
Impact of Reduced Chemical Fertilizer and Organic Amendments on Yield, Nitrogen Use Efficiency, and Soil Microbial Dynamics in Chinese Flowering Cabbage


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Site
2.2. Experimental Design
2.3. Determination Items and Methods
2.3.1. Soil Indicators
2.3.2. Yield and Nitrogen Fertilizer Contribution Rate
2.4. Data Processing
3. Results
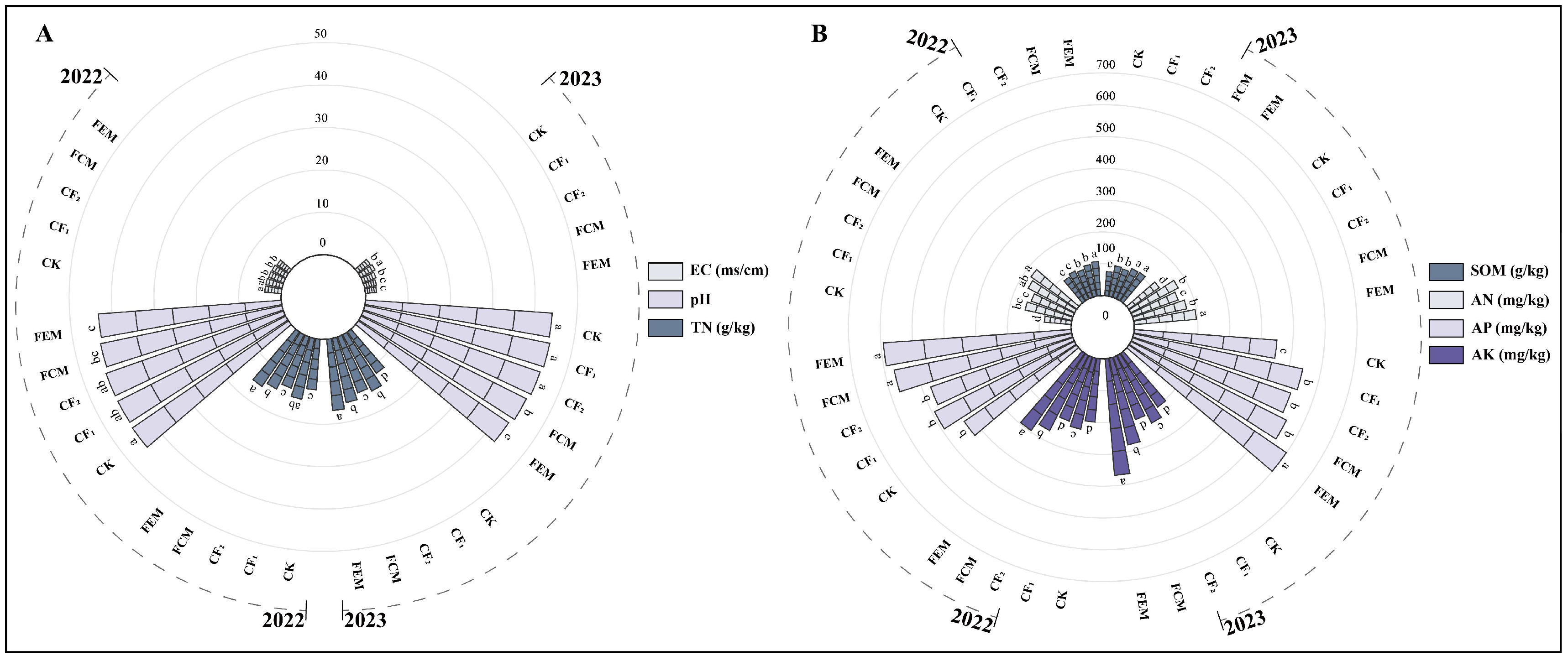
3.1. Influence of Reduced Chemical Fertilizer Combined with Organic Fertilizer on Rhizosphere Soil Nutrients
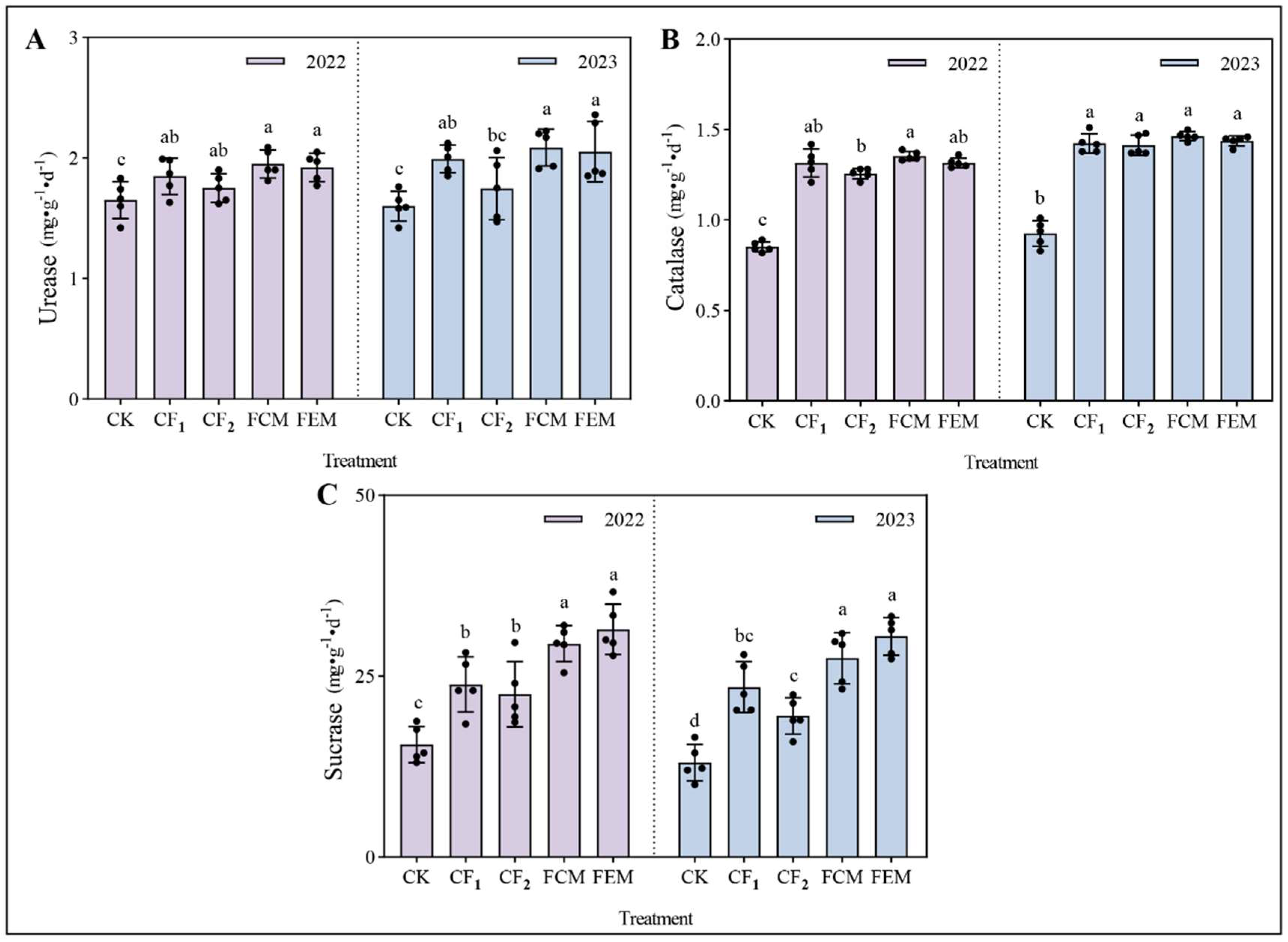
3.2. Impact of Reduced Chemical Fertilizer with Organic Amendments on Rhizosphere Soil Enzyme Activities
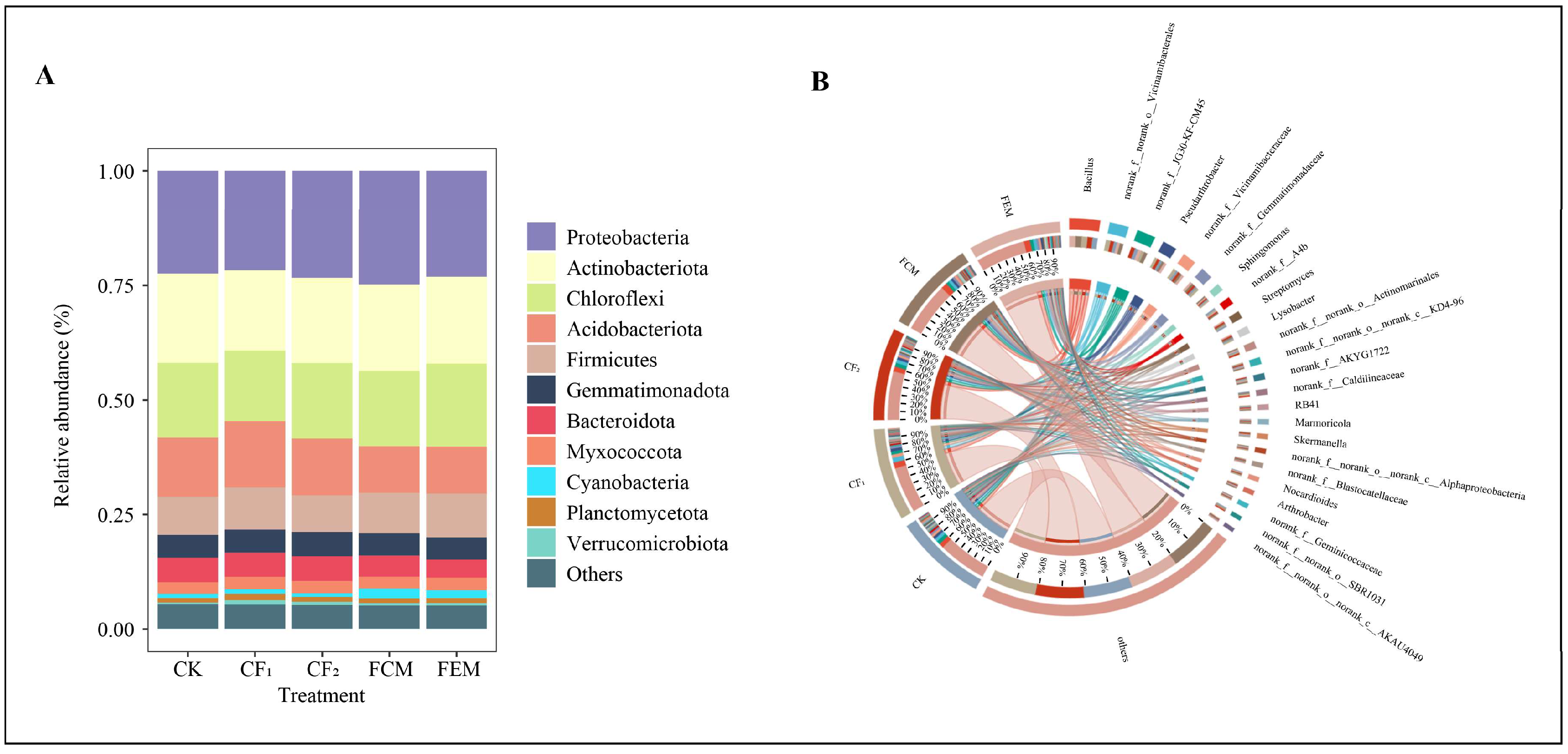
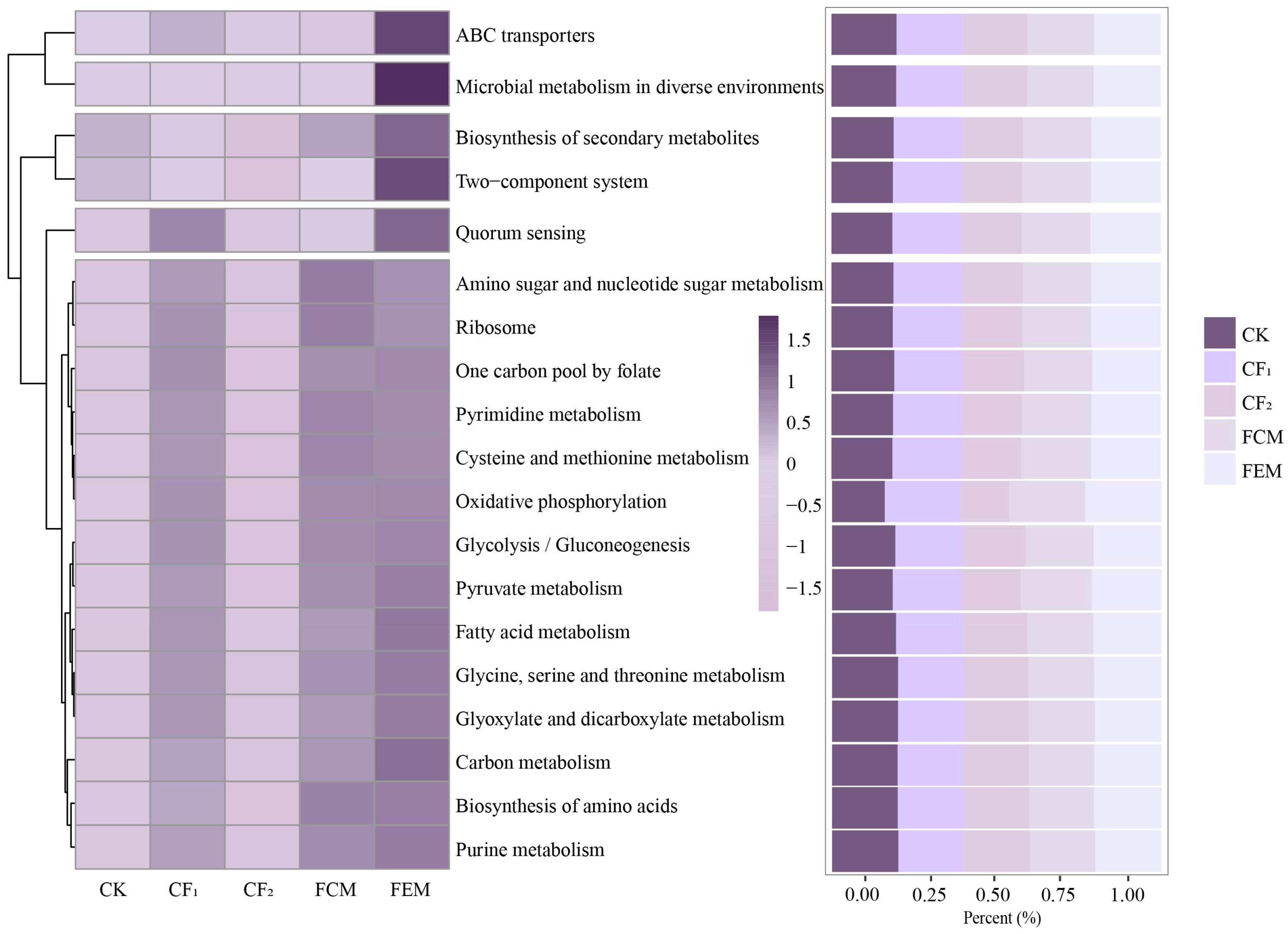
3.3. Effects of Reduced Chemical Fertilizer and Organic Fertilizer Co-Application on Bacterial Community Structure and Composition
3.4. Interaction Analysis Among Microbial Community Structure, Functional Metabolism, and Soil Environmental Factors
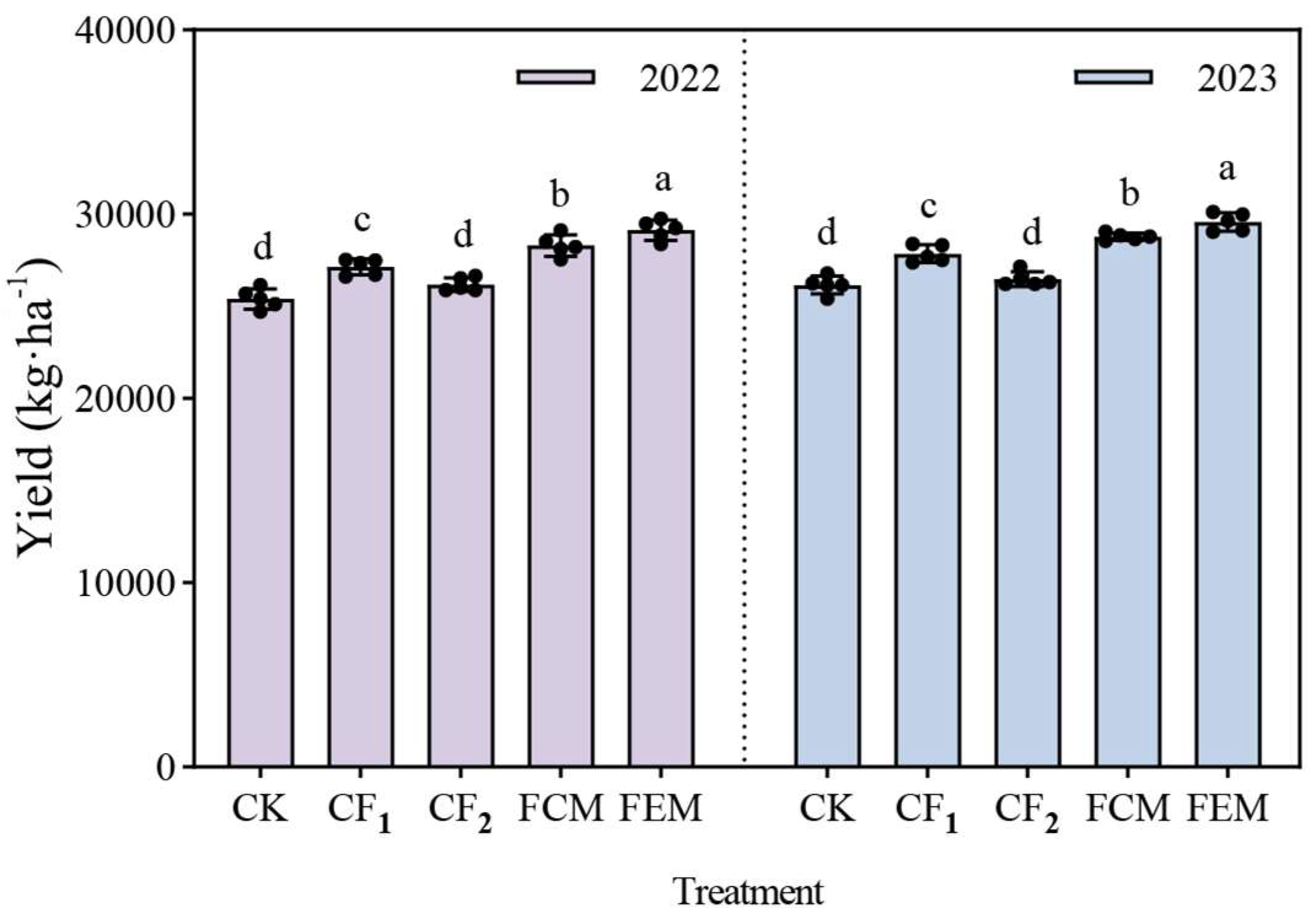
3.5. Yield Response of Flowering Chinese Cabbage to Reduced Chemical Fertilizer Integrated with Organic Fertilizer
