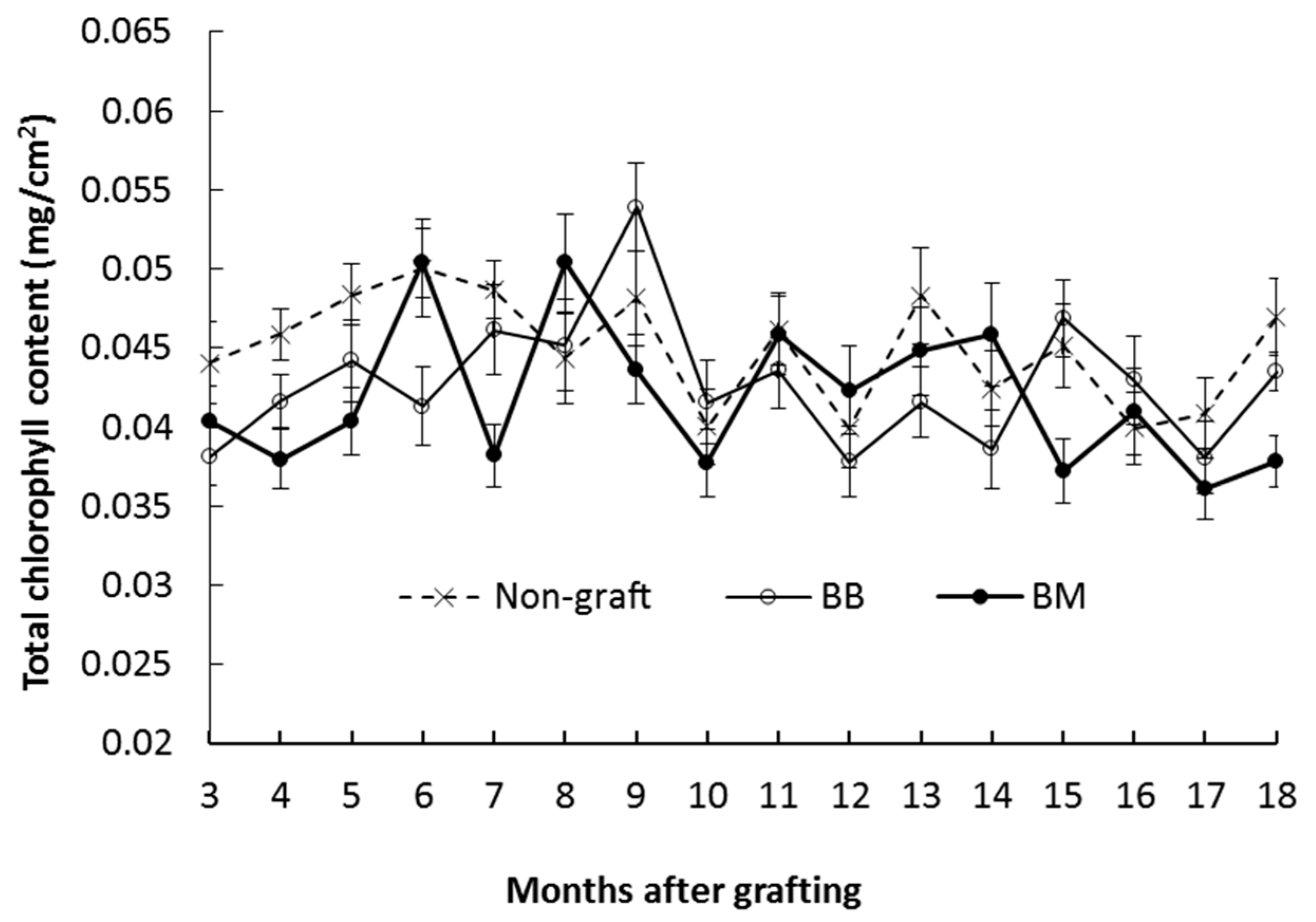
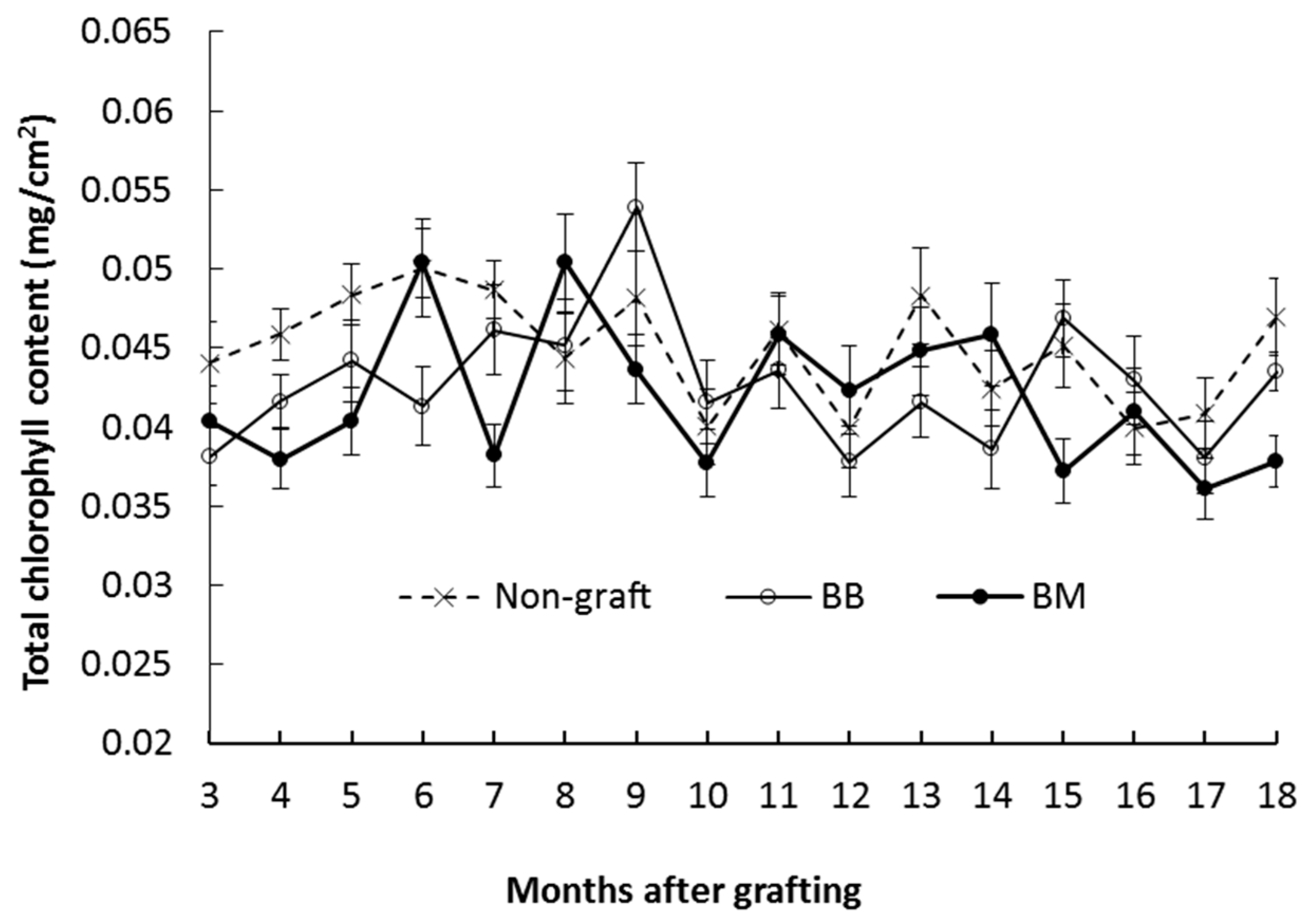
3.2.2. Total Chlorophyll Content
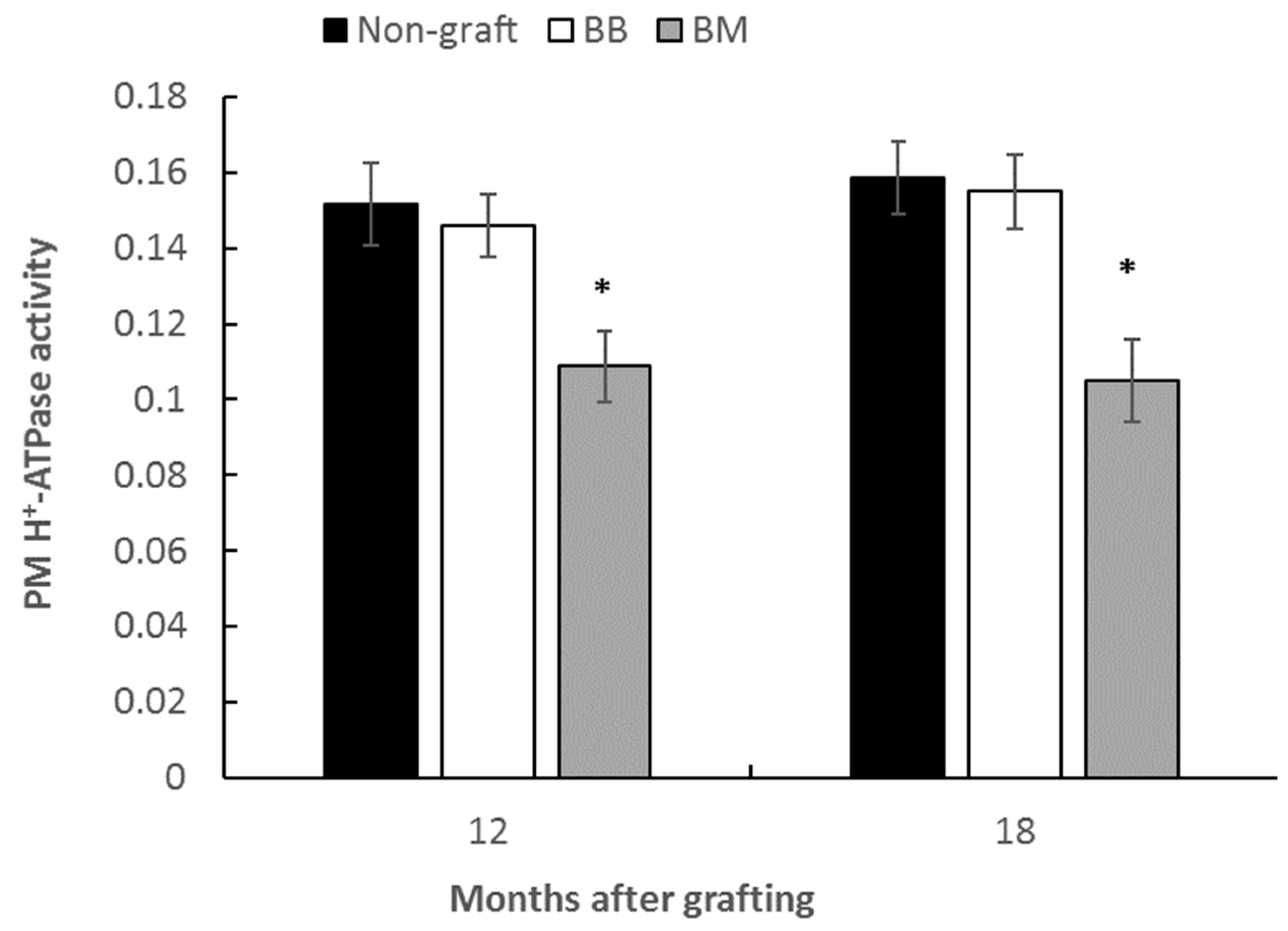
3.3. Effect of Rootstocks on the Plasma Membrane ATPase Activity of Stems
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ragone, D. Breadfruit, Artocarpus Altilis (Parkinson) Fosberg-Promoting the Conservation and Use of Underutilized and Neglected Crops. 10; Leibniz Institute of Plant Genetics and Crop Plant Research (IPK): Gatersleben, Germany, 1997; pp. 1–77. [Google Scholar]
- Daley, O.; Gloster, M.; Roberts-Nkrumah, L.B. Assessment and characterization of damage by hurricane Tomas to major tree crops with special emphasis on breadfruit (Artocarpus altilis) and breadnut (Artocarpus camansi) in St. Lucia and St. Vincent and the Grenadines. Proc. Caribb. Food Crop Soc. 2012, 48, 124–131. [Google Scholar]
- Zhou, Y.; Underhill, S.J.R. Plasma membrane H-ATPase activity and graft success of breadfruit (Artocarpus altilis) onto interspecific rootstocks of marang (A. odoratissimus) and pedalai (A. sericicarpus). Plant Biol. 2018, 20, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Taylor, M.B.; Underhill, S.J.R. Dwarfing of breadfruit (Artocarpus altilis) trees: Opportunities and challenges. Am. J. Exp. Agric. 