
Monitoring of Fluorescence Characteristics of Satsuma Mandarin (Citrus unshiu Marc.) during the Maturation Period

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Sample Preparation
2.2.2. Peel and Flesh Extraction
2.2.3. Soluble Solid Content and Acidity Measurement
2.2.4. Fluorescence Spectrum Measurement
2.2.5. Data Processing
3. Results and Discussion
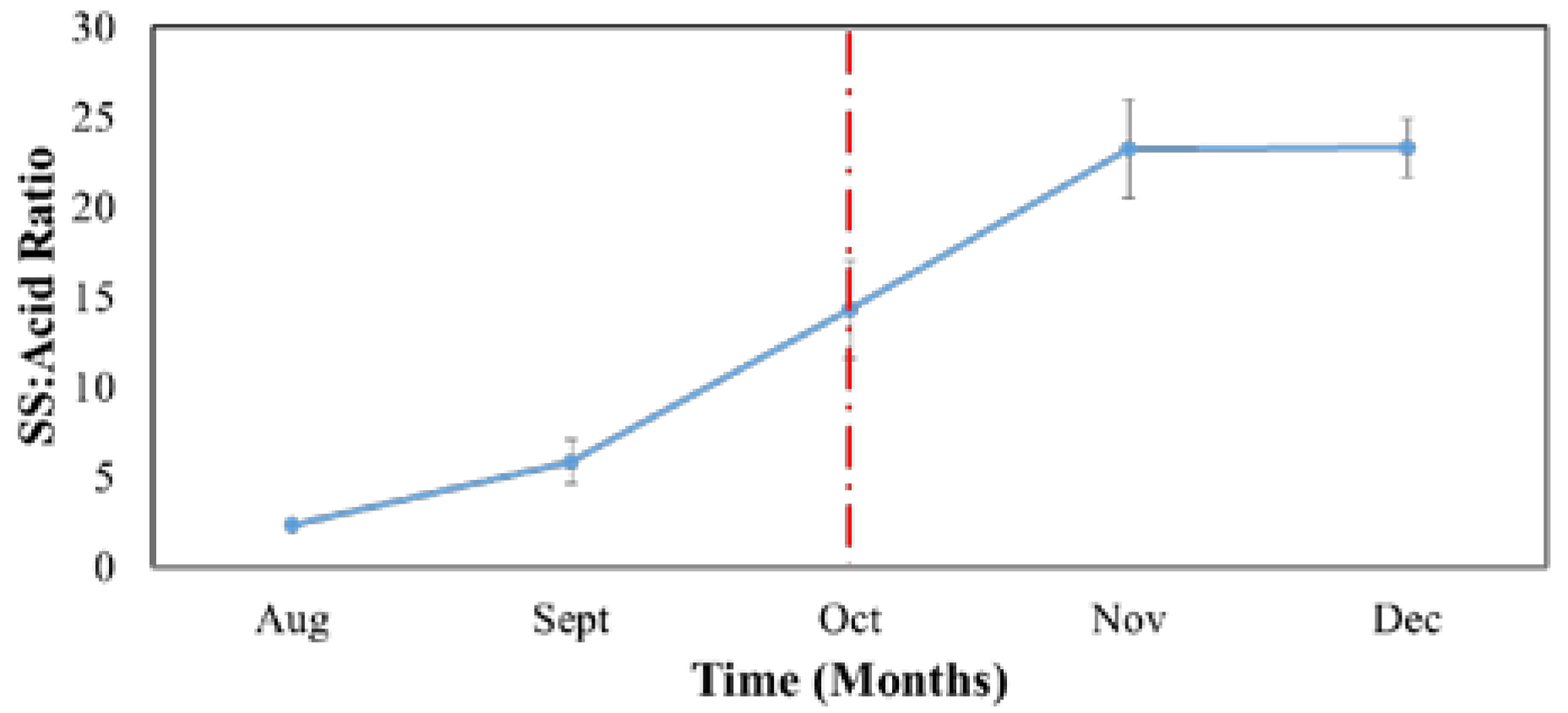
3.1. Fruit Growth and Development
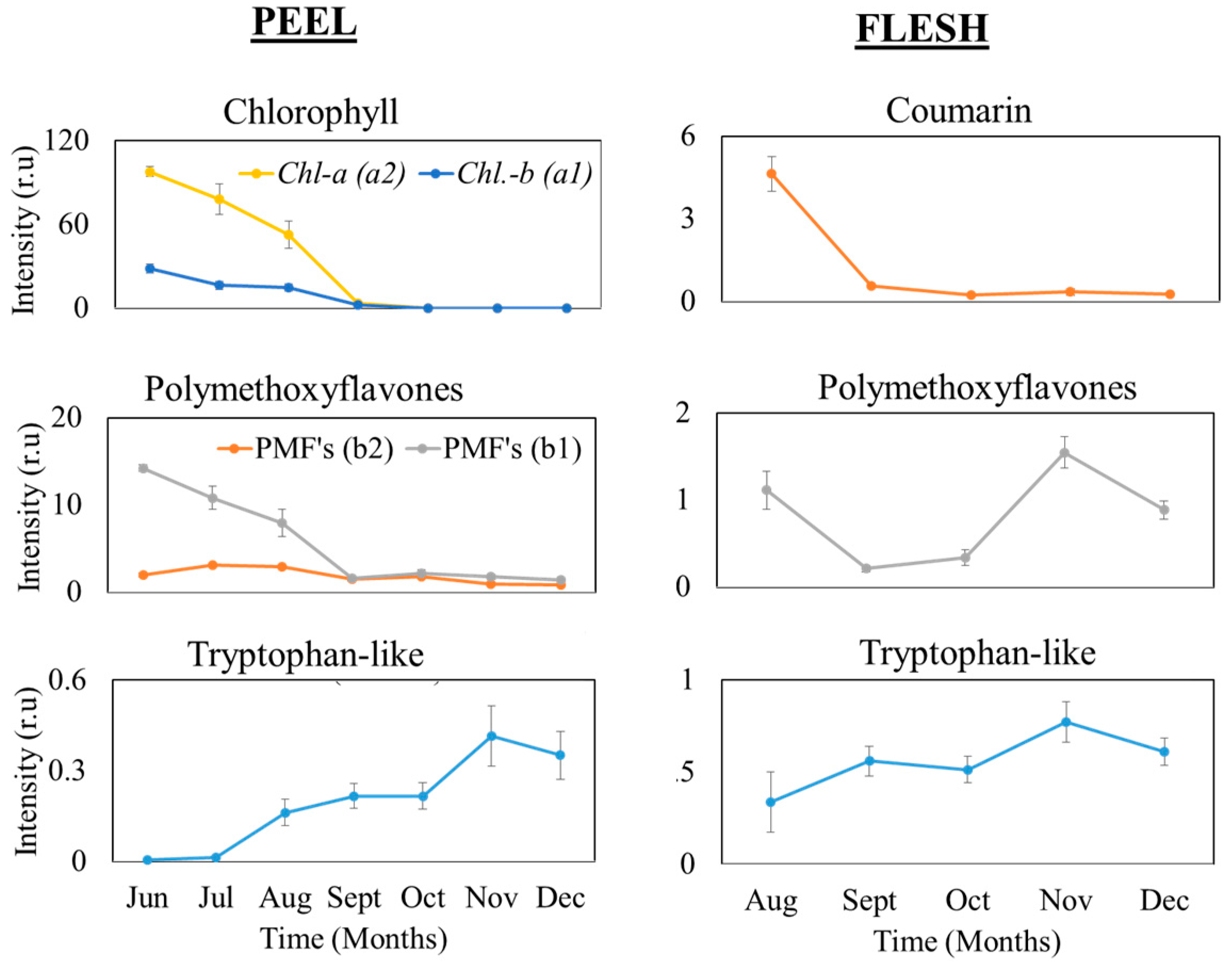
3.2. Suspected Compound in Satsuma Mandarin (unshiu) Excitation and Emission Matrices (EEM) during Maturing Period