3.6. Nitrogen Use Efficiency (NUE) of Flowering Chinese Cabbage Under Reduced Chemical Fertilizer and Organic Fertilizer Regimes
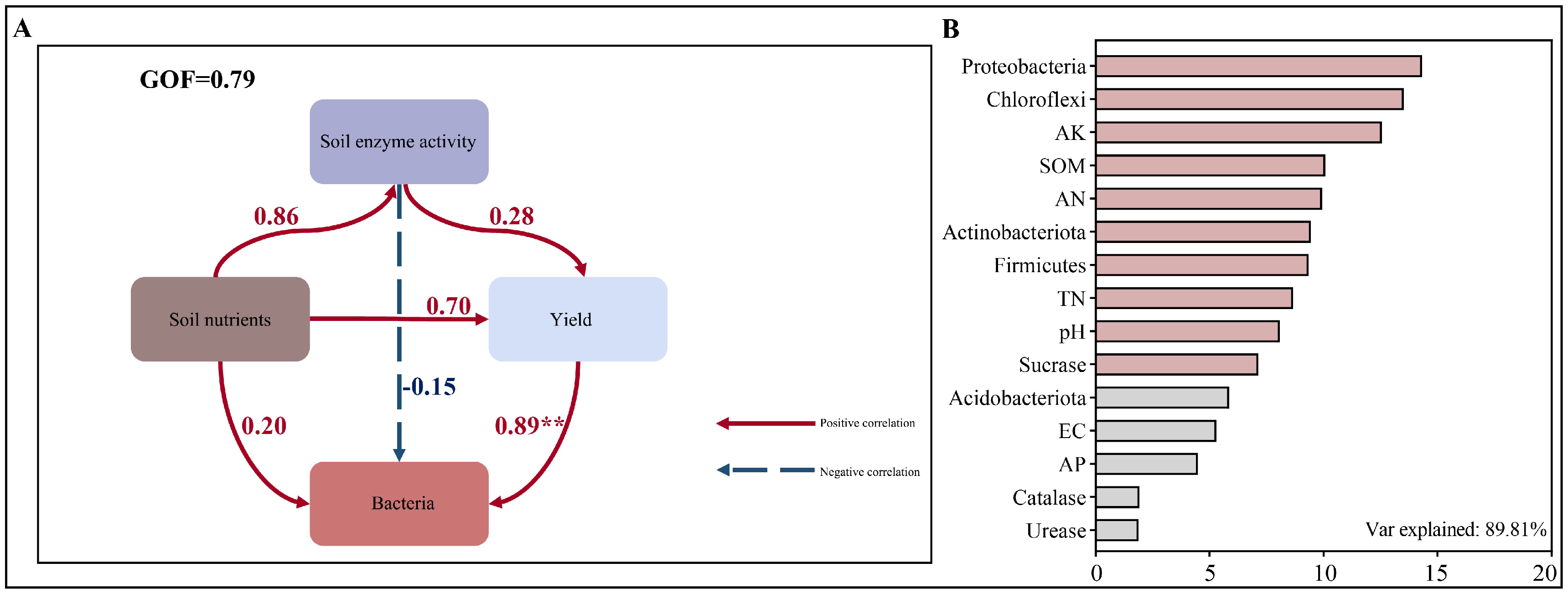
3.7. Correlation Analysis Between Crop Yield and Rhizosphere Soil Environmental Parameters
4. Discussion
4.1. Effects of Reduced Chemical Fertilization Combined with Organic Fertilizer Application on Soil Nutrients and Enzyme Activities in Chinese Flowering Cabbage
4.2. Impact of Reduced Chemical Fertilization Integrated with Organic Amendments on Soil Microbial Community in Chinese Flowering Cabbage
4.3. Influence of Chemical Fertilizer Reduction Coupled with Organic Fertilization on Yield and Nitrogen Use Efficiency of Chinese Flowering Cabbage
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dai, Y.; Zhang, F.-S.; Guan, J.-T.; Wang, S.-X.; Zhang, H.; Li, G.-L.; Sun, R.-F.; Li, F.; Zhang, S.-J. Single-Cell Transcriptomic Analysis of Flowering Regulation and Vernalization in Chinese Cabbage Shoot Apex. Hortic. Res. 2024, 11, uhae214. [Google Scholar] [CrossRef]
- Zhang, H.-Y.; Cun, Y.-H.; Wang, J.-J.; Wu, M.-Y.; Li, X.-J.; Liang, Q.-X.; Wang, C.; Zhao, L.; Deng, J.-Q. Acetylsalicylic acid and salicylic acid alleviate postharvest leaf senescence in Chinese flowering cabbage (Brassica rapa var. parachinensis). Postharvest Biol. Technol. 2022, 194, 112070. [Google Scholar] [CrossRef]
