1. Introduction
Chrysanthemum (
Chrysanthemum ×
morifolium Ramat.) is one of the most economically important cut flowers worldwide and accounts for approximately 9.8% of the global cut flower trade, ranking second after roses based on 2022 trade statistics [
1]. In South Korea, chrysanthemum is also a major floricultural crop, occupying 296 ha of cultivation area in 2023 and accounting for the largest proportion among cut flower species [
2]. The total production value reached 26.37 million USD, ranking second among cut flower crops after roses.
Among commercial chrysanthemum cultivars in South Korea, ‘Baekgang’ has gained increasing attention as a promising standard-type cultivar known for its white rust resistance and extended vase life [
3]. Nevertheless, because ‘Baekgang’ is an autumn-flowering cultivar with a natural flowering period in October, forced cultivation during summer exposes plants to supra-optimal temperatures, which can inhibit vegetative growth and reduce flower quality.
Global surface temperature has increased rapidly in recent decades, and continued warming is expected to increase the frequency and intensity of extreme climate events, including severe heatwaves [
4]. In South Korea, the average summer temperature has also increased steadily in recent years [
5], and the annual mean temperature of the Korean Peninsula is projected to rise by 1.8 °C by 2040 and up to 7.0 °C by 2100 compared with current levels [
6]. These climatic conditions can be further intensified in protected cultivation systems. Environmental monitoring of a commercial hydroponic chrysanthemum greenhouse from July–August 2024 showed that root-zone and air temperatures exceeded 32 °C and 37 °C, respectively.
High temperatures during chrysanthemum cultivation can cause severe physiological disorders, including delayed flowering and abnormal inflorescence development [
7,
8,
9]. In particular, heat stress during reproductive development can disrupt normal capitulum formation, resulting in morphological abnormalities such as abnormal bract formation, irregular floret development, and open-center malformed capitula, ultimately reducing the ornamental quality and commercial value of cut flowers [
10,
11].
Maintaining optimal greenhouse temperatures during high-temperature seasons requires substantial installation and energy costs, making whole-greenhouse cooling economically challenging. As an alternative, localized root-zone cooling using chilled-water circulation has attracted attention as an energy-efficient strategy for mitigating heat stress in greenhouse crop production [
12,
13]. Previous studies have reported that localized temperature control can improve plant growth, flowering, and crop quality while reducing energy consumption compared with conventional whole-greenhouse cooling systems [
14,
15,
16].
Despite these advances, most previous studies on root-zone cooling have focused on vegetable crops or seedling production, whereas limited information is available for floricultural crops such as chrysanthemum under commercial summer cultivation conditions. In particular, the effects of root-zone temperature on open-center malformation and flower quality in standard chrysanthemum remain poorly understood. Moreover, few studies have evaluated root-zone cooling thresholds by simultaneously considering flower quality and energy consumption.
Therefore, this study aimed to investigate the effects of root-zone cooling during high-temperature summer cultivation on vegetative growth, flower quality, and open-center malformed capitula in standard chrysanthemum ‘Baekgang’. In addition, this study sought to identify a root-zone cooling threshold that balances flower quality and energy consumption and to evaluate the feasibility of root-zone cooling as an energy-saving environmental control strategy for protected chrysanthemum production.
2. Materials and Methods
2.1. Plant Materials and Growing Conditions
Rooted cuttings of standard chrysanthemum ‘Baekgang’ (Chrysanthemum × morifolium Ramat.) were used in this study. At transplanting, the cuttings had an initial plant height of 11.7 ± 0.9 cm and a stem diameter of 2.96 ± 0.25 mm and were planted at a density of approximately 111 plants m−2. Plants were transplanted on 30 May 2025 and grown for 38 d before the start of the cooling treatments. Root-zone cooling treatments were applied for 63 d, from 7 July to 8 September 2025, in a commercial hydroponic chrysanthemum greenhouse located in Gangseo-gu, Busan, South Korea (35.2148° N, 128.9380° E).
Plants were grown in custom-built hydroponic beds (40.0 m × 1.0 m × 0.2 m; L × W × D) filled with perlite (particle size, 2–5 mm). A nutrient solution was supplied 3–4 times daily through a drip irrigation system. The electrical conductivity (EC) was adjusted from 0.5 to 2.4 dS m−1 according to the growth stage, and the pH was maintained at 5.9.