3.3. Time Series Fluorescence during the Growth and Maturing Periods
3.4. Soluble Solid and Acid Metabolism during the Maturing Period
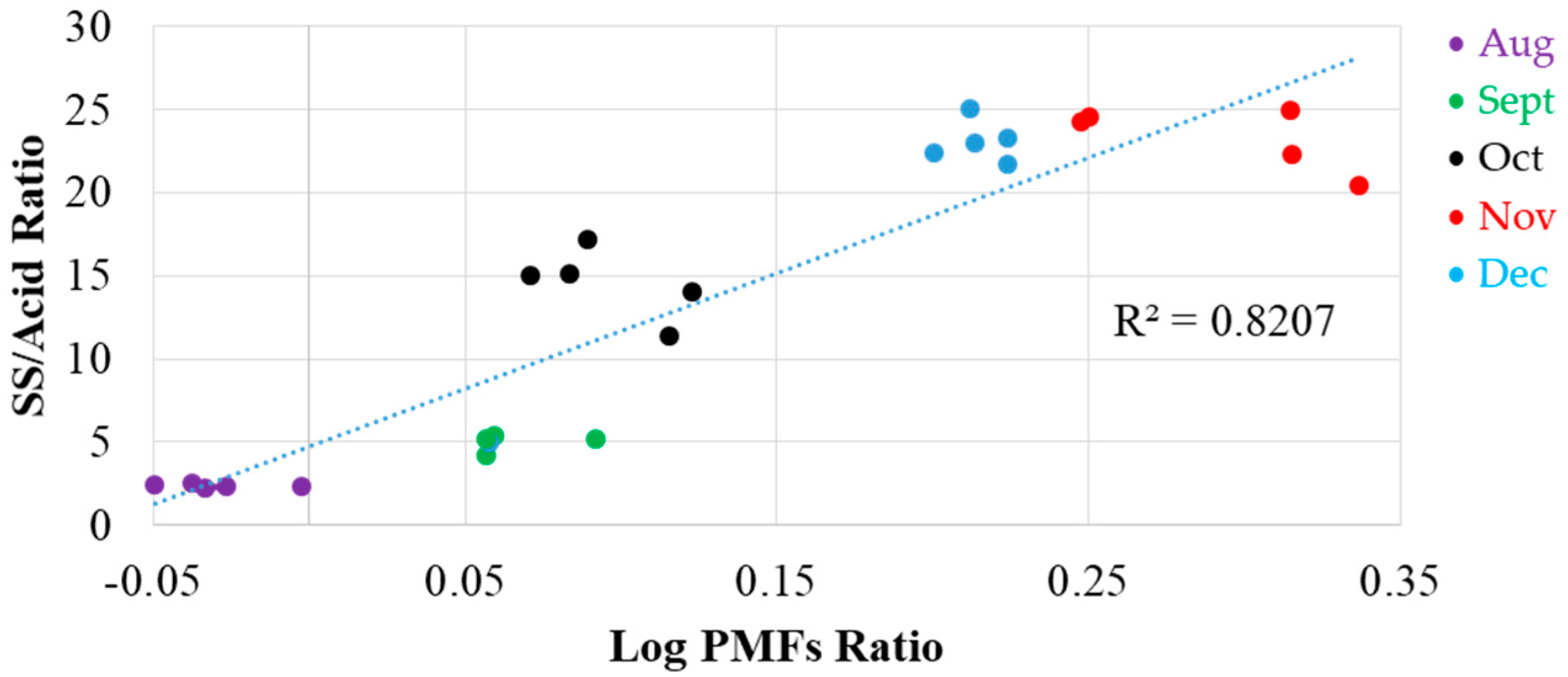
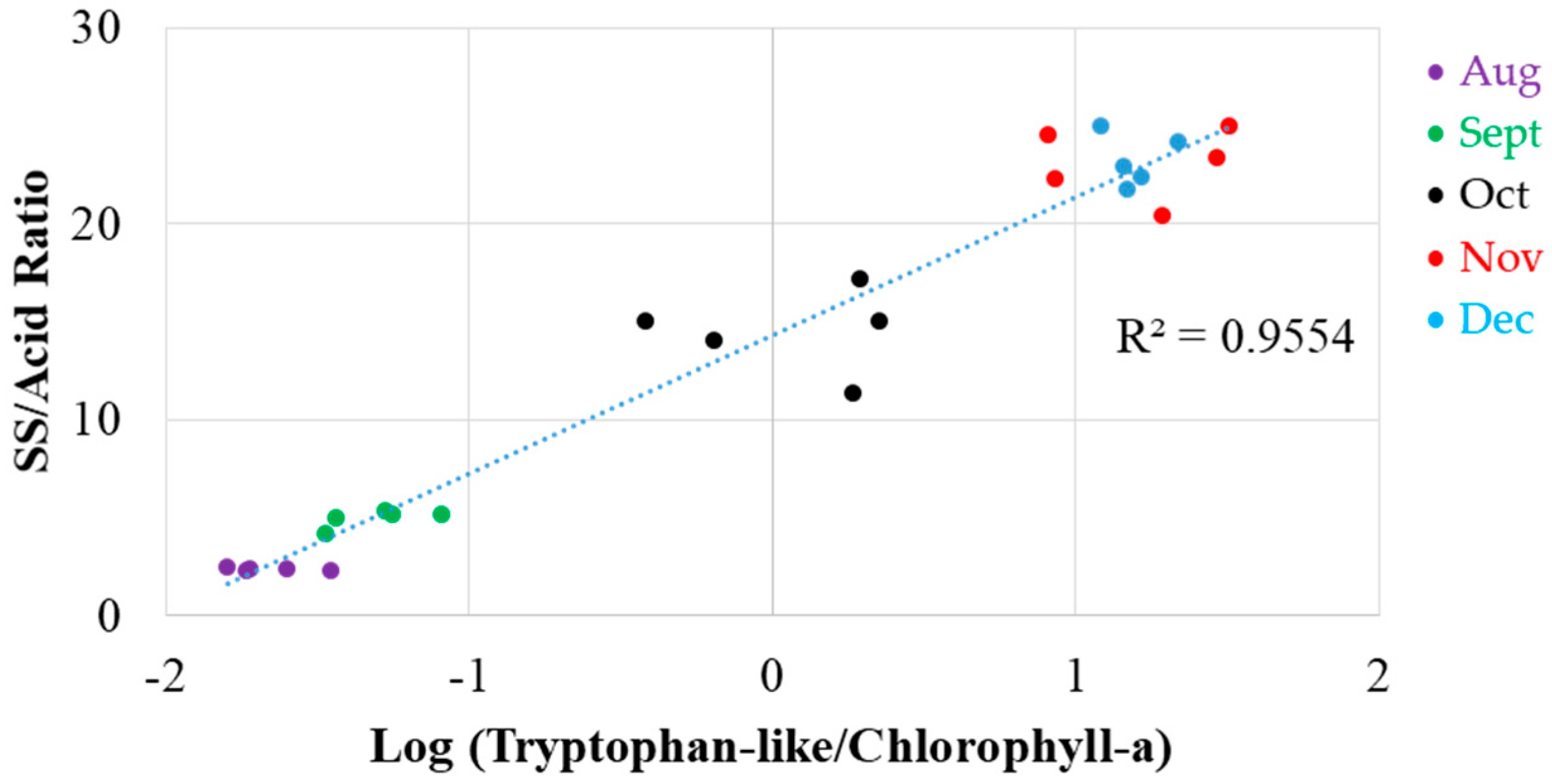
3.5. Potential Maturity Indices in Satsuma Mandarin
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kurai, T. 2016 Japan Citrus Annual; USDA Foreign Agricultural Service: Tokyo, Japan, 2016.
- Kader, A.A. Fruit maturity, ripening, and quality relationships. Acta Hortic. 1999, 57, 203–208. [Google Scholar] [CrossRef]