2014, 4, 1743–1763. [Google Scholar] [CrossRef]
- Roberts-Nkrumah, L.B. Breadnut and Breadfruit Propagation, a Manual for Commercial Propagation; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012; pp. 1–18. [Google Scholar]
- Jones, A.M.P.; Ragone, D.; Tavana, N.G.; Bernotas, D.W.; Murch, S.J. Beyond the bounty: Breadfruit (Artocarpus altilis) for food security and novel foods in the 21st century. Ethnobot. Res. Appl. 2011, 9, 129–149. [Google Scholar] [CrossRef]
- Marini, R.P.; Fazio, G. Apple rootstocks: History, physiology, management, and breeding. Hortic. Rev. 2017, 45, 197–312. [Google Scholar]
- Nimbolkar, P.K.; Awachare, C.; Reddy, Y.T.N.; Chander, S.; Hussain, F. Role of Rootstocks in Fruit Production—A Review. J. Agric. Eng. Food Technol. 2016, 3, 183–188. [Google Scholar]
- Cheng, F.S.; Roose, M.L. Origin and inheritance of dwarfing by the citrus rootstock poncirus trifoliata ‘Flying Dragon’. J. Am. Soc. Hortic. Sci. 1995, 120, 286–291. [Google Scholar] [CrossRef]
