
Promoting Effects of Piriformospora indica on Plant Growth and Development of Tissue-Cultured Cerasus humilis Seedlings
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Fungal Preparation
2.2. Explant Treatment
2.3. Fungus Preparation
2.4. In-Vitro Inoculation
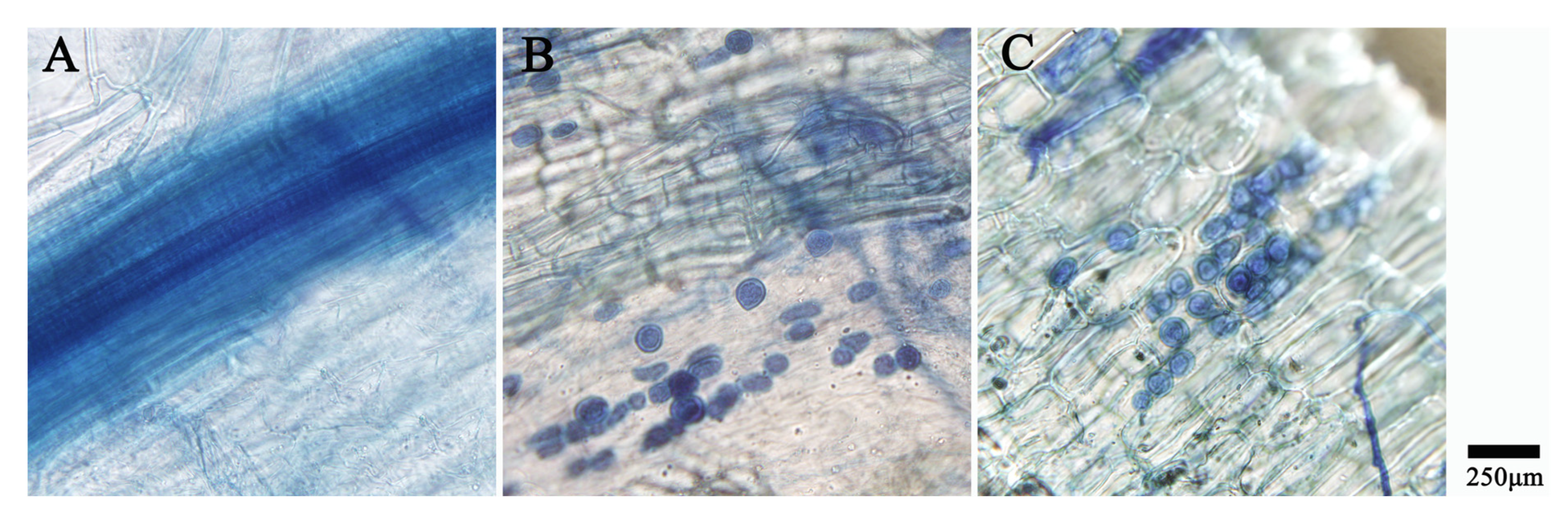
2.5. Piriformospora indica Colonization Detection
2.6. Determination of Plant Growth Parameters
2.7. Determination of Photosynthetic Pigments in Tissue-Cultured Cerasus humilis Seedling Leaves
2.8. Determination of Root Activity, POD Activity, and Hormone Content
2.9. Statistics Analysis
3. Results
3.1. Piriformospora indica Colonization Detection Results
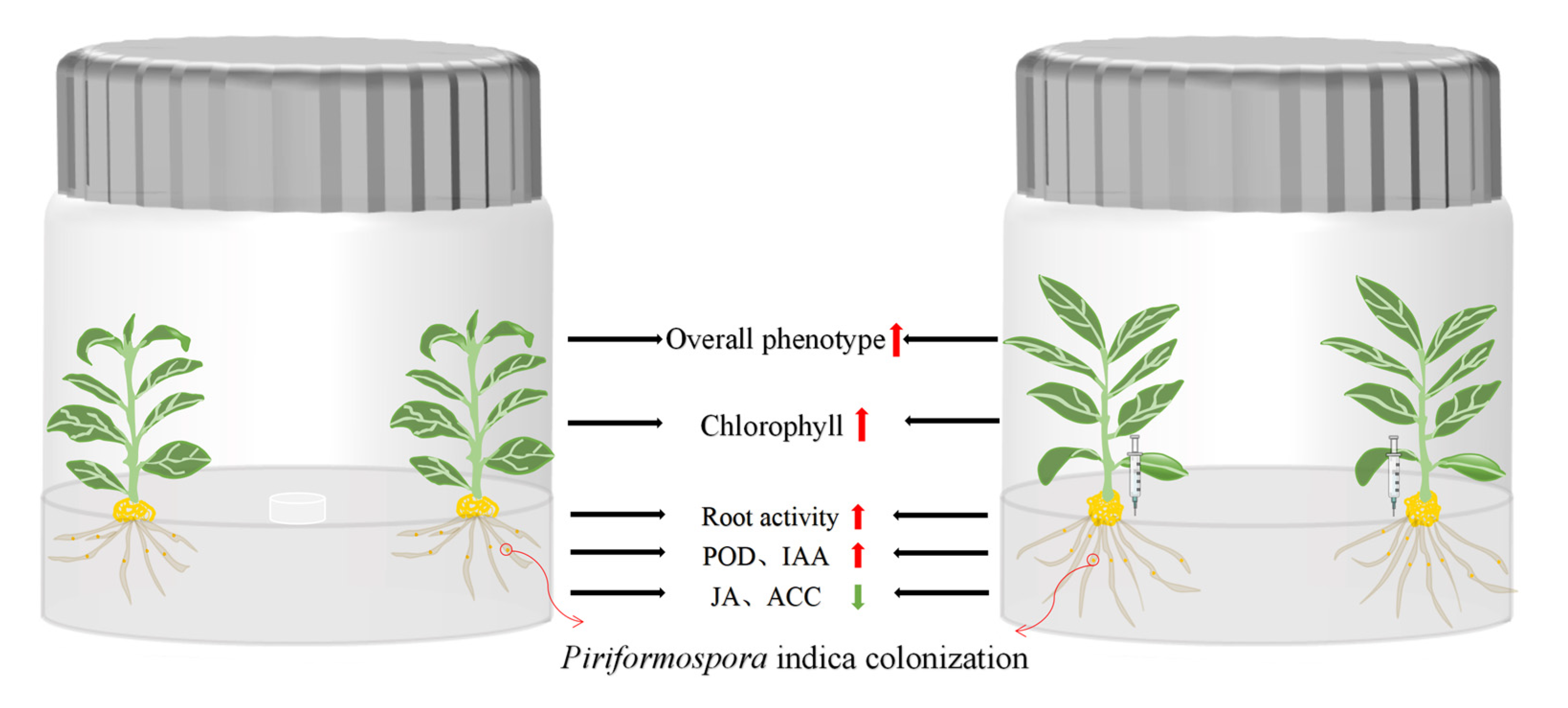
3.2. Effect of Piriformospora indica Colonization on Plant Phenotype of Tissue-Cultured Cerasus humilis Seedlings
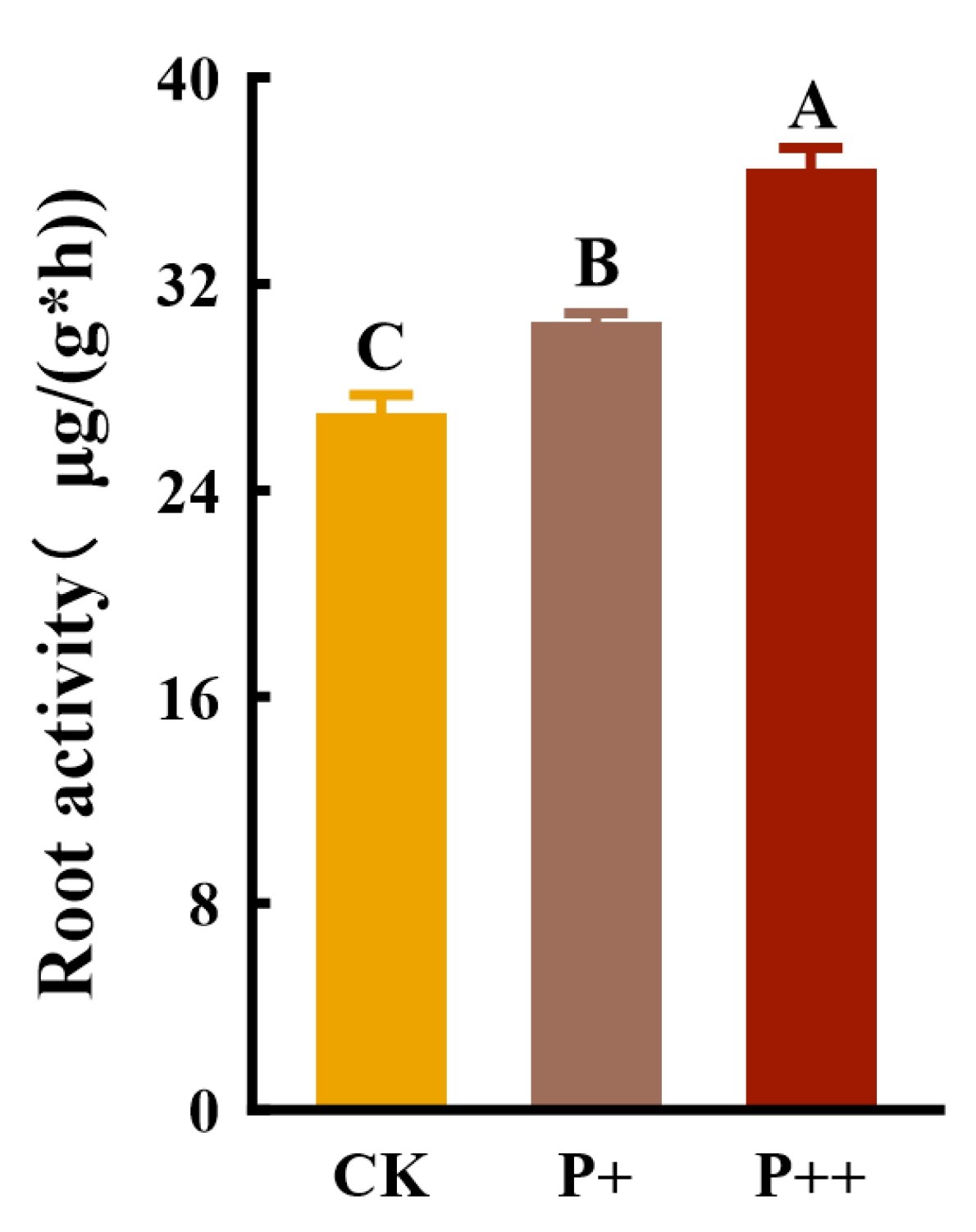
3.3. Effects of Piriformospora indica Colonization on Root Activity of Tissue-Cultured Cerasus humilis Seedlings
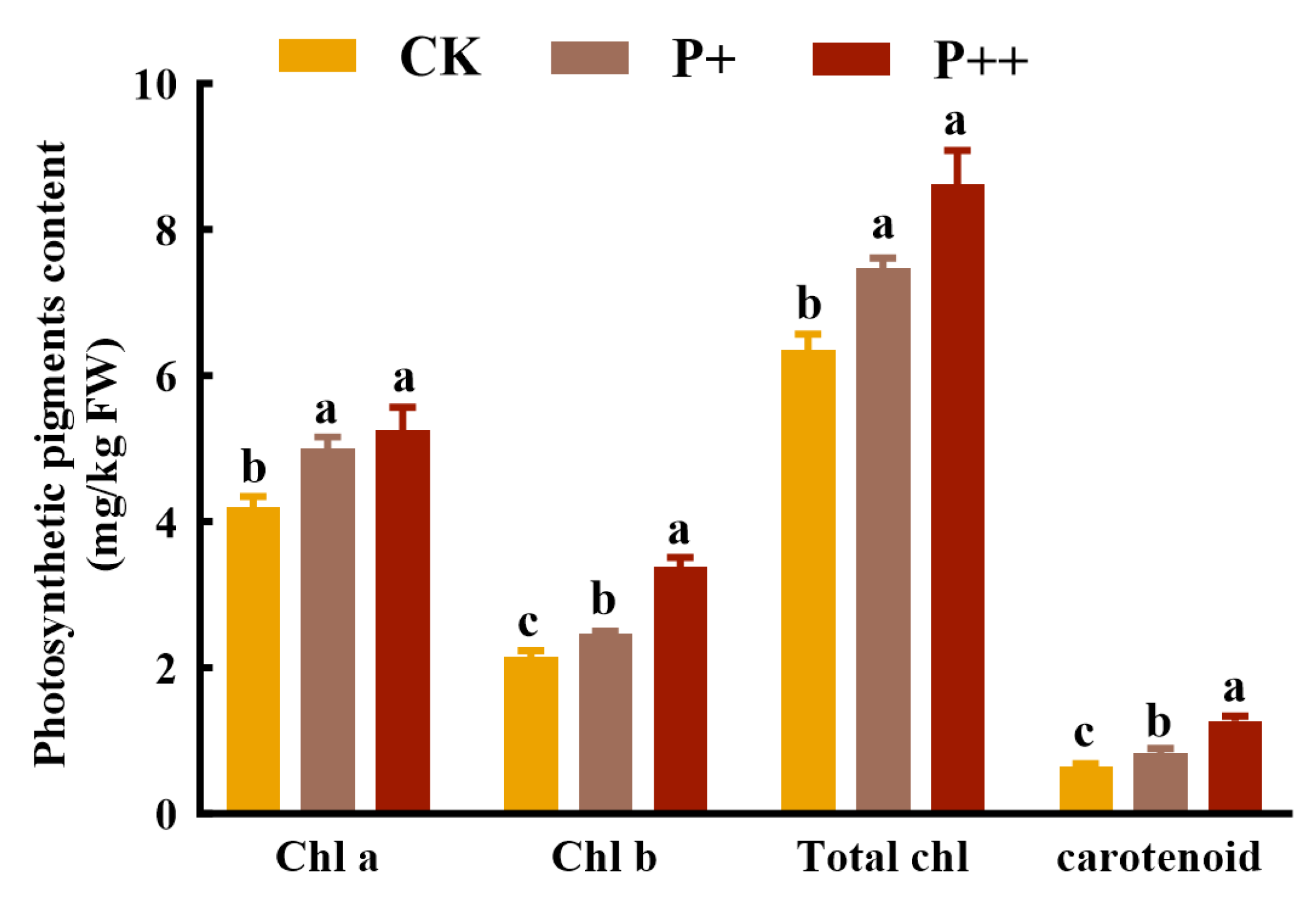
3.4. Effects of Piriformospora indica Colonization on Photosynthetic Pigment Content in Leaves of Tissue-Cultured Cerasus humilis Seedlings
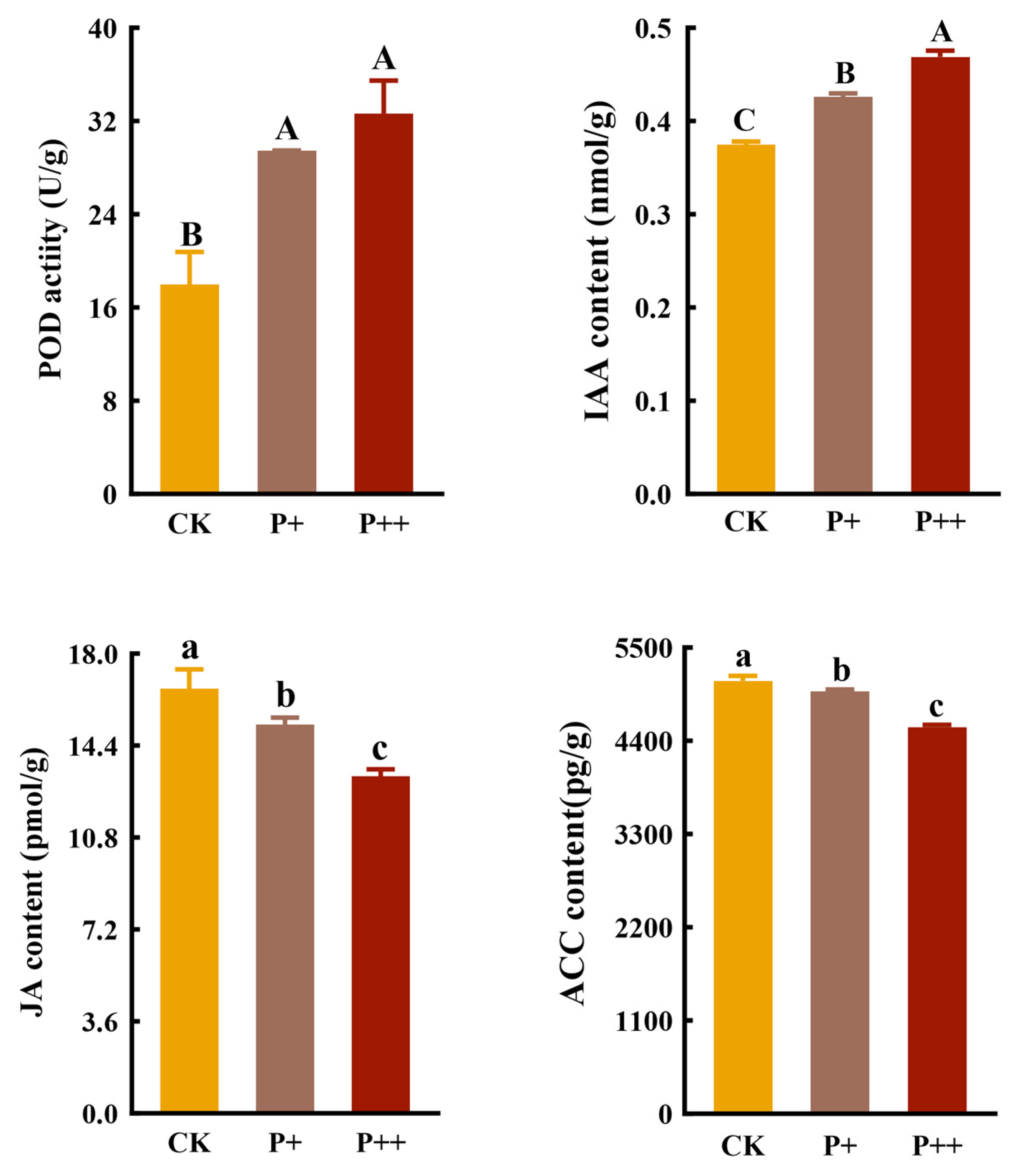