- Reid, M.S. Definition of Maturity. Postharvest Technol. Hortic. Crops 2002, 3311, 55. [Google Scholar]
- Cary, P.R. Citrus Fruit Maturity Vol-26; MPKV: Maharastra, India, 1974. [Google Scholar]
- Iglesias, D.J.; Cercós, M.; Colmenero-Flores, J.M.; Naranjo, M.A.; Ríos, G.; Carrera, E.; Ruiz-Rivero, O.; Lliso, I.; Morillon, R.; Tadeo, F.R.; et al. Physiology of citrus fruiting. Braz. J. Plant Physiol. 2007, 19, 333–362. [Google Scholar] [CrossRef]
- Crisosto, C.H. Stone fruit maturity indices: A descriptive. Postharvest News Inf. 1994, 5, 65N–68N. [Google Scholar]
- Gómez, A.H.; Wang, J.; Pereira, A.G. Mandarin ripeness monitoring and quality attribute evaluation using an electronic nose technique. Trans. ASABE 2007, 50, 2137–2142. [Google Scholar] [CrossRef]
- Yibin, Y.; Rao, X.; Ma, J. Methodology for Nondestructive Inspection of Citrus Maturity with Machine Vision–Transactions of The Chinese Society of Agricultural Engineering. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-NYGU200402034.htm (accessed on 6 April 2017).