- Haq, M.Z.; Robbani, M.; Ali, M.; Hasan, M.M.; Hasan, M.M.; Uddin, M.J.; Begum, M.; da Silva, J.A.T.; Pan, X.-Y.; Karim, M.R. Damage and management of cyclone Sidr-affected homestead tree plantations: A case study from Patuakhali, Bangladesh. Nat. Hazards 2012, 64, 1305–1322. [Google Scholar] [CrossRef]
- Parthiban, K.T.; Krishnakumar, N.; Karthick, N. (Eds.) Introduction to Forestry & Agroforestry; Scientific Publishers: Jodhpur, India, 2018; p. 367. [Google Scholar]
- Calvert, G. An Assessment of Tree Susceptibility and Resistance to Cyclones—with Particular Reference to Severe Tropical Cyclone Yasi; Greening Australia, Townsville City Council and Ergon Energy: Queensland, Australia, 2011; Available online: https://www.greeningaustralia.org.au/wp-content/uploads/2017/11/RESEARCH_Yasi_TreeReport_NewFormat.pdf (accessed on 15 January 2019).
- Tropicos. Available online: http://www.tropicos.org/NameSearch.aspx (accessed on 15 January 2019).
- Nandwani, D.; Kuniyuki, A.H. Grafting and improvement of breadfruit production in micronesia. In Proceedings of the International Symposium on Harnessing the Potential of Horticulture in the Asian-Pacific Region, Coolum, Australia, 1–3 September 2005; Volume 694, pp. 307–310. [Google Scholar]
- Medagoda, I.; Chandrarathna, W.M. Grafting of breadfruit (Artocarpus altilis) using breadnut (Artocarpus camansi) as root stock. Acta Hortic. 2007, 757, 149–152. [Google Scholar] [CrossRef]
- Solomon, F.J.; Roberts-Nkrumah, L.B. An evaluation of factors influencing successful grafting of breadfruit on chataigne rootstock. Proc. Caribb. Food Crop. Soc. 2008, 44, 304–312. [Google Scholar]
- Palmgren, M.G. Plant plasma membrane H+-ATPase: Powerhouses for nutrient uptake. Annu. Rev. Plant Biol. 2001, 52, 817–845. [Google Scholar] [CrossRef] [PubMed]
- Goebel, R. Breadfruit-the Australian scene. In Proceedings of the International Symposium on Breadfruit Research and Development, Nadi, Fiji, 16–19 April 2007; Volume 757, pp. 141–148. [Google Scholar]
- Goebel, R.; Breadfruit. Peninsula Gardening Notes, Department of Primary Industries and Fisheries, Queensland. 2004. Available online: http://plant.daleysfruit.com.au/l/breadfruit-tress-677.pdf (accessed on 15 January 2019).