To induce flowering, short-day treatment was initiated approximately 1 month after transplanting and maintained for 2 weeks by providing a dark period from 17:30 to 06:20. Following the short-day treatment, a four-day night-interruption treatment was applied uniformly to all treatments from 23:30 to 03:30 using white LED lamps (A60 type, 12 W, CRI 80; Hangzhou Yuzhong, Hangzhou, China). This temporary night-interruption treatment is commonly used in commercial chrysanthemum production to reduce physiological disorders associated with an abrupt transition from short-day to long-day photoperiodic conditions.
Environmental conditions were monitored using an integrated weather sensor (ATMOS 41, METER Group, Pullman, WA, USA). During the experimental period, the mean ambient temperature was 28.5 ± 3.3 °C, with daytime and nighttime means of 30.5 ± 2.9 °C and 26.8 ± 2.5 °C, respectively. The maximum and minimum air temperatures were 35.7 °C and 20.3 °C, respectively, and the mean relative humidity was 85.1 ± 11.3%. Changes in air temperature, relative humidity, and solar radiation during the experimental period are shown in
Figure 1.
2.2. Root-Zone Cooling System and Treatments
The experiment consisted of a conventional non-cooled control (NC) and three root-zone cooling treatments with different cooling activation thresholds. Each treatment was applied to a separate 40 m cultivation bed. Because the root-zone cooling treatments were imposed at the whole-bed level and only one commercial-scale bed was available for each treatment, potential bed effects and treatment effects could not be completely separated in the present experimental design. To characterize spatial variation within the commercial-scale beds, each bed was divided lengthwise into three equal sections (approximately 13.3 m each), which were used as within-bed spatial subsamples for plant sampling and root-zone temperature monitoring. These sections were not regarded as fully independent bed-level biological replicates.
Each cooling bed was equipped with a separate cooling unit consisting of a 100 L water tank, a chiller (DA-500B, Daeil, Busan, Republic of Korea), a circulation pump (PH-045M, Wilo, Busan, Republic of Korea), and a booster pump (PH-125W, Wilo, Republic of Korea). The three cooling systems operated independently, and chilled water was not shared among treatments. Four rows of 16-mm-diameter polyethylene pipes were installed parallel to the bed length and embedded at a depth of 10 cm below the substrate surface. This depth corresponded to the midpoint of the 20-cm-deep bed, and the four pipes were evenly arranged across the 1.0 m bed width to promote uniform distribution of root-zone cooling. The non-cooled control bed was managed conventionally by the grower without cooling pipes or chilled-water circulation.
The cooling treatments were defined by activation thresholds of 28 °C, 25 °C, and 22 °C, referred to as the high-temperature threshold treatment (HT), medium-temperature threshold treatment (MT), and low-temperature threshold treatment (LT), respectively. These thresholds were selected based on the reported favorable temperature range for chrysanthemum growth and development and on preliminary monitoring of summer root-zone temperature in the same commercial greenhouse. Because chrysanthemum growth and developmental responses have been reported to be favorable at approximately 18–22 °C, depending on cultivar [
17], the LT threshold of 22 °C was selected near the upper end of this favorable temperature range. The chilled-water temperature was set at 18 °C to provide sufficient cooling capacity to maintain the root zone near the assigned threshold. Based on preliminary monitoring during the previous summer, in which root-zone temperature exceeded 32 °C under commercial greenhouse conditions, the MT and HT thresholds of 25 °C and 28 °C were established as stepwise higher temperature exposure levels representing moderate and mild root-zone cooling, respectively. Chilled water was circulated automatically using digital temperature controllers (FOX-2001, Conotec, Busan, Republic of Korea) connected to independent control sensors (FS-200N, Conotec, Republic of Korea) installed at a depth of 10 cm in the root zone. Cooling was initiated when the root-zone temperature exceeded the assigned threshold by 0.1 °C and stopped when it decreased to 0.1 °C below the threshold.
Root-zone temperature was monitored using soil sensors (TEROS 12, METER Group, Pullman, WA, USA) installed at a depth of 10 cm. Data were recorded every 15 min using a ZL6 data logger (METER Group, Pullman, WA, USA). Power consumption was measured separately for each cooling treatment. The chiller, circulation pump, and booster pump of each cooling unit were connected to a dedicated power distributor, and cumulative electricity consumption was recorded using a smart plug (Tapo P110M, TP-Link, Irvine, CA, USA). Thus, the reported power consumption represents the total electricity use of each cooling unit and was expressed as kWh per bed during the experimental period. A schematic diagram of the root-zone cooling system is shown in
Figure 2.