- Fu, X.; Xiao, W.; Wang, H.; Chen, Y.; Wang, R.; Wang, Y. Effects of fertigation with carboxymethyl cellulose potassium on water conservation, salt suppression, and maize growth in salt-affected soil. Agric. Water Manag. 2023, 287, 108436. [Google Scholar] [CrossRef]
- Rasool, C.; Guo, X.-M.; Wang, Z.-C.; Hassan, M.; Aleem, M.; Javed, Q.; Chen, S. Effect of Buried Straw Layer Coupled with Fertigation on Florescence and Yield Parameters of Chinese Cabbage Under Greenhouse Environment. J. Soil Sci. Plant Nutr. 2020, 20, 598–609. [Google Scholar] [CrossRef]
- Rasool, G.; Guo, X.-M.; Wang, Z.-C.; Sheng, C.-D.; Hamoud, Y.-A.; Javed, Q. Response of Fertigation Under Buried Straw Layer on Growth, Yield, and Water-fertilizer Productivity of Chinese Cabbage Under Greenhouse Conditions. Commun. Soil Sci. Plant Anal. 2019, 50, 1030–1043. [Google Scholar] [CrossRef]
- Gong, Z.-Y.; Gao, X.; Huang, Y.; Li, R.; Wang, D.-S.; Shen, Q.-R. Research on continuous application of bio-organic fertilizer for improving greenhouse cucumber yield and quality. J. Nanjing Agric. Univ. 2016, 39, 777–783. [Google Scholar] [CrossRef]