- Richardson, A.D.; Duigan, S.P.; Berlyn, G.P. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol. 2002, 153, 185–194. [Google Scholar] [CrossRef]
- Yemm, E.; Willis, A. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1956, 57, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Webster, A.D. Rootstock and interstock effects on deciduous fruit tree vigour, precocity, and yield productivity. N. Z. J. Crop Hortic. Sci. 1995, 23, 373–382. [Google Scholar] [CrossRef]
- Forner-Giner, M.A.; Rodriguez-Gamir, J.; Martinez-Alcantara, B.; Quinones, A.; Iglesias, D.J.; Primo-Millo, E.; Forner, J. Performance of Navel orange trees grafted onto two new dwarfing rootstocks (Forner-Alcaide 517 and Forner-Alcaide 418). Sci. Hortic. 2014, 179, 376–387. [Google Scholar] [CrossRef]
- Clearwater, M.; Seleznyova, A.; Thorp, T.; Blattmann, P.; Barnett, A.; Lowe, R.; Austin, P. Vigor-controlling rootstocks affect early shoot growth and leaf area development of kiwifruit. Tree Physiol. 2006, 26, 505–515. [Google Scholar] [CrossRef]
- Seleznyova, A.N.; Thorp, T.G.; White, M.; Tustin, S.; Costes, E. Application of architectural analysis and AMAPmod methodology to study dwarfing phenomenon: The branch structure of ‘Royal Gala’ apple grafted on dwarfing and non-dwarfing rootstock/interstock combinations. Ann. Bot. 2003, 91, 665–672. [Google Scholar] [CrossRef]
- Lochard, R.G.; Schneider, G.W. Stock and scion growth relationships and the dwarfing mechanism in apple. Hortic. Rev. 1981, 3, 315–375. [Google Scholar]
- Salvatierra, M.A.; Gemma, H.; Iwahori, S. Partitioning of Carbohydrates and Development of Tissues in the Graft Union of Peaches Grafted on Prunus tomentosa Thunb. Rootstock. J. Jpn. Soc. Hortic. Sci. 1998, 67, 175–182. [Google Scholar] [CrossRef]
- Foster, T.M.; McAtee, P.A.; Waite, C.N.; Boldingh, H.L.; McGhie, T.K. Apple dwarfing rootstocks exhibit an imbalance in carbohydrate allocation and reduced cell growth and metabolism. Hortic. Res. 2017, 4, 17009. [Google Scholar] [CrossRef]
- Brown, C.S.; Young, E.; Pharr, D.M. Rootstock and scion effects on the seasonal distribution of dry weight and carbohydrates in young apple trees. J. Am. Soc. Hortic. Sci. 1985, 110, 696–701. [Google Scholar]
- Smeekens, S.; Ma, J.; Hanson, J.; Rolland, F. Sugar signals and molecular networks controlling plant growth. Curr. Opin. Plant Biol. 2010, 13, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chan, K.; Wang, T.; Hedley, C.; Offler, C.; Patrick, J. Intracellular sucrose communicates metabolic demand to sucrose transporters in developing pea cotyledons. J. Exp. Bot. 2009, 60, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Roldán, M.; Gómez-Mena, C.; Ruiz-García, L.; Salinas, J.; Martínez-Zapater, J.M. Sucrose availability on the aerial part of the plant promotes morphogenesis and flowering of Arabidopsis in the dark. Plant J. 2002, 20, 581–590. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhu, J.-Y.; Roh, J.; Marchive, C.; Kim, S.-K.; Meyer, C.; Sun, Y.; Wang, W.; Wang, Z.-Y. TOR signaling promotes accumulation of BZR1 to balance growth with carbon availability in Arabidopsis. Curr. Biol. 2016, 26, 1854–1860. [Google Scholar] [CrossRef] [PubMed]
