


Calcium Route in the Plant and Blossom-End Rot Incidence



Abstract
1. Introduction
2. Function of Calcium in Plants
2.1. Structural Role of Calcium
2.2. Enzymatic Role of Calcium
2.3. Calcium and Signal Transduction
2.3.1. Calcium Proteins
2.3.2. Calcium Channels
2.3.3. Calcium Efflux Systems
3. Plant Calcium Uptake by the Root System
4. Calcium Uptake Through Foliar Application
5. Calcium Uptake Through the Fruit
6. Calcium Translocations
6.1. Calcium Translocations Within the Plant
6.2. Leaf or Fruit?
6.3. Calcium Translocation Within the Fruit
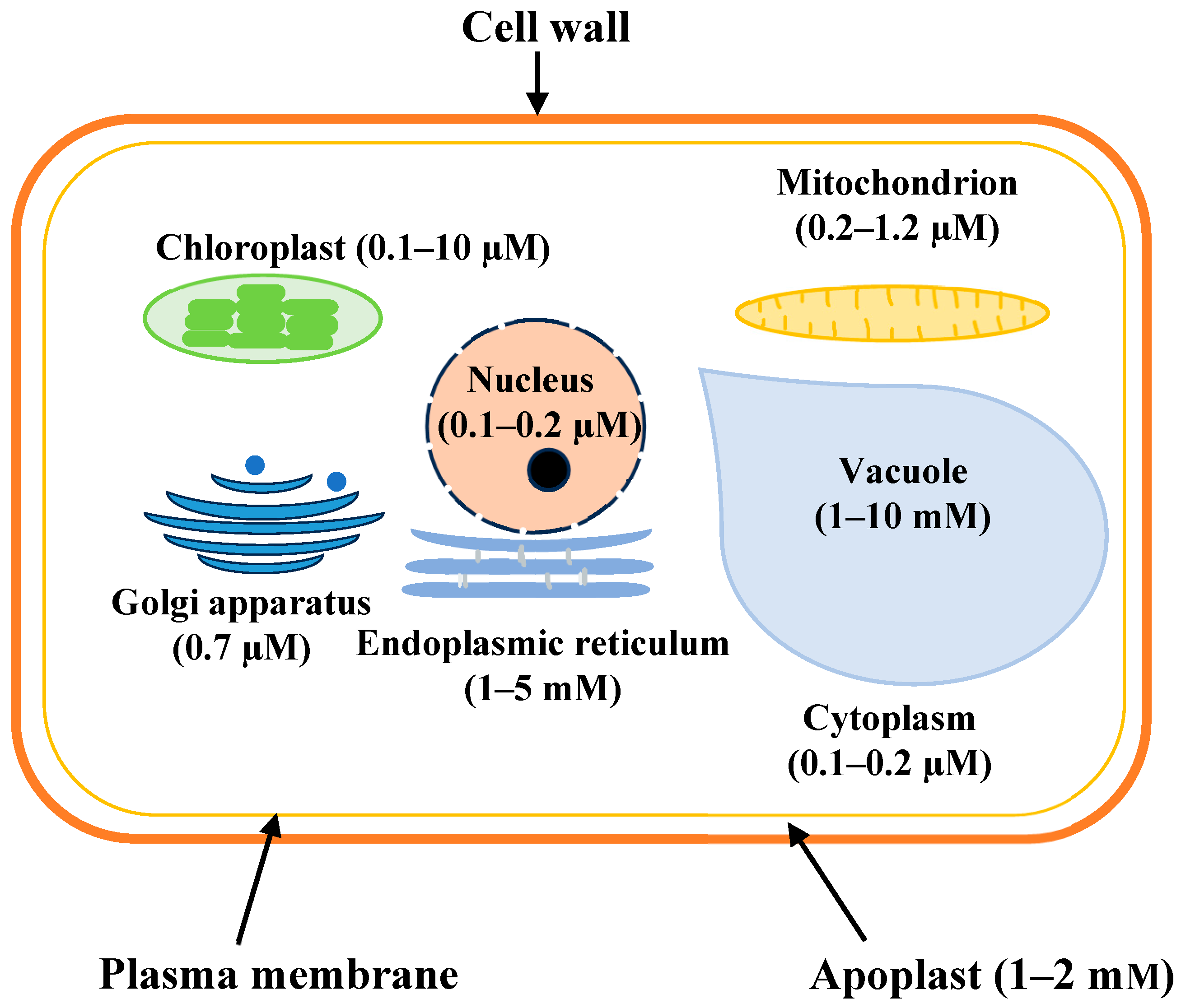
7. Calcium Compartmentalization Within the Cell
8. Calcium Deficiency Disorders

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Deficiency Symptoms | Crops | Description | Reference |
|---|---|---|---|
| Blossom-end rot | Bell pepper, tomato, watermelon, eggplant, squash | Blossom-end rot in fruit and vegetables develops dry, brown/black, sunken spots, leading to rotting that may cover most of the fruit. | [21,23,25,177,187,188] |
| Blackheart | Celery | Young leaf tissue collapses and turns black, usually at the center (heart) of celery. | [132,186] |
| Bitter pit | Apple | Development of brown/black depressed spots on the fruits. | [21,138,164] |
| Empty pod | Peanut | Poor or no development of the seed kernel, resulting in the empty pod/shell of the peanut. | [21] |
| Tip burn | Cabbage, Chinese cabbage, other cabbages | The tip of rapidly growing young leaves becomes necrotic. | [24,26,183] |
| Brussels sprouts, lettuce | Necrosis of the tip of rapidly growing young leaves. | [24,184,185] | |
| Chervil | The tip of rapidly growing young leaves becomes necrotic. | [24,182] | |
| Chicory, escarole, onion, fennel, potatoes | Necrosis of the tip of rapidly growing young leaves. | [24] | |
| Brown heart | Leafy vegetables | Necrosis of the tip of young leaves that covers the entire leaf successively. | [21] |
| Fruit cracking | Tomato, cherry, apple | Splitting of skin or cuticle. | [21] |
8.1. Genesis of Blossom-End Rot Development
8.2. Incidence of BER Based on Variety, Season, and Truss
9. Control of BER
10. Climate Change and Calcium Availability
11. Recent Advances and Future Directions
11.1. Nanotechnology-Based Calcium Delivery Systems
11.2. CRISPR-Mediated Gene Manipulation of Calcium Transporters
11.3. Imaging and Biosensors for Calcium Dynamics
11.4. Future Directions
12. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Keiser, J.R.; Mullen, R.E. Calcium and relative humidity effects on soybean seed nutrition and seed quality. Crop Sci. 1993, 33, 1345–1349. [Google Scholar] [CrossRef]
- McLaughlin, S.B.; Wimmer, R. Tansley Review No. 104 Calcium physiology and terrestrial ecosystem processes. New Phytol. 1999, 142, 373–417. [Google Scholar] [CrossRef]
- Zartdinova, R.; Nikitin, A. Calcium in the life cycle of legume root nodules. Indian J. Microbiol. 2023, 63, 410–420. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology Online Fifth Edition [WWW Document]. 2011. Available online: http://5e.plantphys.net (accessed on 31 May 2025).