3.5. Effects of Piriformospora indica Colonization on POD Activity and Hormones in Roots of Tissue-Cultured Cerasus humilis Seedlings
4. Discussion
4.1. The Successful Colonization of Piriformospora indica Significantly Promoted the Growth of Tissue-Cultured Cerasus humilis Seedlings
4.2. After the Colonization of Piriformospora indica, the Root and POD Activity Increased, Which Was Beneficial to Root Growth and Development
4.3. The Colonization of Piriformospora indica Greatly Promoted the Accumulation of Photosynthetic Pigments in Tissue-Cultured Cerasus humilis Seedlings
4.4. Piriformospora indica Promoted the Root Growth of Cerasus humilis Seedlings by Inducing IAA Biosynthesis and Inhibiting the Accumulation of JA and ACC
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| P. indica | Piriformospora indica |
| C. humilis | Cerasus humilis Potato dextrose agar |
| PDA | |
| P+ | P. indica colony segments |
| P++ | P. indica spore suspensions |
| POD | Peroxidase |
| IAA | Indole-3-acetic-acid |
| JA | Jasmonic acid |
| ACC | 1-Aminocyclopropane-1-carboxylic acid |
| AMF | Arbuscular mycorrhizal fungi |
References
- Li, H.; Yue, J.; Xia, W.; Li, T.; Huang, X.; Zhang, Y.; Fu, X. Exploring the beneficial effects and mechanisms of Cerasus humilis (Bge.) Sok fruit for calcium supplementation and promotion. Food Biosci. 2023, 54, 102846. [Google Scholar] [CrossRef]
- Liu, M.; Du, J.J. Distribution of germplasm resources and performance of varietal traits in Cerasus humilis. Deciduous Fruit 2010, 42, 18–21. [Google Scholar] [CrossRef]
- Yang, R.; Yang, Y.; Hu, Y.; Yin, L.; Qu, P.; Wang, P.; Mu, X.; Zhang, S.; Xie, P.; Cheng, C.; et al. Comparison of Bioactive Compounds and Antioxidant Activities in Differentially Pigmented Cerasus humilis Fruits. Molecules 2023, 28, 6272. [Google Scholar] [CrossRef]
- Li, W.; Li, O.; Zhang, A.; Li, L.; Hao, J.; Jin, J.; Yin, S. Genotypic diversity of phenolic compounds and antioxidant capacity of Chinese dwarf cherry (Cerasus humilis (Bge.) Sok.) in China. Sci. Hortic. 2014, 175, 208–213. [Google Scholar] [CrossRef]
- Wu, Y.F.; Li, X.Y.; Guo, J.L.; Li, L.G.; Li, D.F.; Zhang, W.; Shao, Z.S. Research progress and application prospect of Cerasus humilis in adapting to abiotic stress. Spec. Wild Econ. Anim. Plant Res. 2022, 44, 104–110. [Google Scholar] [CrossRef]
- Mahanta, M.; Gantait, S. Trends in plant tissue culture and genetic improvement of gerbera. Hortic. Plant J. 2024, 11, 974–988. [Google Scholar] [CrossRef]