- Sveinsdottira, H.; Yan, F.; Zhu, Y.Y.; Peiter-Volk, T.; Schubert, S. Seed ageing-induced inhibition of germination and post-germination root growth is related to lower activity of plasma membrane H+-ATPase in maize roots. J. Plant Physiol. 2009, 166, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Okumura, M.; Inoue, S.; Kuwata, K.; Kinoshita, T. Photosynthesis Activates Plasma Membrane H+-ATPase via Sugar Accumulation. Plant Physiol. 2016, 171, 580–589. [Google Scholar] [CrossRef]
- Caesar, K.; Elgass, K.; Chen, Z.H.; Huppenberger, P.; Witthoft, J.; Schleifenbaum, F.; Blatt, M.R.; Oecking, C.; Harter, K. A fast brassinolide-regulated response pathway in the plasma membrane of Arabidopsis thaliana. Plant J. 2011, 66, 528–540. [Google Scholar] [CrossRef]
- Noda, K.; Okuda, H.; Iwagaki, I. Indole acetic acid and abscisic acid levels in new shoots and fibrous roots of citrus scion-rootstock combinations. Sci. Hortic. 2000, 84, 245–254. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Graft | Homograft | Marang Rootstock | |
|---|---|---|---|
| Final scion height (cm) * | 138.63 ± 10.43 b | 131.47 ± 11.02 b | 54.02 ± 7.34 a |
| Main stem diameter (cm) * | 2.73 ± 0.10 b | 2.69 ± 0.05 b | 1.80 ± 0.21 a |
| Node number on main stems * | 20.70 ± 3.32 a | 19.21 ± 2.37 a | 21.01 ± 2.46 a |
| Number of branches per plant * | 1.00 ± 0.32 b | 1.41 ± 0.24 b | 0.00 ± 0.00 a |
| Average growth rate in main stems (cm/month) | 6.00 ± 0.52 b | 5.70 ± 0.18 b | 2.41 ± 0.42 a |
| Average growth rate in branches (cm/month) | 4.89 ± 0.34 b | 6.11 ± 0.47 b | 0.00 ± 0.00 a |
| Average length of internodes (cm) | 2.98 ± 0.18 b | 3.20 ± 0.08 b | 1.81 ± 0.16 a |
| Final length of second internode (cm) * | 4.32 ± 0.39 b | 4.99 ± 0.71 b | 1.34 ± 0.21 a |
| leaf number per plant * | 6.20 ± 0.71 b | 7.20 ± 0.37 b | 3.51 ± 0.32 a |
| Leaf length * | 72.00 ± 4.40 b | 77.80 ± 3.01 b | 46.20 ± 4.23 a |
| Leaf width * | 56.60 ± 0.71 b | 60.00 ± 0.37 b | 35.75 ± 0.32 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Underhill, S.J.R. A Dwarf Phenotype Identified in Breadfruit (Artocarpus altilis) Plants Growing on Marang (A. odoratissimus) Rootstocks. Horticulturae 2019, 5, 40. https://doi.org/10.3390/horticulturae5020040
Zhou Y, Underhill SJR. A Dwarf Phenotype Identified in Breadfruit (Artocarpus altilis) Plants Growing on Marang (A. odoratissimus) Rootstocks. Horticulturae. 2019; 5(2):40. https://doi.org/10.3390/horticulturae5020040
Chicago/Turabian StyleZhou, Yuchan, and Steven J. R. Underhill. 2019. "A Dwarf Phenotype Identified in Breadfruit (Artocarpus altilis) Plants Growing on Marang (A. odoratissimus) Rootstocks" Horticulturae 5, no. 2: 40. https://doi.org/10.3390/horticulturae5020040
APA StyleZhou, Y., & Underhill, S. J. R. (2019). A Dwarf Phenotype Identified in Breadfruit (Artocarpus altilis) Plants Growing on Marang (A. odoratissimus) Rootstocks. Horticulturae, 5(2), 40. https://doi.org/10.3390/horticulturae5020040