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Chapter 6—Functions of Macronutrients A2-Marschner, Petra. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Saito, S.; Uozumi, N. Calcium-regulated phosphorylation systems controlling uptake and balance of plant nutrients. Front. Plant Sci. 2020, 11, 44. [Google Scholar] [CrossRef]
- Kim, M.C.; Chung, W.S.; Yun, D.J.; Cho, M.J. Calcium and calmodulin-mediated regulation of gene expression in plants. Mol. Plant 2009, 2, 13–21. [Google Scholar] [CrossRef]
- Feng, D.; Wang, X.; Gao, J.; Zhang, C.; Liu, H.; Liu, P.; Sun, X. Exogenous calcium: Its mechanisms and research advances involved in plant stress tolerance. Front. Plant Sci. 2023, 14, 1143963. [Google Scholar] [CrossRef]
- Hu, W.; Liu, J.; Liu, T.; Zhu, C.; Wu, F.; Jiang, C.; Wu, Q.; Chen, L.; Lu, H.; Shen, G.; et al. Exogenous calcium regulates the growth and development of Pinus massoniana detecting by physiological, proteomic, and calcium-related gene expression analysis. Plant Physiol. Biochem. 2023, 196, 1122–1136. [Google Scholar] [CrossRef]
- Hu, W.; Tian, S.B.; Di, Q.; Duan, S.H.; Dai, K. Effects of exogenous calcium on mesophyll cell ultrastructure, gas exchange, and photosystem II in tobacco (Nicotiana tabacum Linn.) under drought stress. Photosynthetica 2018, 56, 1204–1211. [Google Scholar] [CrossRef]
- Sun, X.; Pan, B.; Wang, Y.; Xu, W.; Zhang, S. Exogenous calcium improved resistance to Botryosphaeria dothidea by increasing autophagy activity and salicylic acid level in pear. Mol. Plant-Microbe Interact. 2020, 33, 1150–1160. [Google Scholar] [CrossRef]
- Marschner, H. 8-Functions of Mineral Nutrients: Macronutrients. In Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 1995. [Google Scholar]
- Kulik, L.V.; Epel, B.; Lubitz, W.; Messinger, J. Electronic structure of the Mn4O x Ca cluster in the S0 and S2 States of the oxygen-evolving complex of photosystem II based on pulse 55Mn-ENDOR and EPR spectroscopy. J. Am. Chem. Soc. 2007, 129, 13421–13435. [Google Scholar] [CrossRef]
- McAinsh, M.R.; Hetherington, A.M. Encoding specificity in Ca2+ signalling systems. Trends Plant Sci. 1998, 3, 32–36. [Google Scholar] [CrossRef]
- Simon, E.W. The symptoms of calcium deficiency in plants. New Phytol. 1978, 80, 1–15. [Google Scholar] [CrossRef]
- Alonso, M.T.; Torres-Vidal, P.; Calvo, B.; Rodriguez, C.; Delrio-Lorenzo, A.; Rojo-Ruiz, J.; Garcia-Sancho, J.; Patel, S. Use of aequorin-based indicators for monitoring Ca2+ in acidic organelles. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2023, 1870, 119481. [Google Scholar] [CrossRef]
- Vaughan, M.K. Arginine vasotocin and vertebrate reproduction. In The Pineal Gland; CRC Press: Boca Raton, FL, USA, 2020; pp. 125–163. [Google Scholar]
- Lecourieux, D.; Mazars, C.; Pauly, N.; Ranjeva, R.; Pugin, A. Analysis and effects of cytosolic free calcium increases in response to elicitors in Nicotiana plumbaginifolia cells. Plant Cell 2002, 14, 2627–2641. [Google Scholar] [CrossRef]
- Carafoli, E. Calcium–a universal carrier of biological signals: Delivered on 3 July 2003 at the Special FEBS Meeting in Brussels. FEBS J. 2005, 272, 1073–1089. [Google Scholar] [CrossRef]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley & Sons: Hoboken, NJ, USA, 1995. [Google Scholar]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Morard, P.; Lacoste, L.; Silvestre, J. Effects of calcium deficiency on nutrient concentration of xylem sap of excised tomato plants. J. Plant Nutr. 2000, 23, 1051–1062. [Google Scholar] [CrossRef]
- Díaz-P’erez, J.C.; Hook, J.E. Plastic-mulched bell pepper (Capsicum annuum L.) plant growth and fruit yield and quality as influenced by irrigation rate and calcium fertilization. HortScience 2017, 52, 774–781. [Google Scholar] [CrossRef]
- Olle, M.; Bender, I. Causes and control of calcium deficiency disorders in vegetables: A review. J. Hortic. Sci. Biotechnol. 2009, 84, 577–584. [Google Scholar] [CrossRef]
- Taylor, M.D.; Locascio, S.J. Blossom-end rot: A calcium deficiency. J. Plant Nutr. 2004, 27, 123–139. [Google Scholar] [CrossRef]
- Aloni, B.; Pashkar, T.; Libel, R. The possible involvement of gibberellins and calcium in tipburn of Chinese cabbage: Study of intact plants and detached leaves. Plant Growth Regul. 1986, 4, 3–11. [Google Scholar] [CrossRef]
- Geraldson, C.M. The use of calcium for control of blossom-end rot of Tomatoes. Proc. Fla. State Hortic. Soc. 1956, 68, 197–202. [Google Scholar]
- Adams, P.; El-Gizawy, A.M. Effect of calcium stress on the calcium status of tomatoes grown in NFT. Acta Hortic. 1988, 222, 15–22. [Google Scholar] [CrossRef]
- Banuelos, G.S.; Offermann, G.P.; Seim, E.C. High relative humidity promotes blossom-end rot on growing tomato fruit. HortScience 1985, 20, 894–895. [Google Scholar] [CrossRef]
- Barke, R.E. Absorption and translocation of calcium foliar sprays in relation to the incidence of blossom-end rot in tomatoes. Qld. J. Agric. Anim. Sci. 1968, 25, 179–197. [Google Scholar]
- Cho, I.H.; Lee, E.H.; Kim, T.Y.; Woo, Y.H.; Kwon, Y.S. Effects of high humidity on occurrence of tomato blossom-end rot. J. Korean Soc. Hortic. Sci. 1998, 39, 47–249. [Google Scholar]
- El-Gizawy, A.M.; Adams, P. Effect of temporary calcium stress on the calcium status of tomato fruit and leaves. Symp. Nutr. Grow. Tech. Plant Substrates 1985, 178, 37–44. [Google Scholar] [CrossRef]
- Gutteridge, C.G.; Bradfield, E.G. Root pressure stops blossom-end rot. Grower 1983, 100, 25–26. [Google Scholar]
- Saure, M.C. Blossom-end rot of tomato (Lycopersicon esculentum Mill.)—A calcium-or a stress-related disorder? Sci. Hortic. 2001, 90, 193–208. [Google Scholar] [CrossRef]
- Spurr, A.R. Anatomical aspects of blossom-end rot in the tomato with special reference to calcium nutrition. Hilgardia 1959, 28, 269–295. [Google Scholar] [CrossRef]
- Wada, T.; Ikeda, H.; Ikeda, M.; Furukawa, H. Effects of foliar application of calcium solutions on the incidence of blossom-end rot of tomato fruit. J. Jpn. Soc. Hortic. Sci. 1996, 65, 553–558. [Google Scholar] [CrossRef]
- Westerhout, J. Relation of fruit development to the incidence of blossom-end rot of tomatoes. Neth. J. Agric. Sci. 1962, 10, 223–234. [Google Scholar] [CrossRef]
- Wui, M.; Takano, T. Effect of Temperature and Concentration of Nutrient Solution during the Stage of the Fruit Development on the Incidence of Blossom-End Rot in Fruits of Tomato, Lycopersicon esculentum L. Environ. Control Biol. 1995, 33, 7–14. [Google Scholar] [CrossRef]
- De Freitas, S.T.; McElrone, A.J.; Shackel, K.A.; Mitcham, E.J. Calcium partitioning and allocation and blossom-end rot development in tomato plants in response to whole-plant and fruit-specific abscisic acid treatments. J. Exp. Bot. 2014, 65, 235–247. [Google Scholar] [CrossRef]
- Abdelkader, M.; Elkhawaga, F.A.; Suliman, A.A.; Puchkov, M.; Kuranova, K.N.; Mahmoud, M.H.; Abdelkader, M.F. Understanding the Regular Biological Mechanism of Susceptibility of Tomato Plants to Low Incidences of Blossom-End Rot. Horticulturae 2024, 10, 648. [Google Scholar] [CrossRef]
- Kabir, M.Y.; Díaz-Pérez, J.C.; Doyle, J.W.; Berenguer, E.I.; van der Knaap, E.; Nambeesan, S.U. The Effect of Calcium Application and Irrigation on Development of Blossom-end Rot in Tomato. In Proceedings of the 2017 ASHS Annual Conference, Waikoloa, HI, USA, 19–22 September 2017. [Google Scholar]
- De Freitas, S.T.; Padda, M.; Wu, Q.; Park, S.; Mitcham, E.J. Dynamic alternations in cellular and molecular components during blossom-end rot development in tomatoes expressing sCAX1, a constitutively active Ca2+/H+ antiporter from Arabidopsis. Plant Physiol. 2011, 156, 844–855. [Google Scholar] [CrossRef]
- Gao, H.; Wu, X.; Zorrilla, C.; Vega, S.E.; Palta, J.P. Fractionating of calcium in tuber and leaf tissues explains the calcium deficiency symptoms in potato plant overexpressing CAX1. Front. Plant Sci. 2020, 10, 1793. [Google Scholar] [CrossRef]
- Park, S.; Kim, C.K.; Pike, L.M.; Smith, R.H.; Hirschi, K.D. Increased calcium in carrots by expression of an Arabidopsis H+/Ca 2+ transporter. Mol. Breed. 2004, 14, 275–282. [Google Scholar] [CrossRef]
- Wu, Q.; Shigaki, T.; Han, J.S.; Kim, C.K.; Hirschi, K.D.; Park, S. Ectopic expression of a maize calreticulin mitigates calcium deficiency-like disorders in sCAX1-expressing tobacco and tomato. Plant Mol. Biol. 2012, 80, 609–619. [Google Scholar] [CrossRef]
- Zorrilla, C.; Schabow, J.E.; Chernov, V.; Palta, J.P. CAX1 vacuolar antiporter overexpression in potato results in calcium deficiency in leaves and tubers by sequestering calcium as calcium oxalate. Crop Sci. 2019, 59, 176–189. [Google Scholar] [CrossRef]
- Wyatt, S.E.; Tsou, P.L.; Robertson, D. Expression of the high capacity calcium-binding domain of calreticulin increases bioavailable calcium stores in plants. Transgenic Res. 2002, 11, 1–10. [Google Scholar] [CrossRef]
- Kuronuma, T.; Watanabe, H. Identification of the Causative Genes of Calcium Deficiency Disorders in Horticulture Crops: A Systematic Review. Agriculture 2021, 11, 906. [Google Scholar] [CrossRef]
- Thor, K. Calcium—Nutrient and messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef]
- Hirschi, K.D. The calcium conundrum. Both versatile nutrient and specific signal. Plant Physiol. 2004, 136, 2438–2442. [Google Scholar] [CrossRef]
- Weng, X.; Li, H.; Ren, C.; Zhou, Y.; Zhu, W.; Zhang, S.; Liu, L. Calcium regulates growth and nutrient absorption in poplar seedlings. Front. Plant Sci. 2022, 13, 887098. [Google Scholar] [CrossRef]
- Kitano, M.; Araki, T.; Yoshida, S.; Eguchi, H. Dependence of calcium uptake on water absorption and respiration in roots of tomato plants (Lycopersicon esculentum Mill.). Biotronics Environ. Control Environ. Biol. 1999, 28, 121–130. Available online: https://www.researchgate.net/publication/237406446 (accessed on 2 July 2025).