- Antonucci, F.; Pallottino, F.; Paglia, G.; Palma, A.; D’Aquino, S.; Menesatti, P. Non-destructive estimation of mandarin maturity Status through portable VIS-NIR spectrophotometer. Food Bioprocess Technol. 2011, 4, 809–813. [Google Scholar] [CrossRef]
- Christensen, J.; Povlsen, V.T.; Sørensen, J. Application of Fluorescence Spectroscopy and Chemometrics in the Evaluation of Processed Cheese During Storage. J. Dairy Sci. 2003, 86, 1101–1107. [Google Scholar] [CrossRef]
- Momin, M.A.; Kondo, N.; Kuramoto, M.; Ogawa, Y.; Yamamoto, K.; Shiigi, T. Investigation of excitation wavelength for fluorescence emission of citrus peels based on UV-VIS spectra. Eng. Agric. Environ. Food 2012, 5, 126–132. [Google Scholar] [CrossRef]
- Betemps, D.L.; Fachinello, J.C.; Galarça, S.P.; Portela, N.M.; Remorini, D.; Massai, R.; Agati, G. Non-destructive evaluation of ripening and quality traits in apples using a multiparametric fluorescence sensor. J. Sci. Food Agric. 2012, 92, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Ghozlen, N.B.; Cerovic, Z.G.; Germain, C.; Toutain, S.; Latouche, G. Non-destructive optical monitoring of grape maturation by proximal sensing. Sensors 2010, 10, 10040–10068. [Google Scholar] [CrossRef] [PubMed]
- Song, H.S.; Lan Phi, N.T.; Park, Y.-H.; Sawamura, M. Volatile profiles in cold-pressed peel oil from korean and japanese shiranui (Citrus unshiu Marcov. × C. sinensis Osbeck × C. reticulata Blanco). Biosci. Biotechnol. Biochem 2006, 70, 737–739. [Google Scholar] [CrossRef] [PubMed]
- Seymour, G.B.; Taylor, J.E.; Tucker, G.A. Biochemistry of Fruit Ripening; Springer: Dordrecht, The Netherlands, 1993; ISBN 978-94-010-4689-3. [Google Scholar]
- Lawaetz, A.J.; Stedmon, C.A. Fluorescence intensity calibration using the raman scatter peak of water. Appl. Spectrosc. 2009, 63, 936–940. [Google Scholar] [CrossRef] [PubMed]
- Bain, J.M. Morphological, anatomical, and physiological changes in the developing fruit of the Valencia orange, Citrus sinensis (L.) Osbeck. Aust. J. Bot. 1958, 6, 1–23. [Google Scholar] [CrossRef]
- Igoshi, M.; Yamaguchi, I.; Takahashi, N.; Hirose, K. Plant growth substances in the young fruit of Citrus unshiu. Agric. Biol. Chem. 1971, 35, 629–631. [Google Scholar] [CrossRef]
- Takahashi, N.; Yamaguchi, I.; Kono, T.; Igoshi, M.; Hirose, K.; Suzuki, K. Characterization of plant growth substances in Citrus unshiu and their change in fruit development. Plant Cell Physiol. 1975, 16, 1101–1111. [Google Scholar] [CrossRef]
- Goren, R.; Monselise, S.P. Inter-relations of hesperidin, some other natural components and certain enzyme systems in developing shamouti orange fruits. J. Hortic. Sci. 1965, 40, 83–99. [Google Scholar] [CrossRef]
- Fujii, H.; Shimada, T.; Sugiyama, A.; Nishikawa, F.; Endo, T.; Nakano, M.; Ikoma, Y.; Shimizu, T.; Omura, M. Profiling ethylene-responsive genes in mature mandarin fruit using a citrus 22K oligoarray. Plant Sci. 2007, 173, 340–348. [Google Scholar] [CrossRef]
- Fidelibus, M.W.; Koch, K.E.; Davies, F.S. Gibberellic acid alters sucrose, hexoses, and their gradients in peel tissues during color break delay in “Hamlin” orange. J. Am. Soc. Hortic. Sci. 2008, 133, 760–767. [Google Scholar]
- Agustí, M.; Martinez-Fuentes, A.; Mesejo, C. Citrus fruit quality. Physiological basis and techniques of improvement. Agrociencia 2002, 6, 1–16. [Google Scholar]
- Ustin, S.L.; Gitelson, A.A.; Jacquemoud, S.; Schaepman, M.; Asner, G.P.; Gamon, J.A.; Zarco-Tejada, P. Retrieval of foliar information about plant pigment systems from high resolution spectroscopy. Remote Sens. Environ. 2009, 113, S67–S77. [Google Scholar] [CrossRef]