- Gao, P.; Zhang, T.; Lei, X.-Y.; Chen, X.-W.; Lu, Y.-H.; Fan, L.-S.-P.; Huang, J.; Gao, J.-Y.; Zhang, Z.-H.; Zhang, H.-M. Improvement of soil fertility and rice yield after long-term application of cow manure combined with inorganic fertilizers. J. Integr. Agric. 2023, 22, 2221–2232. [Google Scholar] [CrossRef]
- Lv, Z.-Z.; Wu, X.-D.; Hou, H.-X.; Ji, J.-H.; Liu, X.-M.; Liu, Y.-R. Effect of different application ratios of chemical and organic fertilizers on soil quality in double cropping paddy fields. J. Plant Nutr. Fertil. 2017, 23, 904–913. [Google Scholar] [CrossRef]
- Gu, S.-Y.; Geng, Z.-M.; Wang, R.; Nie, Y.-L.; Hu, Y.; Cai, H.-S.; Wang, J.-J. Effect of different ratio of bio- organic fertilizer on saline- alkali soil improvement. J. Northeast Agric. Univ. (Engl. Ed.). 2014, 45, 26–30. [Google Scholar] [CrossRef]
- Zhao, N.; Dai, C.-Y.; Han, G.-J.; Fang, C.-Y.; Chen, N.-L.; Zhang, Z.; Huang, H.-X. Effect of organic management on soil fertility and wheat production in Minqin oasis. ARAA 2012, 30, 76–81. [Google Scholar] [CrossRef]
- Seufert, V.; Ramankutty, N.; Foley, J.-A. Comparing the yields of organic and conventional agriculture. Nature 2012, 485, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Kumar, U.; Kumar, N.