- Hewitt, E.J. The essential nutrient elements: Requirements and interactions. In Plant Physiology; Academic Press: Cambridge, MA, USA, 1963; pp. 137–360. [Google Scholar]
- White, P.J. The pathways of calcium movement to the xylem. J. Exp. Bot. 2001, 52, 891–899. [Google Scholar] [CrossRef]
- Ho, L.C.; White, P.J. A cellular hypothesis for the induction of blossom-end rot in tomato fruit. Ann. Bot. 2005, 95, 571–581. [Google Scholar] [CrossRef]
- Epstein, E. Mineral Nutrition of Plants: Principles and Perspectives; John Wiley & Sons: New York, NY, USA, 1972. [Google Scholar]
- Fuller, G.M.; Ellison, J.J.; McGill, M.; Sordahl, L.A.; Brinkley, B.R. Studies on the inhibitory role of calcium in the regulation of microtubule assembly in vitro and in vivo. In Microtubules and Microtubule Inhibitors; North-Holland Publishing Co.: Amsterdam, The Netherlands, 1975; pp. 379–390. [Google Scholar]
- Ninkuu, V.; Yan, J.; Fu, Z.; Yang, T.; Ziemah, J.; Ullrich, M.S.; Kuhnert, N.; Zeng, H. Lignin and its pathway-associated phytoalexins modulate plant defense against fungi. J. Fungi 2022, 9, 52. [Google Scholar] [CrossRef]
- Daher, F.B.; Braybrook, S.A. How to let go: Pectin and plant cell adhesion. Front. Plant Sci. 2015, 6, 523. [Google Scholar] [CrossRef]
- Wehr, J.B.; Menzies, N.W.; Blamey, F.P.C. Inhibition of cell-wall autolysis and pectin degradation by cations. Plant Physiol. Biochem. 2004, 42, 485–492. [Google Scholar] [CrossRef]
- Gutiérrez-Sánchez, A.; Plasencia, J.; Monribot-Villanueva, J.L.; Rodríguez-Haas, B.; Ruíz-May, E.; Guerrero-Analco, J.A.; Sánchez-Rangel, D. Virulence factors of the genus Fusarium with targets in plants. Microbiol. Res. 2023, 277, 127506. [Google Scholar] [CrossRef]
- Ralet, M.C.; Dronnet, V.; Buchholt, H.C.; Thibault, J.F. Enzymatically and chemically de-esterified lime pectins: Characterisation, polyelectrolyte behaviour and calcium binding properties. Carbohydr. Res. 2001, 336, 117–125. [Google Scholar] [CrossRef]
- Goulao, L.F.; Santos, J.; de Sousa, I.; Oliveira, C.M. Patterns of enzymatic activity of cell wall-modifying enzymes during growth and ripening of apples. Postharvest Biol. Technol. 2007, 43, 307–318. [Google Scholar] [CrossRef]
- Massiot, P.; Baron, A.; Drilleau, J.F. Characterisation and enzymatic hydrolysis of cell-wall polysaccharides from different tissue zones of apple. Carbohydr. Polym. 1994, 25, 145–154. [Google Scholar] [CrossRef]
- Wallace, A.; Frolich, E.; Lunt, O.R. Calcium requirements of higher plants. Nature 1966, 209, 634. [Google Scholar] [CrossRef]
- Lahaye, P.A.; Epstein, E. Calcium and salt toleration by bean plants. Physiol. Plant. 1971, 25, 213–218. [Google Scholar] [CrossRef]
- Asher, C.J.; Edwards, D.G. Modern solution culture techniques. In Inorganic Plant Nutrition; Springer: Berlin/Heidelberg, Germany, 1983; pp. 94–119. [Google Scholar] [CrossRef]
- Cramer, G.R. Sodium-calcium interactions under salinity stress. In Salinity: Environment-Plants-Molecules; Springer: Dordrecht, The Netherlands, 2002; pp. 205–227. [Google Scholar]
- Horst, W.J.; Wang, Y.; Eticha, D. The role of the root apoplast in aluminium-induced inhibition of root elongation and in aluminium resistance of plants: A review. Ann. Bot. 2010, 106, 185–197. [Google Scholar] [CrossRef]
- Kirkby, E.A.; Pilbeam, D.J. Calcium as a plant nutrient. Plant Cell Environ. 1984, 7, 397–405. [Google Scholar] [CrossRef]
- Lovegrove, A.; Hooley, R. Gibberellin and abscisic acid signalling in aleurone. Trends Plant Sci. 2000, 5, 102–110. [Google Scholar] [CrossRef]
- Bush, D.S.; Cornejo, M.J.; Huang, C.N.; Jones, R.L. Ca2+-stimulated secretion of α-amylase during development in barley aleurone protoplasts. Plant Physiol. 1986, 82, 566–574. [Google Scholar] [CrossRef]
- Luan, S.; Wang, C. Calcium signaling mechanisms across kingdoms. Annu. Rev. Cell Dev. Biol. 2021, 37, 311–340. [Google Scholar] [CrossRef]
- Naz, M.; Afzal, M.R.; Raza, M.A.; Pandey, S.; Qi, S.; Dai, Z.; Du, D. Calcium (Ca2+) signaling in plants: A plant stress perspective. South Afr. J. Bot. 2024, 169, 464–485. [Google Scholar] [CrossRef]
- Bu, Y.; Fu, W.; Chen, J.; Takano, T.; Liu, S. Description of AtCAX4 in response to abiotic stress in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 856. [Google Scholar] [CrossRef]
- Pirayesh, N.; Giridhar, M.; Khedher, A.B.; Vothknecht, U.C.; Chigri, F. Organellar calcium signaling in plants: An update. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2021, 1868, 118948. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Shankar, A.; Nalini Chandran, A.K.; Sharma, M.; Jung, K.H.; Suprasanna, P.; Pandey, G.K. Emerging concepts of potassium homeostasis in plants. J. Exp. Bot. 2020, 71, 608–619. [Google Scholar] [CrossRef]
- Xiao, Y.; Ma, C.; Li, M.; Zhangzhong, L.; Song, P.; Li, Y. Interaction and adaptation of phosphorus fertilizer and calcium ion in drip irrigation systems: The perspective of emitter clogging. Agric. Water Manag. 2023, 282, 108269. [Google Scholar] [CrossRef]
- Verma, S.; Negi, N.P.; Narwal, P.; Kumari, P.; Kisku, A.V.; Gahlot, P.; Mittal, N.; Kumar, D. Calcium signaling in coordinating plant development, circadian oscillations and environmental stress responses in plants. Environ. Exp. Bot. 2022, 201, 104935. [Google Scholar] [CrossRef]
- Ghosh, S.; Bheri, M.; Pandey, G.K. Delineating calcium signaling machinery in plants: Tapping the potential through functional genomics. Curr. Genom. 2021, 22, 404–439. [Google Scholar] [CrossRef]
- Fedrizzi, L.; Lim, D.; Carafoli, E. Calcium and signal transduction. Biochem. Mol. Biol. Educ. 2008, 36, 175–180. [Google Scholar] [CrossRef]
- Lecourieux, D.; Ranjeva, R.; Pugin, A. Calcium in plant defence-signalling pathways. New Phytol. 2006, 171, 249–269. [Google Scholar] [CrossRef]
- Paliyath, G.; Thompson, J.E. Calcium-and calmodulin-regulated breakdown of phospholipid by microsomal membranes from bean cotyledons. Plant Physiol. 1987, 83, 63–68. [Google Scholar] [CrossRef]
- Rudd, J.J.; Franklin-Tong, V.E. Calcium signaling in plants. Cell. Mol. Life Sci. 1999, 55, 214–232. [Google Scholar] [CrossRef]
- McAinsh, M.R.; Pittman, J.K. Shaping the calcium signature. New Phytol. 2009, 181, 275–294. [Google Scholar] [CrossRef]
- Putney, J.W., Jr. Calcium signaling: Up, down, up, down.... What’s the point? Science 1998, 279, 191–192. [Google Scholar] [CrossRef]
- Luan, S.; Kudla, J.; Rodriguez-Concepcion, M.; Yalovsky, S.; Gruissem, W. Calmodulins and calcineurin B–like proteins: Calcium sensors for specific signal response coupling in plants. Plant Cell 2002, 14 (Suppl. S1), S389–S400. [Google Scholar] [CrossRef]
- Sanders, D.; Pelloux, J.; Brownlee, C.; Harper, J.F. Calcium at the crossroads of signaling. Plant Cell 2002, 14 (Suppl. S1), S401–S417. [Google Scholar] [CrossRef]