- Li, S.; Lo, C.-Y.; Ho, C.-T. Hydroxylated polymethoxyflavones and methylated flavonoids in sweet orange (Citrus sinensis) peel. J. Agric. Food Chem. 2006, 54, 4176–4185. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Iyi, N.; Hashizume, H.; Shimomura, S.; Ando, T. Preparation of integrated coumarin/cyanine systems within an interlayer of phyllosilicate and fluorescence resonance energy transfer. Chem. Mater. 2009, 21, 1179–1181. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, H.; Sun, Y.; Zhang, Y.; Liu, H.; Cheng, J.; Bi, S.; Zhang, H. Study of interaction between protein and main active components in Citrus aurantium L. by optical spectroscopy. J. Lumin. 2010, 130, 270–279. [Google Scholar] [CrossRef]
- El-Otmani, D.M.; Lovatt, C.J.; Jr, C.W.C.; Agustí, M. Plant growth regulators in citriculture: Factors regulating endogenous levels in Citrus tissues. Crit. Rev. Plant Sci. 1995, 14, 367–412. [Google Scholar] [CrossRef]
- Takagi, T.; Mukai, H.; Ichikawa, T.; Suzuki, T. Effects of temperature and sugar accumulation in fruits on color development of satsuma mandarin. J. Jpn. Soc. Hortic. Sci. 1994, 62, 725–731. [Google Scholar] [CrossRef]
- Veldhuis, M.K.; Swift, L.J.; Scott, W.C. Fully-methoxylated flavones in Florida orange juices. J. Agric. Food Chem. 1970, 18, 590–592. [Google Scholar] [CrossRef]
- Musialik, M.; Kuzmicz, R.; Pawłowski, T.S.; Litwinienko, G. Acidity of hydroxyl groups: An overlooked Influence on antiradical properties of flavonoids. J. Org. Chem. 2009, 74, 2699–2709. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; KrishnaRaj, S.; Saxena, P.K. Tryptophan is a precursor for melatonin and serotonin biosynthesis in in vitro regenerated St. John’s wort (Hypericum perforatum L. cv. Anthos) plants. Plant Cell Rep. 2000, 19, 698–704. [Google Scholar] [CrossRef]
- Radwanski, E.R.; Last, R.L. Tryptophan biosynthesis and metabolism: Biochemical and molecular genetics. Plant Cell 1995, 7, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.-C.; Parry, A.; Tena, M.; Jorrin, J.; Edwards, R. Abiotic elicitation of coumarin phytoalexins in sunflower. Phytochemistry 1995, 38, 1185–1191. [Google Scholar] [CrossRef]
- Ladaniya, M. Citrus Fruit: Biology, Technology and Evaluation; Academic Press: Croydon, UK, 2010; ISBN 978-0-08-055623-9. [Google Scholar]
- Thimann, K.V.; Bonner, W.D. Inhibition of plant growth by protoanemonin and coumarin, and its prevention by BAL. Proc. Natl. Acad. Sci. USA 1949, 35, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Olmo, M.; Nadas, A.; García, J.M. Nondestructive methods to evaluate maturity level of oranges. J. Food Sci. 2000, 65, 365–369. [Google Scholar] [CrossRef]








© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muharfiza; Al Riza, D.F.; Saito, Y.; Itakura, K.; Kohno, Y.; Suzuki, T.; Kuramoto, M.; Kondo, N. Monitoring of Fluorescence Characteristics of Satsuma Mandarin (Citrus unshiu Marc.) during the Maturation Period. Horticulturae 2017, 3, 51. https://doi.org/10.3390/horticulturae3040051
Muharfiza, Al Riza DF, Saito Y, Itakura K, Kohno Y, Suzuki T, Kuramoto M, Kondo N. Monitoring of Fluorescence Characteristics of Satsuma Mandarin (Citrus unshiu Marc.) during the Maturation Period. Horticulturae. 2017; 3(4):51. https://doi.org/10.3390/horticulturae3040051
Chicago/Turabian StyleMuharfiza, Dimas Firmanda Al Riza, Yoshito Saito, Kenta Itakura, Yasushi Kohno, Tetsuhito Suzuki, Makoto Kuramoto, and Naoshi Kondo. 2017. "Monitoring of Fluorescence Characteristics of Satsuma Mandarin (Citrus unshiu Marc.) during the Maturation Period" Horticulturae 3, no. 4: 51. https://doi.org/10.3390/horticulturae3040051
APA StyleMuharfiza, Al Riza, D. F., Saito, Y., Itakura, K., Kohno, Y., Suzuki, T., Kuramoto, M., & Kondo, N. (2017). Monitoring of Fluorescence Characteristics of Satsuma Mandarin (Citrus unshiu Marc.) during the Maturation Period. Horticulturae, 3(4), 51. https://doi.org/10.3390/horticulturae3040051