2.3. Measurements of Growth and Photosynthetic Parameters
Growth parameters were measured at 11 d intervals beginning on 7 July 2025, which was defined as 0 days after treatment (DAT 0). At each sampling date, three plants were randomly selected from each section, resulting in nine plants per treatment. Plant height and stem diameter were measured before destructive harvest. After harvest, roots were gently washed under running tap water to remove perlite particles. Shoot and root fresh masses were measured, and shoot and root dry masses were determined after oven-drying at 80 °C for 72 h.
Photosynthetic parameters, including net CO2 assimilation rate (An), transpiration rate (E), and stomatal conductance (gs), were measured at 11 d intervals starting from 18 July 2025 (DAT 11) using a portable photosynthesis system (LI-6800, LI-COR, Lincoln, NE, USA). Measurements were conducted between 10:00 and 14:00 on five randomly selected plants per treatment using a 3 cm2 leaf chamber. To minimize the effect of diurnal variation, the order of treatment measurements was randomized within each measurement cycle. Gas-exchange parameters were recorded automatically at 1 min intervals. Leaves were acclimated to the chamber conditions for approximately 5–10 min until steady-state gas-exchange parameters were reached, and stabilized values were used for analysis. The chamber environment was maintained at a photosynthetic photon flux density (PPFD) of 915 µmol m−2 s−1, leaf temperature of 28 °C, relative humidity of 70%, CO2 concentration of 420 µmol mol−1, and flow rate of 700 µmol s−1 with a fan speed of 10,000 rpm. Measurements were conducted on the uppermost fully expanded leaf at each sampling date.
2.4. Measurement of Flowering Quality
On 8 September 2025 (DAT 63), nine flowering stems per treatment were harvested, with three stems randomly selected from each section. Crop management practices, including pinching, disbudding, and lateral shoot removal, were performed uniformly across all treatments according to conventional cultivation methods. The harvested stems were initially trimmed to 65 cm and then re-cut to 50 cm. The upper 20 leaves were retained, and all lower leaves were removed. The cut flowers were placed in a vase solution containing a commercial floral preservative (FloraLife® Express Universal Flower Food, FloraLife, Walterboro, SC, USA) at a concentration of 5 mL per 500 mL of tap water. The postharvest environment was maintained at a PPFD of 50 µmol m−2 s−1, a 12.5 h photoperiod, a temperature of 24.5 ± 0.8 °C, and a relative humidity of 71.2 ± 5.7%.
Flower quality traits were evaluated at the full bloom stage, defined as the stage at which the ray florets were fully expanded and the capitulum no longer increased in visible size (
Figure 3). Measured parameters included peduncle length, peduncle diameter, capitulum diameter, capitulum height, and the incidence of malformed capitula. Peduncle length was measured from the base of the involucre to the first fully expanded leaf, and peduncle diameter was measured immediately below the involucre. Capitulum diameter was determined as the average of two perpendicular measurements taken across the widest part of the capitulum. Capitulum height was measured from the base of the involucre to the top of the ray florets. Capitula showing an open center, characterized by visible exposure of the central disc region due to incomplete coverage by ray florets, were classified as malformed. The incidence of malformed capitula was calculated for each section as the number of malformed capitula divided by the total number of evaluated capitula and expressed as a percentage.
2.5. Statistical Analysis
Data were analyzed using SAS software version 9.4 (SAS Institute, Cary, NC, USA). For each response variable, the mean value of the three plants or stems sampled within each section was used as a section-level observation, resulting in three within-bed spatial subsamples per treatment. Because each cooling treatment was applied to a single cultivation bed, these section-level observations were used to characterize within-bed spatial variation rather than to provide independent bed-level biological replication. Accordingly, statistical results were interpreted as treatment-associated responses under commercial-scale demonstration conditions.
Growth and photosynthetic parameters were analyzed using a mixed model in PROC MIXED, with root-zone cooling treatment (TRT), days after treatment (DAT), and their interaction (TRT × DAT) as fixed effects. Section nested within treatment was included as a random effect to account for within-bed spatial variation. Because biomass measurements were obtained by destructive sampling at each date, repeated observations represented measurements from the same sections over time rather than repeated measurements of the same individual plants.
Continuous flowering quality traits, including peduncle length, peduncle diameter, capitulum diameter, and capitulum height, were analyzed using one-way ANOVA based on section-level observations. Mean separation was performed using Duncan’s multiple range test at α = 0.05. Figures were prepared using SigmaPlot software version 11.0 (Systat Software Inc., San Jose, CA, USA).