- Bouché, N.; Scharlat, A.; Snedden, W.; Bouchez, D.; Fromm, H. A novel family of calmodulin-binding transcription activators in multicellular organisms. J. Biol. Chem. 2002, 277, 21851–21861. [Google Scholar] [CrossRef]
- Doherty, C.J.; Van Buskirk, H.A.; Myers, S.J.; Thomashow, M.F. Roles for Arabidopsis CAMTA transcription factors in cold-regulated gene expression and freezing tolerance. Plant Cell 2009, 21, 972–984. [Google Scholar] [CrossRef]
- Laohavisit, A.; Davies, J.M. Multifunctional annexins. Plant Sci. 2009, 177, 532–539. [Google Scholar] [CrossRef]
- Miedema, H.; Bothwell, J.H.; Brownlee, C.; Davies, J.M. Calcium uptake by plant cells–channels and pumps acting in concert. Trends Plant Sci. 2001, 6, 514–519. [Google Scholar] [CrossRef]
- White, P.J. Calcium channels in higher plants. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1465, 171–189. [Google Scholar] [CrossRef]
- White, P.J.; Bowen, H.C.; Demidchik, V.; Nichols, C.; Davies, J.M. Genes for calcium-permeable channels in the plasma membrane of plant root cells. Biochim. Biophys. Acta (BBA)-Biomembr. 2002, 1564, 299–309. [Google Scholar] [CrossRef]
- White, P.J. Depolarization-activated calcium channels shape the calcium signatures induced by low-temperature stress. New Phytol. 2009, 183, 6–8. Available online: http://www.jstor.org/stable/40302000 (accessed on 2 July 2025). [CrossRef]
- Miedema, H.; Demidchik, V.; Véry, A.A.; Bothwell, J.H.; Brownlee, C.; Davies, J.M. Two voltage-dependent calcium channels co-exist in the apical plasma membrane of Arabidopsis thaliana root hairs. New Phytol. 2008, 179, 378–385. [Google Scholar] [CrossRef]
- Amtmann, A.; Blatt, M.R. Regulation of macronutrient transport. New Phytol. 2009, 181, 35. [Google Scholar] [CrossRef]
- Kim, M.D.; Kim, Y.H.; Kwon, S.Y.; Yun, D.J.; Kwak, S.S.; Lee, H.S. Enhanced tolerance to methyl viologen-induced oxidative stress and high temperature in transgenic potato plants overexpressing the CuZnSOD, APX and NDPK2 genes. Physiol. Plant. 2010, 140, 153–162. [Google Scholar] [CrossRef]
- Moran, N. Osmoregulation of leaf motor cells. FEBS Lett. 2007, 581, 2337–2347. [Google Scholar] [CrossRef]
- Harper, J.F. Dissecting calcium oscillators in plant cells. Trends Plant Sci. 2001, 6, 395–397. [Google Scholar] [CrossRef]
- Klüsener, B.; Boheim, G.; Liss, H.; Engelberth, J.; Weiler, E.W. Gadolinium-sensitive, voltage-dependent calcium release channels in the endoplasmic reticulum of a higher plant mechanoreceptor organ. EMBO J. 1995, 14, 2708–2714. [Google Scholar] [CrossRef]
- Hirschi, K.D. Vacuolar H+/Ca2+ transport: Who’s directing the traffic? Trends Plant Sci. 2001, 6, 100–104. [Google Scholar] [CrossRef]
- Wu, Z.; Liang, F.; Hong, B.; Young, J.C.; Sussman, M.R.; Harper, J.F.; Sze, H. An endoplasmic reticulum-bound Ca2+/Mn2+ pump, ECA1, supports plant growth and confers tolerance to Mn2+ stress. Plant Physiol. 2002, 130, 128–137. [Google Scholar] [CrossRef]
- Blatt, M.R. Cellular signaling and volume control in stomatal movements in plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 221–241. [Google Scholar] [CrossRef]
- Ritchie, S.M.; Swanson, S.J.; Gilroy, S. From common signalling components to cell-specific responses: Insights from the cereal aleurone. Physiol. Plant. 2002, 115, 342–351. [Google Scholar] [CrossRef]
- Evans, D.E.; Williams, L.E. P-type calcium ATPases in higher plants–biochemical, molecular and functional properties. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 1998, 1376, 1–25. [Google Scholar] [CrossRef]
- Huang, L.; Berkelman, T.; Franklin, A.E.; Hoffman, N.E. Characterization of a gene encoding a Ca (2+)-ATPase-like protein in the plastid envelope. Proc. Natl. Acad. Sci. USA 1993, 90, 10066–10070. [Google Scholar] [CrossRef]
- Harper, J.F.; Hong, B.; Hwang, I.; Guo, H.Q.; Stoddard, R.; Huang, J.F.; Palmgren, M.G.; Sze, H. A novel calmodulin-regulated Ca2+-ATPase (ACA2) from Arabidopsis with an N-terminal autoinhibitory domain. J. Biol. Chem. 1998, 273, 1099–1106. [Google Scholar] [CrossRef]
- Bonza, M.C.; Morandini, P.; Luoni, L.; Geisler, M.; Palmgren, M.G.; De Michelis, M.I. At-ACA8 encodes a plasma membrane-localized calcium-ATPase of Arabidopsis with a calmodulin-binding domain at the N terminus. Plant Physiol. 2000, 123, 1495–1506. [Google Scholar] [CrossRef]
- George, L.; Romanowsky, S.M.; Harper, J.F.; Sharrock, R.A. The ACA10 Ca2+-ATPase regulates adult vegetative development and inflorescence architecture in Arabidopsis. Plant Physiol. 2008, 146, 716. [Google Scholar] [CrossRef]
- Schiøtt, M.; Romanowsky, S.M.; Bækgaard, L.; Jakobsen, M.K.; Palmgren, M.G.; Harper, J.F. A plant plasma membrane Ca2+ pump is required for normal pollen tube growth and fertilization. Proc. Natl. Acad. Sci. USA 2004, 101, 9502–9507. [Google Scholar] [CrossRef] [PubMed]
- Geisler, M.; Frangne, N.; Gomes, E.; Martinoia, E.; Palmgren, M.G. The ACA4 gene of Arabidopsis encodes a vacuolar membrane calcium pump that improves salt tolerance in yeast. Plant Physiol. 2000, 124, 1814–1827. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Kim, B.G.; Cheong, Y.H.; Pandey, G.K.; Luan, S. A Ca2+ signaling pathway regulates a K+ channel for low-K response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 12625–12630. [Google Scholar] [CrossRef]
- Liang, F.; Cunningham, K.W.; Harper, J.F.; Sze, H. ECA1 complements yeast mutants defective in Ca2+ pumps and encodes an endoplasmic reticulum-type Ca2+-ATPase in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1997, 94, 8579–8584. [Google Scholar] [CrossRef] [PubMed]
- Mills, R.F.; Doherty, M.L.; López-Marqués, R.L.; Weimar, T.; Dupree, P.; Palmgren, M.G.; Pittman, J.K.; Williams, L.E. ECA3, a Golgi-localized P2A-type ATPase, plays a crucial role in manganese nutrition in Arabidopsis. Plant Physiol. 2008, 146, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chanroj, S.; Wu, Z.; Romanowsky, S.M.; Harper, J.F.; Sze, H. A distinct endosomal Ca2+/Mn2+ pump affects root growth through the secretory process. Plant Physiol. 2008, 147, 1675–1689. [Google Scholar] [CrossRef]
- Cheng, N.H.; Pittman, J.K.; Shigaki, T.; Lachmansingh, J.; LeClere, S.; Lahner, B.; Salt, D.E.; Hirschi, K.D. Functional association of Arabidopsis CAX1 and CAX3 is required for normal growth and ion homeostasis. Plant Physiol. 2005, 138, 2048–2060. [Google Scholar] [CrossRef]
- Hirschi, K.D.; Korenkov, V.D.; Wilganowski, N.L.; Wagner, G.J. Expression of Arabidopsis CAX2 in tobacco. Altered metal accumulation and increased manganese tolerance. Plant Physiol. 2000, 124, 125–134. [Google Scholar] [CrossRef]