-A.; Shahid, M.; Gupta, V.-V.-S.-R.; Panneerselvam, P.; Mohanty, S.; Kaviraj, M.; Kumar, A.; Chatterjee, D.; Lal, B.; et al. Continuous application of inorganic and organic fertilizers over 47 years in paddy soil alters the bacterial community structure and its influence on rice production. Agric. Ecosyst. Environ. 2018, 262, 65–75. [Google Scholar] [CrossRef]
- Huang, L.-F.; Song, L.-X.; Xia, X.-J.; Mao, W.-H.; Shi, K.; Zhou, Y.-H.; Yu, J.-N. Plant-Soil Feedbacks and Soil Sickness: From Mechanisms to Application in Agriculture. Chem. Ecol. 2013, 39, 232–242. [Google Scholar] [CrossRef]
- Cai, F.; Pang, G.; Li, R.-X.; Li, R.; Gu, X.-P.; Chen, W. Bioorganic fertilizer maintains a more stable soil microbiome than chemical fertilizer for monocropping. Biol. Fertil. Soils 2017, 53, 861–872. [Google Scholar] [CrossRef]
- Schoebitz, M.; Mengual, C.; Roldán, A. Combined effects of clay immobilized Azospirillum brasilense and Pantoea dispersa and organic olive residue on plant performance and soil properties in the revegetation of a semiarid area. Sci. Total Environ. 2014, 466–467, 67–73. [Google Scholar] [CrossRef]