4. Discussion
4.1. Root-Zone Cooling Improved Vegetative Growth Without Directly Enhancing Leaf Gas Exchange
Root-zone cooling improved vegetative growth of standard chrysanthemum ‘Baekgang’ under high-temperature summer conditions, as reflected by increased plant height, stem diameter, and shoot and root biomass. This response indicates that the root-zone thermal environment played an important role in regulating plant growth, even though the aerial environment remained supra-optimal. High root-zone temperatures can impair root physiological activity by increasing maintenance respiration, accelerating carbohydrate depletion, and disrupting water, nutrient, and hormonal metabolism [
18,
19]. Conversely, maintaining the root zone within a favorable temperature range can reduce belowground heat stress and sustain root function.
The beneficial effects of root-zone cooling observed in this study are consistent with previous findings that root temperature can be a critical determinant of whole-plant performance under heat stress. In creeping bentgrass, high root-zone temperature inhibited shoot and root growth even when air temperature was maintained within a favorable range, whereas lowering root-zone temperature alleviated growth inhibition under high air temperature [
20]. Similarly, root-zone cooling has been reported to improve root physiological activity and xylem development in tomato seedlings under high air temperature, thereby enhancing water and mineral nutrient transport [
14]. These findings support the interpretation that improved root activity contributed to the enhanced shoot growth and biomass accumulation observed in the cooled treatments.
Although photosynthetic parameters changed significantly over time, root-zone cooling did not produce a consistent treatment effect on net CO
2 assimilation rate, transpiration rate, or stomatal conductance. This suggests that the growth-promoting effect of root-zone cooling was not primarily driven by direct stimulation of instantaneous leaf-level photosynthetic capacity. Similar discrepancies between growth responses and instantaneous gas-exchange measurements have been reported in root-zone temperature studies, where improved plant performance was associated not only with photosynthetic rate but also with changes in root activity, water relations, and carbon allocation [
19,
21]. While leaf gas exchange was evaluated as instantaneous midday snapshots at the single-leaf level, the root-zone cooling system operated continuously throughout the 24 h day–night cycle. This continuous cooling likely reduced excessive root maintenance respiration during both the day and night [
19]. Mitigating this belowground respiratory carbon loss would enhance the whole-plant carbon balance and long-term biomass accumulation, thereby compensating for the lack of a detectable increase in midday photosynthetic rates.
4.2. Root-Zone Cooling Reduced Open-Center Malformed Capitula Under Summer Heat Stress
The major floral abnormality observed in this study was open-center malformed capitula, characterized by visible exposure of the central disc region due to incomplete coverage by ray florets. Open-center disorder is a commercially important defect in standard chrysanthemum because it directly reduces ornamental quality and marketability. Previous studies have shown that high temperature during reproductive development can delay flowering and induce abnormal inflorescence development in chrysanthemum [
7,
8,
9]. In particular, the floret-forming stage, which occurs shortly after the initiation of short-day treatment, is highly sensitive to temperature stress because floral organs are actively differentiating from the inflorescence meristem [
8]. Similar temperature-related reproductive disorders have been reported in other chrysanthemum cultivars and floricultural crops. High day and night temperatures delayed budding and flowering and induced abnormal or withered flowers in heat-sensitive chrysanthemum cultivars such as ‘Swifty Rosso’ and ‘Swifty Yellow’ [
9]. In Lilium × formolongi, high root-zone temperature inhibited bolting and flowering induction by disrupting photoperiodic and hormone-related pathways [
22], whereas root-zone cooling promoted flowering or supported healthy growth in Cyclamen persicum and Fuchsia × hybrida under summer heat conditions [
15]. These findings support the importance of both aerial and root-zone thermal conditions in stabilizing reproductive development and flower quality in ornamental crops.
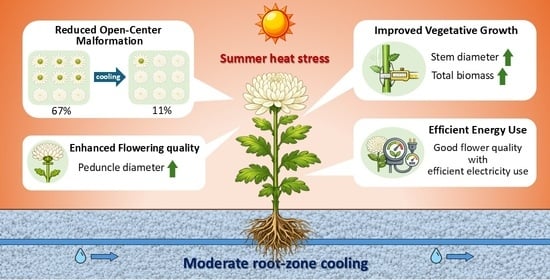
Root-zone cooling substantially reduced the incidence of open-center malformed capitula, indicating that belowground temperature management can influence reproductive development even when air temperature remains high. Although the present study did not directly measure plant water status, carbohydrate allocation, root-derived hormones, or capitulum temperature, previous studies have shown that root-zone temperature can affect sap flow, carbohydrate status, and hormone-related responses in tomato [
23] and can improve growth and photosynthetic performance under hot aerial environments in lettuce [
21,
24]. These findings suggest that root-zone temperature can influence reproductive development through integrated root-to-shoot physiological regulation, although the specific mechanisms may differ among species. In the present study, root-zone cooling increased root biomass and peduncle diameter in several treatments and reduced the incidence of open-center malformation, particularly in MT. These measured responses suggest that root-zone temperature management can influence both root and peduncle development as well as capitulum morphology under summer heat stress.