- Cao, Q.; Du, J.J. Study on the factors affecting the survival of transplanted seedlings of Cerasus humilis. J. Shanxi Agric. Univ. (Nat. Sci. Ed.) 2009, 29, 238–242.s. [Google Scholar] [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Knight, C.G.; Nicolitch, O.; Williams, A. Harnessing rhizosphere microbiomes for drought-resilient crop production. Science 2020, 368, 270–274. [Google Scholar] [CrossRef]
- Basiru, S.; Ait Si Mhand, K.; Elfermi, R.; Khatour, I.; Errafii, K.; Legeay, J.; Hijri, M. Enhancing chickpea growth through arbuscular mycorrhizal fungus inoculation: Facilitating nutrient uptake and shifting potential pathogenic fungal communities. Mycorrhiza 2024, 35, 1. [Google Scholar] [CrossRef]
- Varma, A.; Savita, V.; Sudha; Sahay, N.; Butehorn, B.; Franken, P. Piriformospora indica, a cultivable plant-growth-promoting root endophyte. Appl. Environ. Microbiol. 1999, 65, 2741–2744. [Google Scholar] [CrossRef]
- Li, L.; Feng, Y.; Qi, F.; Hao, R. Research Progress of Piriformospora indica in Improving Plant Growth and Stress Resistance to Plant. J. Fungi 2023, 9, 965. [Google Scholar] [CrossRef]