- Brennan, E.-B.; Acosta, M.-V. Cover cropping frequency is the main driver of soil microbial changes during six years of organic vegetable production. Soil Biol. Biochem. 2017, 109, 188–204. [Google Scholar] [CrossRef]
- Finkel, O.-M.; Castrillo, C.; Paredes, S.-H.; González, I.-S.; Dangl, J.-L. Understanding and exploiting plant beneficial microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef]
- Han, J.-Q.; Dong, Y.-Y.; Zhang, M. Chemical fertilizer reduction with organic fertilizer effectively improve soil fertility and microbial community from newly cultivated land in the Loess Plateau of China. Appl Soil Ecol. 2021, 165, 103966. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agricultural Chemical Analysis, 3rd ed.; China Agricultural Press: Beijing, China, 2000; pp. 25–144. Available online: https://libopac.bift.edu.cn (accessed on 28 June 2025).
- Yang, J.-H.; Wang, C.-L.; Dai, H.-L. Soil Agrochemica Analysis and Environmental Monitoring; China Land Publishing House: Beijing, China, 2008. Available online: https://www.shukui.net/book/1828543.html (accessed on 28 June 2025).
- Guan, S.-Y.; Zhang, D.; Zhang, Z. Soil Enzyme and Its Research Methods; Chinese Agricultural Press: Beijing, China, 1986; pp. 294–297. Available online: http://opac.stlib.cn (accessed on 28 June 2025).