- Luo, G.Z.; Wang, H.W.; Huang, J.; Tian, A.G.; Wang, Y.J.; Zhang, J.S.; Chen, S.Y. A putative plasma membrane cation/proton antiporter from soybean confers salt tolerance in Arabidopsis. Plant Mol. Biol. 2005, 59, 809–820. [Google Scholar] [CrossRef]
- Hepler, P.K.; Winship, L.J. Calcium at the cell wall-cytoplast interface. J. Integr. Plant Biol. 2010, 52, 147–160. [Google Scholar] [CrossRef]
- De Kreij, C. Interactive effects of air humidity, calcium and phosphate on blossom-end rot, leaf deformation, production and nutrient contents of tomato. J. Plant Nutr. 1996, 19, 361–377. [Google Scholar] [CrossRef]
- Clarkson, D.T. Roots and the delivery of solutes to the xylem. Philosophical Transactions of the Royal Society of London. Ser. B Biol. Sci. 1993, 341, 5–17. [Google Scholar] [CrossRef]
- Moore, C.A.; Bowen, H.C.; Scrase-Field, S.; Knight, M.R.; White, P.J. The deposition of suberin lamellae determines the magnitude of cytosolic Ca2+ elevations in root endodermal cells subjected to cooling. Plant J. 2002, 30, 457–465. [Google Scholar] [CrossRef]
- Park, S.; Waterland, N.L. Evaluation of calcium application methods on delaying plant wilting under water deficit in bedding plants. Agronomy 2021, 11, 1383. [Google Scholar] [CrossRef]
- Santos, E.; Montanha, G.S.; Agostinho, L.F.; Polezi, S.; Marques, J.P.R.; de Carvalho, H.W.P. Foliar calcium absorption by tomato plants: Comparing the effects of calcium sources and adjuvant usage. Plants 2023, 12, 2587. [Google Scholar] [CrossRef]
- Haleema, B.; Shah, S.T.; Basit, A.; Hikal, W.M.; Arif, M.; Khan, W.; Said-Al Ahl, H.A.; Fhatuwani, M. Comparative Effects of Calcium, Boron, and Zinc Inhibiting Physiological Disorders, Improving Yield and Quality of Solanum lycopersicum. Biology 2024, 13, 766. [Google Scholar] [CrossRef]
- Li, Y.; Ma, J.; Gao, X.; Tie, J.; Wu, Y.; Tang, Z.; Hu, L.; Yu, J. Exogenous brassinosteroids alleviate calcium deficiency-induced tip-burn by maintaining cell wall structural stability and higher photosynthesis in mini Chinese Cabbage. Front. Plant Sci. 2022, 13, 999051. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ma, J.; Zhang, W.; Gao, X.; Wang, X.; Chen, W.; Dawuda, M.M.; Li, W.; Hu, L. Quality Response of Two Mini Chinese Cabbage Cultivars to Different Calcium Levels. Foods 2025, 14, 872. [Google Scholar] [CrossRef]
- Geraldson, C.M. Studies on control of blackheart of celery. Proc. Fla. State Hortic. Soc. 1952, 65, 171–172. [Google Scholar]
- Vergara, C.; Araujo, K.E.C.; Santos, A.P.; de Oliveira, F.F.; de Souza Silva, G.; de Oliveira Miranda, N.; de Souza Barreto, E.; da Silva, I.K.; de Medeiros, J.F. Use of crushed eggshell to control tomato blossom-end rot. Res. Soc. Dev. 2024, 13, e2213545667. [Google Scholar] [CrossRef]
- Geraldson, C.M. The control of blackheart of celery. Proc. Am. Soc. Hortic. Sci. 1954, 63, 353–358. [Google Scholar]
- Kleemann, M. Effects of salinity, nutrients and spraying with cacl2 solution on the development of calcium deficiency in chervil (Anthriscus cerefolium (L.) Hoffm.) and curled parsley (Petroselinum crispum (Mill.) nym. Convar. Crispum). In Integrated View of Fruit and Vegetable Quality; CRC Press: Boca Raton, FL, USA, 2000; pp. 41–53. [Google Scholar]
- Schmitz-Eiberger, M.; Haefs, R.; Noga, G. Calcium deficiency-influence on the antioxidative defense system in tomato plants. J. Plant Physiol. 2002, 159, 733–742. [Google Scholar] [CrossRef]
- Parađiković, N.; Lončarić, Z.; Bertić, B.; Vukadinović, V. Influence of Ca-foliar application on yield and quality of sweet pepper in glasshouse conditions. Poljoprivreda 2004, 10, 24–27. Available online: https://www.researchgate.net/publication/47727972 (accessed on 31 May 2025).
- Raven, J.A. H+ and Ca2+ in phloem and symplast: Relation of relative immobility of the ions to the cytoplasmic nature of the transport paths. New Phytol. 1977, 79, 465–480. [Google Scholar] [CrossRef]
- Kohl, W. Die calciumverteilung in apfeln und ihre veranderung wahrend des wachstums. Die Gartenbauwissenschaft 1966, 31, 513–547. [Google Scholar]
- Perring, M.A.; Jackson, C.H. The mineral composition of apples. Calcium concentration and bitter pit in relation to mean mass per apple. J. Sci. Food Agric. 1975, 26, 1493–1502. [Google Scholar] [CrossRef]
- Bangerth, F. Calcium-related physiological disorders of plants. Annu. Rev. Phytopathol. 1979, 17, 97–122. [Google Scholar] [CrossRef]
- Chamel, A.R. Permeability characteristics of isolated ‘Golden Delicious’ apple fruit cuticles with regard to calcium. J. Am. Soc. Hortic. Sci. 1989, 114, 804–809. [Google Scholar] [CrossRef]
- Glenn, G.M.; Poovaiah, B.W. Cuticular permeability to calcium compounds in ‘Golden Delicious’ apple fruit. J. Am. Soc. Hortic. Sci. 1985, 110, 192–195. [Google Scholar] [CrossRef]
- Schlegel, T.K.; Schönherr, J. Penetration of calcium chloride into apple fruits as affected by stage of fruit development. Acta Hortic. 2002, 594, 527–533. [Google Scholar] [CrossRef]
- Wojcik, P. Effect of calcium chloride sprays at different water volumes on “Szampion” apple calcium concentration. J. Plant Nutr. 2001, 24, 639–650. [Google Scholar] [CrossRef]
- Sarijan, A.; Limbongan, A.A.; Ekowati, N.Y.; Kusumah, R. Calcium nitrate application on tomatoes to increase blossom-end rot disease resistance. IOP Conf. Ser. Earth Environ. Sci. 2025, 1471, 012025. [Google Scholar] [CrossRef]
- Assaha, D.V.; Petang, L.Y. Bone meal enhances the growth and yield of the tomato cultivar Cobra F1 by increasing fruit Ca content and alleviating blossom-end rot. Asian J. Agric. 2024, 8, 1–25. [Google Scholar] [CrossRef]
- Madani, B.; Mirshekari, A.; Sofo, A.; Tengku Muda Mohamed, M. Preharvest calcium applications improve postharvest quality of papaya fruits (Carica papaya L. cv. Eksotika II). J. Plant Nutr. 2016, 39, 1483–1492. [Google Scholar] [CrossRef]
- Parsa, Z.; Roozbehi, S.; Hosseinifarahi, M.; Radi, M.; Amiri, S. Integration of pomegranate peel extract (PPE) with calcium sulphate (CaSO4): A friendly treatment for extending shelf-life and maintaining postharvest quality of sweet cherry fruit. J. Food Process. Preserv. 2021, 45, e15089. [Google Scholar] [CrossRef]
- Zhai, J.; Gao, Y.; Zhang, X.W.; Han, L.J.; Bi, H.A.; Li, Q.M.; Ai, X. Effects of silicon and calcium on photosynthesis, yield and quality of cucumber in solar-greenhouse. Acta Hortic. Sin. 2019, 46, 701–713. [Google Scholar] [CrossRef]
- Ahn, J.; Park, M.; Kim, J.; Huq, E.; Kim, J.; Kim, D.H. Physiological and transcriptomic analyses of healthy and blossom-end-rot (BER)-defected fruit of chili pepper (Capsicum annuum. L.). Hortic. Environ. Biotechnol. 2024, 65, 971–980. [Google Scholar] [CrossRef]
- Ferguson, I.B.; Watkins, C.B. Bitter pit in apple fruit. Hortic. Rev. 1989, 11, 289–355. [Google Scholar] [CrossRef]