- Tripathi, S. Development of Piriformospora indica Based Bioinoculant for Some Economically Important Plants. Ph.D. Thesis, Amity University, Noida, India, 2010. Available online: http://hdl.handle.net/10603/24181 (accessed on 25 March 2025).
- Saleem, S.; Sekara, A.; Pokluda, R. Serendipita indica-A Review from Agricultural Point of View. Plants 2022, 11, 3417. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Mensah, R.A.; Liu, F.; Tian, N.; Qi, Q.; Yeh, K.; Xuhan, X.; Cheng, C.; Lai, Z. Effects of Piriformospora indica on rooting and growth of tissue-cultured banana (Musa acuminata cv. Tianbaojiao) seedlings. Sci. Hortic. 2019, 257, 108649. [Google Scholar] [CrossRef]
- Baishya, D.; Deka, P.; Kalita, M.C. In vitro co-cultivation of Piriformospora indica filtrate for improve biomass productivity in Artemisia annua (L.). Symbiosis 2015, 66, 37–46. [Google Scholar] [CrossRef]
- Franken, P. The plant strengthening root endophyte Piriformospora indica: Potential application and the biology behind. Appl. Microbiol. Biotechnol. 2012, 96, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, B.; Hao, X.; Zhang, Y.; Wang, T.; Lu, Z.; Lai, Z.; Cheng, C. Piriformospora indica promotes the growth and enhances the root rot disease resistance of gerbera. Sci. Hortic. 2022, 297, 110946. [Google Scholar] [CrossRef]