- Zhang, S.-N.; Wang, Y.; Sun, L.-T.; Qiu, C.; Ding, Y.-Q.; Gu, H.-L. Organic mulching positively regulates the soil microbial communities and ecosystem functions in tea plantation. BMC Microbiol. 2020, 20–103. [Google Scholar] [CrossRef]
- Guo, H.-J.; Gao, J.; Zhang, Q.; Li, J.-H.; Jin, D.-S.; Xu, M.-G. Differences and reasons for the effects of organic fertilizer on the pH of acidic and alkaline soils in China. Chin. J. Appl. Environ. Biol. 2024, 30, 496–503. [Google Scholar] [CrossRef]
- Wang, X.-X.; Zhao, F.-J.; Zhang, G.-X.; Zhang, Y.-Y.; Yang, L.-J. Vermicompost Improves Tomato Yield and Quality and the Biochemical Properties of Soils with Different Tomato Planting History in a Greenhouse Study. Front. Plant Sci. 2017, 8, 1978. [Google Scholar] [CrossRef]
- Chintala, R.; Mollinedo, J.; Schumacher, T.-E. Effect of biochar on chemical properties of acidic soil. Arch. Agron. Soil Sci. 2014, 60, 393–404. [Google Scholar] [CrossRef]
- Zhang, Y.-Y.; Bai, G.-W.; Yu, J.-H.; Wang, F.; Liu, L.-J.; Zhu, L.; Wu, Y.-D.-L. Screening, identification, and enzyme production condition optimization of a cellulose-degrading bacterial strain. Microbiol. China 2025, 1–20. [Google Scholar] [CrossRef]
- Pinggera, J.; Geisseler, D.; Piepho, H.-P.; Joergensen, R.-G.; Ludwig, B. Effect of substrate quality on the N uptake routes of soil microorganisms in different soil depths. Pedobiologia 2015, 58, 211–218. [Google Scholar] [CrossRef]
- Li, Q.-J.; Zhang, D.-Q.; Song, Z.-X.; Ren, L.-R.; Jin, X.; Fang, W.-S.; Yan, D.-D.; Li, Y.; Wang, Q.-W.; Cao, A. Organic fertilizer activates soil beneficial microorganisms to promote strawberry growth and soil health after fumigation. Environ. Pollut. 2022, 295, 118653. [Google Scholar] [CrossRef]
- Liu, Q.; Mu, X.; Zhao, G.-J.; Cao, Q.-C.; Sun, W.-L. Effects of fertilization on soil water use efficiency and crop yield on the loess plateau, China. Appl. Ecol. Env. Res. 2020, 18, 6555–6568. [Google Scholar] [CrossRef]
- Ding, J.-L.; Jiang, X.; Ma, M.-C.; Zhou, B.-K.; Guan, D.-K.; Zhao, B.; Zhou, J.; Cao, F.-M.; Li, L.; Li, J. Effect of 35 years inorganic fertilizer and manure amendment on structure of bacterial and archaeal communities in black soil of northeast China. Appl. Soil Ecol. 2016, 105, 187–195. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.-C.; Wang, H.-Y.; Gai, X.-P. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Torsvik, V.; Øvreås, L. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Liu, Z.-D.; Jiao, Y.-Q.; Yin, J.; Dong, L.; Wang, B.-B.; Zhang, K.-L.; Zheng, X.-X.; Hong, Y.-B.; Zhang, H.-X.; Xie, C.; et al. Productivity and quality of banana in response to chemical fertilizer reduction with bio-organic fertilizer: Insight into soil properties and microbial ecology. Agric. Ecosyst. Environ. 2021, 322, 107659. [Google Scholar] [CrossRef]
- Liu, J.; Shu, A.-P.; Song, W.-F.; Shi, W.-C.; Li, M.-C.; Zhang, W.-X.; Li, Z.-Z.; Liu, G.-G.; Yuan, F.-M.; Zhang, S.-X.; et al. Long-term organic fertilizer substitution increases rice yield by improving soil properties and regulating soil bacteria. Geoderma 2021, 404, 115287. [Google Scholar] [CrossRef]
- Aloo, B.-N.; Makumba, B.-A.; Mbega, E.-R. The potential of Bacilli rhizobacteria for sustainable crop production and environmental sustainability. Microbiol. Res. 2019, 219, 26–39. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.-L.; Ramirez, K.-S.; Zaneveld, J.; Bradford, M.-A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.-L.; Zhang, L.-S. Alleviation of vermicompost to obstacle in sterilized continuous cropping soil in strawberry production. Plant Nutr. Fert. Sci. 2016, 22, 759–767. [Google Scholar] [CrossRef]
- Chen, X.-J.; Cong, W.-W.; Chen, Y.-L.; Lu, N.; Cheng, J.; Cao, X.-L.; Dai, H.-W.; Yao, F.-X.; Sheng, H.-J.; Qian, X.-Q. Effect of earthworm manure on improvement of red soil and its effect on growth of non-pelleting Chinese cabbage. J. Agric. Sci. 2016, 44, 248–250. [Google Scholar] [CrossRef]
- Yu, Y.-C.; Wang, C.; Sun, M.-S.; Liu, M.-L. Meliorative effect of earthworm-mycorrhiza interaction on coastal saline soil. J. China Agric. Univ. 2019, 24, 123–129. [Google Scholar]