- Ferguson, I.B.; Watkins, C.B. Cation distribution and balance in apple fruit in relation to calcium treatments for bitter pit. Sci. Hortic. 1983, 19, 301–310. [Google Scholar] [CrossRef]
- Huang, Z.; Takemoto, T.; Omwange, K.A.; Orsino, M.; Konagaya, K.; Kondo, N. Early detection of Blossom-End Rot in green peppers using fluorescence and normal color images in visible region. Food Control 2025, 172, 111156. [Google Scholar] [CrossRef]
- Karley, A.J.; White, P.J. Moving cationic minerals to edible tissues: Potassium, magnesium, calcium. Curr. Opin. Plant Biol. 2009, 12, 291–298. [Google Scholar] [CrossRef]
- Gilliham, M.; Dayod, M.; Hocking, B.J.; Xu, B.; Conn, S.J.; Kaiser, B.N.; Leigh, R.A.; Tyerman, S.D. Calcium delivery and storage in plant leaves: Exploring the link with water flow. J. Exp. Bot. 2011, 62, 2233–2250. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, S.T.; Shackel, K.A.; Mitcham, E.J. Abscisic acid triggers whole-plant and fruit-specific mechanisms to increase fruit calcium uptake and prevent blossom end rot development in tomato fruit. J. Exp. Bot. 2011, 62, 2645–2656. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.C. Environmental effects on the diurnal accumulation of 45Ca by young fruit and leaves of tomato plants. Ann. Bot. 1989, 63, 281–288. [Google Scholar] [CrossRef]
- Ho, L.C.; Belda, R.; Brown, M.; Andrews, J.; Adams, P. Uptake and transport of calcium and the possible causes of blossom-end rot in tomato. J. Exp. Bot. 1993, 44, 509–518. [Google Scholar] [CrossRef]
- Tadesse, T.; Nichols, M.A.; Hewett, E.W.; Fisher, K.J. Relative humidity around the fruit influences the mineral composition and incidence of blossom-end rot in sweet pepper fruit. J. Hortic. Sci. Biotechnol. 2001, 76, 9–16. [Google Scholar] [CrossRef]
- Fischer, A.M. Nutrient remobilization during leaf senescence. Annu. Rev. Senescence Process. Plants 2007, 26, 87–107. [Google Scholar]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2011; ISBN 9780123849052. [Google Scholar]
- Song, W.; Yi, J.; Kurniadinata, O.F.; Wang, H.; Huang, X. Linking fruit Ca uptake capacity to fruit growth and pedicel anatomy, a cross-species study. Front. Plant Sci. 2018, 9, 575. [Google Scholar] [CrossRef]
- Bar-Tal, A.; Aloni, B.; Karni, L.; Oserovitz, J.; Hazan, A.; Itach, M.; Gantz, S.; Avidan, A.; Posalski, I.; Tratkovski, N.; et al. Nitrogen nutrition of greenhouse pepper. I. Effects of nitrogen concentration and NO3: NH4 ratio on yield, fruit shape, and the incidence of blossom-end rot in relation to plant mineral composition. HortScience 2001, 36, 1244–1251. [Google Scholar] [CrossRef]
- Schönherr, J.; Bukovac, M.J. Ion exchange properties of isolated tomato fruit cuticular membrane: Exchange capacity, nature of fixed charges and cation selectivity. Planta 1973, 109, 73–93. [Google Scholar] [CrossRef]
- Yermiyahu, U.; Nir, S.; Ben-Hayyim, G.; Kafkafi, U. Quantitative competition of calcium with sodium or magnesium for sorption sites on plasma membrane vesicles of melon (Cucumis melo L.) root cells. J. Membr. Biol. 1994, 138, 55–63. [Google Scholar] [CrossRef]
- De Freitas, S.T.; do Amarante, C.V.; Labavitch, J.M.; Mitcham, E.J. Cellular approach to understand bitter pit development in apple fruit. Postharvest Biol. Technol. 2010, 57, 6–13. [Google Scholar] [CrossRef]
- Dris, R.; Niskanen, R.; Fallahi, E. Nitrogen and calcium nutrition and fruit quality of commercial apple cultivars grown in Finland. J. Plant Nutr. 1998, 21, 2389–2402. [Google Scholar] [CrossRef]
- Lanauskas, J.; Kvikliene, N. Effect of calcium foliar application on some fruit quality characteristics of ‘Sinap Orlovskij’apple. Agron. Res. 2006, 4, 31–36. [Google Scholar]
- Park, S.; Cheng, N.H.; Pittman, J.K.; Yoo, K.S.; Park, J.; Smith, R.H.; Hirschi, K.D. Increased calcium levels and prolonged shelf life in tomatoes expressing Arabidopsis H+/Ca2+ transporters. Plant Physiol. 2005, 139, 1194–1206. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, S.T.; Mitcham, E.I. 3 factors involved in fruit calcium deficiency disorders. Hortic. Rev. 2012, 40, 107–146. [Google Scholar] [CrossRef]
- Picchioni, G.A.; Watada, A.E.; Conway, W.S.; Whitaker, B.D.; Sams, C.E. Postharvest calcium infiltration delays membrane lipid catabolism in apple fruit. J. Agric. Food Chem. 1998, 46, 2452–2457. [Google Scholar] [CrossRef]
- Lund, Z.F. The effect of calcium and its relation to several cations in soybean root growth. Soil Sci. Soc. Am. J. 1970, 34, 456–459. [Google Scholar] [CrossRef]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef]
- Bush, D.S. Calcium regulation in plant cells and its role in signaling. Annu. Rev. Plant Biol. 1995, 46, 95–122. [Google Scholar] [CrossRef]
- Kendall, J.M.; Dormer, R.L.; Campbell, A.K. Targeting aequorin to the endoplasmic reticulum of living cells. Biochem. Biophys. Res. Commun. 1992, 189, 1008–1016. [Google Scholar] [CrossRef]
- Johnson, C.H.; Knight, M.R.; Kondo, T.; Masson, P.; Sedbrook, J.; Haley, A.; Trewavas, A. Circadian oscillations of cytosolic and chloroplastic free calcium in plants. Science 1995, 269, 1863–1865. [Google Scholar] [CrossRef] [PubMed]
- Logan, D.C.; Knight, M.R. Mitochondrial and cytosolic calcium dynamics are differentially regulated in plants. Plant Physiol. 2003, 133, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Pauly, N.; Knight, M.R.; Thuleau, P.; Graziana, A.; Muto, S.; Ranjeva, R.; Mazars, C. The nucleus together with the cytosol generates patterns of specific cellular calcium signatures in tobacco suspension culture cells. Cell Calcium 2001, 30, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Voyle, G. Blossom End Rot Causes and Cures in Garden Vegetables; Michigan State University: Extension, MI, USA, 2021. [Google Scholar]
- de Freitas, S.T.; Amarante, C.D.; Mitcham, E.J. Calcium deficiency disorders in plants. In Postharvest Ripening Physiology of Crops; CRC Press: Boca Raton, FL, USA, 2016; pp. 477–502. ISBN 9781498703819. [Google Scholar]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit calcium: Transport and physiology. Front. Plant Sci. 2016, 7, 569. [Google Scholar] [CrossRef]
- Ren, H.; Zhao, X.; Li, W.; Hussain, J.; Qi, G.; Liu, S. Calcium signaling in plant programmed cell death. Cells 2021, 10, 1089. [Google Scholar] [CrossRef]
- Ojha, R.K.; Jha, S.K. Role of mineral nutrition in management of plant diseases. In Farmers’ Prosperity Through Improved Agricultural Technologies; Singh, H.K., Solankey, S.S., Roy, M.K., Eds.; Jaya Publishing House: Delhi, India, 2021; pp. 241–261. Available online: https://www.researchgate.net/publication/348448043_ (accessed on 2 July 2025).