Open-center formation can also be interpreted from the perspective of capitulum developmental balance. Chrysanthemum capitulum development proceeds through coordinated differentiation and expansion of ray and disc florets, and temperature can affect flower characteristics differently depending on the developmental phase [
11]. Under high temperature, the relative timing of peripheral ray floret expansion and central disc floret development may become unbalanced, resulting in insufficient coverage of the central region by ray florets. Moderate root-zone cooling may have helped stabilize this developmental balance by reducing belowground heat stress during the temperature-sensitive period of capitulum formation.
Post and Lacey [
25] also reported that interrupted short-day treatments could improve flower form in standard chrysanthemums by altering the balance between ray and disc florets. In this study, however, the short-day and subsequent night-interruption treatments were applied uniformly across all treatments. Therefore, the differences in open-center incidence among treatments are more reasonably attributed to thermal conditions, particularly root-zone temperature, rather than to differences in photoperiodic management.
4.3. Moderate Root-Zone Cooling Provided a Better Balance Between Flower Quality and Energy Use
An important finding of this study was that the lowest root-zone temperature did not necessarily produce the best floral quality. Although LT promoted strong vegetative growth, MT showed the lowest incidence of open-center malformed capitula while consuming less electricity than LT. This indicates that the optimal root-zone cooling strategy for chrysanthemum production should not be defined simply as the lowest achievable root-zone temperature. Instead, it should be determined by balancing vegetative growth, reproductive development, flower quality, and energy consumption.
One possible explanation for the lower effectiveness of LT in suppressing open-center disorder is that intensive root-zone cooling under high aerial temperature may have altered the balance between belowground and aboveground thermal environments. Previous work in greenhouse tomato showed that root-zone temperature affects leaf–air temperature relations and plant water status, indicating that root-zone thermal conditions can interact with aerial temperature in determining whole-plant thermal and water balance [
26]. In the present study, LT maintained the mean root-zone temperature at 23.3 °C, whereas the mean daytime and nighttime air temperatures were 30.5 °C and 26.8 °C, respectively. Thus, although LT maintained the root zone near a favorable temperature range, the relatively large contrast between the cooled root zone and the hot aerial environment may have disturbed shoot–root coordination and the balance between vegetative and reproductive development. This interpretation is consistent with the higher vegetative biomass but greater open-center incidence observed in LT compared with MT. The optimum root-zone temperature is not fixed but can shift depending on air temperature and the overall thermal environment [
27], and changes in root-zone temperature can alter whole-plant functionality, including hormonal signaling and assimilate allocation [
28].
From a practical perspective, MT appeared to provide the most favorable compromise between flower quality and energy efficiency. The MT treatment maintained the root zone sufficiently cool to reduce open-center disorder, while avoiding the greater energy cost and possible excessive shoot–root thermal imbalance associated with LT. Although a full economic analysis was beyond the scope of this study, MT was more energy-efficient than LT because it achieved the lowest open-center incidence while requiring 305 kWh bed−1 less electricity than LT. These results suggest that moderate root-zone cooling can serve as an energy-saving environmental control strategy for summer chrysanthemum production. Further studies should quantify shoot–root temperature gradients, capitulum temperature, plant water status, carbohydrate allocation, hormone dynamics, and flowering-related gene expression to clarify the physiological and molecular mechanisms underlying open-center suppression by root-zone cooling.
A limitation of this study is that it was conducted using a single standard chrysanthemum cultivar, ‘Baekgang’, during one summer production season in a commercial greenhouse. In addition, because each cooling treatment was applied to a single commercial-scale cultivation bed, treatment effects and potential bed effects could not be fully separated, and the section-level observations represented within-bed spatial subsamples rather than independent bed-level biological replicates. Furthermore, because specific physiological parameters—such as root respiration, whole-plant carbon balance, shoot–root temperature gradients, and hormone dynamics—were not directly quantified, our mechanistic interpretations serve as plausible hypotheses based on previous literature. Therefore, cultivar-specific responses, seasonal variability, and greenhouse-specific environmental conditions should be considered when applying these results to broader chrysanthemum production systems. Future studies incorporating direct physiological measurements, multiple cultivars, and repeated seasonal trials under diverse greenhouse conditions are needed to validate the generality of the proposed root-zone cooling threshold and improve the practical applicability of this cooling strategy.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}