- Liu, Z.-L.; Gao, J.-J.; Chen, Z.; Yan, W.-Q.; Gu, R.-Y. Effects of nitrogen fertilizer reduction combined with organic fertilizeron yield, quality and nitrogen utilization efficiency of Chinese Cabbage. J. Irrig. Drain. Eng. 2022, 40, 1138–1144. [Google Scholar] [CrossRef]
- Wu, X.-K.; Jiang, Z.-C.; Lu, Z.-X.; Lu, G.; Xu, Y.-H.; Shi, W.-M.; Min, J. Effects of the partial replacement of chemical fertilizer with manure on the yield and nitrogen emissions in leafy vegetable production. Chin. J. Eco-Agric. 2020, 28, 349–356. [Google Scholar] [CrossRef]
- Liu, W.; Wang, S.-T.; Lin, P.; Sun, H.-W.; Hou, J.; Zuo, Q.-Q.; Huo, R. Response of CaCl2-extractable heavy metals, polychlorinated biphenyls, and microbial communities to biochar amendment in naturally contaminated soils. J. Soil Sediment. 2016, 16, 476–485. [Google Scholar] [CrossRef]
- Xun, W.-B.; Xiong, W.; Huang, T.; Wei, R.; Li, D.-C.; Shen, Q.-R.; Lu, Q.; Zhang, R.-F. Swine manure and quicklime have different impacts on chemical properties and composition of bacterial communities of an acidic soil. Appl. Soil Ecol. 2016, 100, 38–44. [Google Scholar] [CrossRef]
- Wu, Y.-R.; Yang, W.-Q.; Li, Q.-Y.; Qiao, Q.-L.; Zhao, S.; Zhang, Y.-L.; Yu, Y.-H.; Zhang, S.-X.; Li, X.-L.; Kou, J.-C. Microbial Community Response to Alpine Meadow Degradation and Its Impact on Soil Nutrient Cycling. Agron. J. 2024, 14, 195. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, C.-J.; Guo, J.-H.; Guan, H.-L.; Li, L.; Tang, G.-M.; Huang, J.-X. The Effects of Cadmium Stress on Seed Germination and Seedling Growth of Bidens Pilosa (L.) and Pennisetum Alopecuroides (L.). J. Yunnan Univ. Nat. Sci. Ed. 2022, 42, 50–57 + 63. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Fertilizer Design | Urea (kg·hm−2) | Calcium Superphosphate (kg·hm−2) | Potassium Sulfate (kg·hm−2) | Organic Fertilizer (kg·hm−2) |
|---|---|---|---|---|---|
| CK | no fertilizer | 0 | 0 | 0 | 0 |
| CF1 | chemical fertilizer only | 260 | 120 | 110 | 0 |
| CF2 | 30% less chemical fertilizer | 182 | 84 | 77 | 0 |
| FCM | 30% less chemical fertilizer + rotted chicken manure | 182 | 84 | 77 | 11,000 |
| FEM | 30% less chemical fertilizer + vermicompost | 182 | 84 | 77 | 13,500 |
| Year | Treatment | Nitrogen Agronomic Efficiency (kg·kg−1) | Nitrogen Partial Factor Productivity (umol·m−2·s−1) | Nitrogen Fertilizer Reductions (%) |
|---|---|---|---|---|
| 2022 | CF1 | 14.47 ± 3.47b | 226.93 ± 1.61d | 6.34 ± 1.50b |
| CF2 | 9.29 ± 2.38b | 312.80 ± 1.96a | 2.97 ± 0.76b | |
| FCM | 24.20 ± 1.71a | 236.65 ± 2.16c | 10.22 ± 0.69a | |
| FEM | 31.14 ± 3.19a | 243.59 ± 2.00b | 12.75 ± 1.23a | |
| 2023 | CF1 | 14.27 ± 1.64b | 232.95 ± 1.77d | 6.12 ± 0.68b |
| CF2 | 3.89 ± 3.10c | 316.29 ± 2.08a | 1.22 ± 0.97c | |
| FCM | 21.96 ± 1.21ab | 240.64 ± 0.73c | 9.13 ± 0.52a | |
| FEM | 28.66 ± 2.73a | 247.33 ± 1.83b | 11.56 ± 1.03a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Li, J.; Zhao, X.; Liu, Z.; Xu, H.; Cao, K.; Ye, L. Impact of Reduced Chemical Fertilizer and Organic Amendments on Yield, Nitrogen Use Efficiency, and Soil Microbial Dynamics in Chinese Flowering Cabbage. Horticulturae 2025, 11, 859. https://doi.org/10.3390/horticulturae11070859
Xu J, Li J, Zhao X, Liu Z, Xu H, Cao K, Ye L. Impact of Reduced Chemical Fertilizer and Organic Amendments on Yield, Nitrogen Use Efficiency, and Soil Microbial Dynamics in Chinese Flowering Cabbage. Horticulturae. 2025; 11(7):859. https://doi.org/10.3390/horticulturae11070859
Chicago/Turabian StyleXu, Jiaxin, Jianshe Li, Xia Zhao, Zhen Liu, Hao Xu, Kai Cao, and Lin Ye. 2025. "Impact of Reduced Chemical Fertilizer and Organic Amendments on Yield, Nitrogen Use Efficiency, and Soil Microbial Dynamics in Chinese Flowering Cabbage" Horticulturae 11, no. 7: 859. https://doi.org/10.3390/horticulturae11070859
APA StyleXu, J., Li, J., Zhao, X., Liu, Z., Xu, H., Cao, K., & Ye, L. (2025). Impact of Reduced Chemical Fertilizer and Organic Amendments on Yield, Nitrogen Use Efficiency, and Soil Microbial Dynamics in Chinese Flowering Cabbage. Horticulturae, 11(7), 859. https://doi.org/10.3390/horticulturae11070859