- Kleemann, M. Development of calcium deficiency symptoms in chervil (Antriscus cerefolium (L.) Hoffm.) and curled parsley (Petroselinum crispum (Mill.) Nym. convar crispum). Zesz. Probl. Postępów Nauk. Rol. 1999, 468, 335–348. [Google Scholar]
- Aloni, B. Enhancement of leaf tip burn by restricting root growth in Chinese cabbage plants. J. Hortic. Sci. 1986, 61, 509–513. [Google Scholar] [CrossRef]
- Collier, G.F.; Tibbitts, T.W. Tipburn of lettuce. Hortic. Rev. 1982, 4, 49–65. [Google Scholar] [CrossRef]
- Cox, E.F.; McKee, J.M.T.; Dearman, A.S. The effect of growth rate on tipburn occurrence in lettuce. J. Hortic. Sci. 1976, 51, 297–309. [Google Scholar] [CrossRef]
- Bible, B.B.; Stiehl, B. Effect of atmospheric modification on the incidence of blackheart and the cation content of celery. Sci. Hortic. 1986, 28, 19–28. [Google Scholar] [CrossRef]
- De Freitas, S.T.; Martinelli, F.; Feng, B.; Reitz, N.F.; Mitcham, E.J. Transcriptome approach to understand the potential mechanisms inhibiting or triggering blossom-end rot development in tomato fruit in response to plant growth regulators. J. Plant Growth Regul. 2018, 37, 183–198. [Google Scholar] [CrossRef]
- Adams, P.; Holder, R. Effects of humidity, Ca and salinity on the accumulation of dry matter and Ca by the leaves and fruit of tomato (Lycopersicon esculentum). J. Hortic. Sci. 1992, 67, 137–142. [Google Scholar] [CrossRef]
- De Kreij, C.; Janse, J.; Van Goor, B.J.; Van Doesburg, J.D.J. The incidence of calcium oxalate crystals in fruit walls of tomato (Lycopersicon esculentum Mill.) as affected by humidity, phosphate and calcium supply. J. Hortic. Sci. 1992, 67, 45–50. [Google Scholar] [CrossRef]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1987; 687p, ISBN 978-1-4020-0008-9/978-9-40-101009-2. [Google Scholar]
- Kabir, M.Y. Abiotic Factors Affecting Plant Physiology and Fruit Yield and Physiological Disorders in Bell Pepper and Tomato. Ph.D. Thesis, University of Georgia, Athens, GA, USA, 2019; 219p. [Google Scholar]
- Horst, W.J. Aluminium tolerance and calcium efficiency of cowpea genotypes. J. Plant Nutr. 1987, 10, 1121–1129. [Google Scholar] [CrossRef]
- Ho, L.C.; Hand, D.J.; Fussell, M. Improvement of tomato fruit quality by calcium nutrition. Int. Symp. Grow. Media Hydroponics 1997, 481, 463–468. [Google Scholar] [CrossRef]
- Ho, L.C.; Adams, P.; Li, X.Z.; Shen, H.; Andrews, J.; Xu, Z.H. Responses of Ca-efficient and Ca-inefficient tomato cultivars to salinity in plant growth, calcium accumulation and blossom-end rot. J. Hortic. Sci. 1995, 70, 909–918. [Google Scholar] [CrossRef]
- Adams, P.; Ho, L.C. The susceptibility of modern tomato cultivars to blossom-end rot in relation to salinity. J. Hortic. Sci. 1992, 67, 827–839. [Google Scholar] [CrossRef]
- Cardoso, M.O.; Queiróz, M.D.; Souza, R.F. Incidence of blossom-end rot in five tomato cultivars grown in soil with three calcium levels. Hortic. Bras. 1995, 13, 172–175. [Google Scholar]
- Sperry, W.J.; Davis, J.M.; Sanders, D.C. Soil moisture and cultivar influence cracking, blossom-end rot, zippers, and yield of staked fresh-market tomatoes. HortTechnology 1996, 6, 21–24. [Google Scholar] [CrossRef]
- Maynard, D.N.; Barham, W.S.; McCombs, C.L. The effect of calcium nutrition of tomatoes as related to the incidence and severity of blossom-end rot. Proc. Amer. Soc. Hort. Sci 1957, 69, 318–322. [Google Scholar]
- Magán, J.J.; Gallardo, M.; Thompson, R.B.; Lorenzo, P. Effects of salinity on fruit yield and quality of tomato grown in soil-less culture in greenhouses in Mediterranean climatic conditions. Agric. Water Manag. 2008, 95, 1041–1055. [Google Scholar] [CrossRef]
- Srivastava, S.; Singh, L.; Debnath, B.; Shukla, M.; Kumar, S.; Rana, M.; Meshram, S. Blossom end-rot (BER): An abiotic devastating disorder of tomato. Clim. Change Environ. Sustain. 2022, 10, 15–22. [Google Scholar] [CrossRef]
- Kumar, V.; Darvhankar, M.; Srivastava, S. Evaluation of Trichoderma species and fungicides on nutritional quality and yield of tomato. Pl. Cell Biotechnol. Mol. Biol 2019, 20, 331–338. [Google Scholar]
- Das, K.A.; Bhosale, R.K.; Chavan, S.S.; Rana, M.; Srivastava, S. Effect of physiological factors and different growth media on Alternaria solani under in vitro condition and eco-friendly management of early blight of tomato (Solanum lycopersicum L.). Ann Biol 2023, 39, 66–72. [Google Scholar]
- Zhou, L.; Fu, Y.; Yang, Z. A genome-wide functional characterization of Arabidopsis regulatory calcium sensors in pollen tubes. J. Integr. Plant Biol. 2009, 51, 751–761. [Google Scholar] [CrossRef]
- Yoshida, Y.; Irie, N.; Vinh, T.D.; Ooyama, M.; Tanaka, Y.; Yasuba, K.I.; Goto, T. Incidence of blossom-end rot in relation to the water-soluble calcium concentration in tomato fruits as affected by calcium nutrition and cropping season. J. Jpn. Soc. Hortic. Sci. 2014, 83, 282–289. [Google Scholar] [CrossRef]
- Barickman, T.C.; Kopsell, D.A.; Sams, C.E. Abscisic acid increases carotenoid and chlorophyll concentrations in leaves and fruit of two tomato genotypes. J. Am. Soc. Hortic. Sci. 2014, 139, 261–266. [Google Scholar] [CrossRef]
- R’him, T.; Jebari, H. Blossom-end rot in relation to morphological parameters and calcium content in fruits of four pepper varieties (Capsicum annuum L.). Biotechnol. Agron. Société Environ. 2008, 12, 361–366. [Google Scholar]
- Grasselly, D.; Rosso, L.; Holgard, S.; Cottet, V.; Navez, B.; Jost, M.; Berier, A. Soilless culture of tomato: Effect of salinity of the nutrient solution. Infos-Ctifl 2008, 239, 41–45. [Google Scholar]
- Robbins, W.R. Relation of nutrient salt concentration to growth of the tomato and to the incidence of blossom-end rot of the fruit. Plant Physiol. 1937, 12, 21. [Google Scholar] [CrossRef]
- Dekock, P.C.; Hall, A.; Inkson, R.H.; Alan Robertson, R. Blossom-end rot in tomatoes. J. Sci. Food Agric. 1979, 30, 508–514. [Google Scholar] [CrossRef]
- Adams, P.; Ho, L.C. Effects of environment on the uptake and distribution of calcium in tomato and on the incidence of blossom-end rot. Plant Soil 1993, 154, 127–132. [Google Scholar] [CrossRef]
- Pill, W.G.; Lambeth, V.N. Effects of soil water regime and nitrogen form on blossom-end rot, yield, water relations, and elemental composition of tomato. J. Am. Soc. Hortic. Sci. 1981, 105, 730–734. [Google Scholar] [CrossRef]
- Paiva, E.A.S.; Sampaio, R.A.; Martinez, H.E.P. Composition and quality of tomato fruit cultivated in nutrient solutions containing different calcium concentrations. J. Plant Nutr. 1998, 21, 2653–2661. [Google Scholar] [CrossRef]
- Van Goor, B.J. Influence of restricted water supply on blossom-end rot and ionic composition of tomatoes grown in nutrient solution. Commun. Soil Sci. Plant Anal. 1974, 5, 13–24. [Google Scholar] [CrossRef]
- Franco, J.A.; Perez-Saura, P.J.; Ferná Ndez, J.A.; Parra, M.; Garcia, A.L. Effect of two irrigation rates on yield, incidence of blossom-end rot, mineral content and free amino acid levels in tomato cultivated under drip irrigation using saline water. J. Hortic. Sci. Biotechnol. 1999, 74, 430–435. [Google Scholar] [CrossRef]
- Franco, J.; Bañón, S.; Madrid, R. Effects of a protein hydrolysate applied by fertigation on the effectiveness of calcium as a corrector of blossom-end rot in tomato cultivated under saline conditions. Sci. Hortic. 1994, 57, 283–292. [Google Scholar] [CrossRef]
- Marcelis, L.F.M.; Ho, L.C. Blossom-end rot in relation to growth rate and calcium content in fruits of sweet pepper (Capsicum annuum L.). J. Exp. Bot. 1999, 50, 357–363. [Google Scholar] [CrossRef]
- Chiu, T.F.; Bould, C. Effects of shortage of calcium and other cations on 45Ca mobility, growth and nutritional disorders of tomato plants (Lycopersicon esculentum). J. Sci. Food Agric. 1976, 27, 969–977. [Google Scholar] [CrossRef]
- Saure, M.C. Why calcium deficiency is not the cause of blossom-end rot in tomato and pepper fruit—A reappraisal. Sci. Hortic. 2014, 174, 151–154. [Google Scholar] [CrossRef]
- Gent, M.P. Effect of degree and duration of shade on quality of greenhouse tomato. HortScience 2007, 42, 514–520. [Google Scholar] [CrossRef]
- Díaz-Perez, J.C. Bell pepper (Capsicum annum L.) crop as affected by shade level: Fruit yield, quality, and postharvest attributes, and incidence of phytophthora blight (caused by Phytophthora capsici Leon.). HortScience 2014, 49, 891–900. [Google Scholar] [CrossRef]
- Kabir, M.Y.; Díaz-Pérez, J.C.; Nambeesan, S.U. Effect of shade levels on plant growth, physiology, and fruit yield in bell pepper (Capsicum annuum L.). In Proceedings of the XI International Symposium on Protected Cultivation in Mild Winter Climates and I International Symposium on Nettings and 1268, Tenerife, Canary Islands, Spain, 27 January 2020; pp. 311–318. [Google Scholar] [CrossRef]
- Kabir, M.Y.; Nambeesan, S.U.; Bautista, J.; Díaz-Pérez, J.C. Plant water status, plant growth, and fruit yield in bell pepper (Capsicum annum L.) under shade nets. Sci. Hortic. 2022, 303, 111241. [Google Scholar] [CrossRef]
- Kabir, M.Y.; Nambeesan, S.U.; Díaz-Pérez, J.C. Shade nets improve vegetable performance. Sci. Hortic. 2024, 334, 113326. [Google Scholar] [CrossRef]
- Jing, T.; Li, J.; He, Y.; Shankar, A.; Saxena, A.; Tiwari, A.; Maturi, K.C.; Solanki, M.K.; Singh, V.; Eissa, M.A.; et al. Role of calcium nutrition in plant Physiology: Advances in research and insights into acidic soil conditions—A comprehensive review. Plant Physiol. Biochem. 2024, 210, 108602. [Google Scholar] [CrossRef]
- Topcu, Y.; Nambeesan, S.U.; van der Knaap, E. Blossom-end rot: A century-old problem in tomato (Solanum lycopersicum L.) and other vegetables. Mol. Hortic. 2022, 2, 1. [Google Scholar] [CrossRef]
- Rezvi, H.U.A.; Tahjib-Ul-Arif, M.; Azim, M.A.; Tumpa, T.A.; Tipu, M.M.H.; Najnine, F.; Dawood, M.F.; Skalicky, M.; Brestič, M. Rice and food security: Climate change implications and the future prospects for nutritional security. Food Energy Secur. 2023, 12, e430. [Google Scholar] [CrossRef]
- Taylor, M.D.; Locascio, S.J.; Alligood, M.R. Blossom-end rot incidence of tomato as affected by irrigation quantity, calcium source, and reduced potassium. HortScience 2004, 39, 1110–1115. [Google Scholar] [CrossRef]
- Lens, F.; Gleason, S.M.; Bortolami, G.; Brodersen, C.; Delzon, S.; Jansen, S. Functional xylem characteristics associated with drought-induced embolism in angiosperms. New Phytol. 2022, 236, 2019–2036. [Google Scholar] [CrossRef]
- Oishy, M.N.; Shemonty, N.A.; Fatema, S.I.; Mahbub, S.; Mim, E.L.; Raisa, M.B.H.; Anik, A.H. Unravelling the effects of climate change on the soil-plant-atmosphere interactions: A critical review. Soil Environ. Health 2025, 3, 100130. [Google Scholar] [CrossRef]
- Taub, D. Effects of rising atmospheric concentrations of carbon dioxide on plants. Nat. Educ. Knowl. 2010, 3, 21. [Google Scholar]
- Ebi, K.L.; Loladze, I. Elevated atmospheric CO2 concentrations and climate change will affect our food’s quality and quantity. Lancet Planet. Health 2019, 3, e283–e284. [Google Scholar] [CrossRef]
- Wang, P.; Lombi, E.; Zhao, F.J.; Kopittke, P.M. Nanotechnology: A new opportunity in plant sciences. Trends Plant Sci. 2016, 21, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Usman, M.; Farooq, M.; Wakeel, A.; Nawaz, A.; Rehman, H.U.; Cheema, S.A. Nanotechnology in agriculture: Current status, challenges and future opportunities. Sci. Total Environ. 2021, 780, 146467. [Google Scholar] [CrossRef]
- Carmona, F.J.; Guagliardi, A.; Masciocchi, N. Nanosized calcium phosphates as novel macronutrient nano-fertilizers. Nanomaterials 2022, 12, 2709. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chen, S.; Li, Y.; Shi, Y. Effect of nano-calcium carbonate on morphology, antioxidant enzyme activity and photosynthetic parameters of wheat (Triticum aestivum L.) seedlings. Chem. Biol. Technol. Agric. 2023, 10, 31. [Google Scholar] [CrossRef]
- Demidchik, V.; Shabala, S.; Isayenkov, S.; Cuin, T.A.; Pottosin, I. Calcium transport across plant membranes: Mechanisms and functions. New Phytol. 2018, 220, 49–69. [Google Scholar] [CrossRef]
- Lowder, L.G.; Zhang, D.; Baltes, N.J.; Paul, J.W., III; Tang, X.; Zheng, X.; Voytas, D.F.; Hsieh, T.F.; Zhang, Y.; Qi, Y. A CRISPR/Cas9 toolbox for multiplexed plant genome editing and transcriptional regulation. Plant Physiol. 2015, 169, 971–985. [Google Scholar] [CrossRef]
- Tuteja, N.; Mahajan, S. Calcium signaling network in plants: An overview. Plant Signal. Behav. 2007, 2, 79–85. [Google Scholar] [CrossRef]
- Shelake, R.M.; Kadam, U.S.; Kumar, R.; Pramanik, D.; Singh, A.K.; Kim, J.Y. Engineering drought and salinity tolerance traits in crops through CRISPR-mediated genome editing: Targets, tools, challenges, and perspectives. Plant Commun. 2022, 3, 100417. [Google Scholar] [CrossRef]
- Sabzehzari, M.; Zeinali, M.; Naghavi, M.R. CRISPR-based metabolic editing: Next-generation metabolic engineering in plants. Gene 2020, 759, 144993. [Google Scholar] [CrossRef] [PubMed]
- Choudry, M.W.; Riaz, R.; Nawaz, P.; Ashraf, M.; Ijaz, B.; Bakhsh, A. CRISPR-Cas9 mediated understanding of plants’ abiotic stress-responsive genes to combat changing climatic patterns. Funct. Integr. Genom. 2024, 24, 132. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Navazio, L.; Szabo, I. The contribution of organelles to plant intracellular calcium signalling. J. Exp. Bot. 2018, 69, 4175–4193. [Google Scholar] [CrossRef]
- Ghosh, S.; Dahiya, M.; Kumar, A.; Bheri, M.; Pandey, G.K. Calcium imaging: A technique to monitor calcium dynamics in biological systems. Physiol. Mol. Biol. Plants 2023, 29, 1777–1811. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.; Long, Y.; Schmitz-Thom, I.; Wang, X.P.; Zhang, C.; Li, H.; Steinhorst, L.; Manishankar, P.; Ren, X.L.; Offenborn, J.N.; et al. Two spatially and temporally distinct Ca2+ signals convey Arabidopsis thaliana responses to K+ deficiency. New Phytol. 2017, 213, 739–750. [Google Scholar] [CrossRef]
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kabir, M.Y.; Díaz-Pérez, J.C. Calcium Route in the Plant and Blossom-End Rot Incidence. Horticulturae 2025, 11, 807. https://doi.org/10.3390/horticulturae11070807
Kabir MY, Díaz-Pérez JC. Calcium Route in the Plant and Blossom-End Rot Incidence. Horticulturae. 2025; 11(7):807. https://doi.org/10.3390/horticulturae11070807
Chicago/Turabian StyleKabir, Md. Yamin, and Juan Carlos Díaz-Pérez. 2025. "Calcium Route in the Plant and Blossom-End Rot Incidence" Horticulturae 11, no. 7: 807. https://doi.org/10.3390/horticulturae11070807
APA StyleKabir, M. Y., & Díaz-Pérez, J. C. (2025). Calcium Route in the Plant and Blossom-End Rot Incidence. Horticulturae, 11(7), 807. https://doi.org/10.3390/horticulturae11070807