- Yin, L.; Qu, P.; Wang, D.; Yan, S.; Gong, Q.; Yang, R.; Hu, Y.; Liu, N.; Cheng, C.; Wang, P.; et al. The Influence of Piriformospora indica Colonization on the Root Development and Growth of Cerasus humilis Cuttings. Plants 2024, 13, 1482. [Google Scholar] [CrossRef]
- Salah, S.M.; Yajing, G.; Dongdong, C.; Jie, L.; Aamir, N.; Qijuan, H.; Weimin, H.; Mingyu, N.; Jin, H. Seed priming with polyethylene glycol regulating the physiological and molecular mechanism in rice (Oryza sativa L.) under nano-ZnO stress. Sci. Rep. 2015, 5, 14278. [Google Scholar] [CrossRef]
- Liu, W.; Tan, M.; Qu, P.; Huo, C.; Liang, W.; Li, R.; Jia, Y.; Fan, X.; Cheng, C. Use of Piriformospora indica to promote growth of strawberry daughter plants. Horticulturae 2022, 8, 370. [Google Scholar] [CrossRef]
- Qiang, X.; Weiss, M.; Kogel, K.H.; Schäfer, P. Piriformospora indica-a mutualistic basidiomycete with an exceptionally large plant host range. Mol. Plant Pathol. 2012, 13, 508–518. [Google Scholar] [CrossRef]
- Chien, Y.; Lin, N. Effects of biohardening with Serendipita indica on strawberry growth and resistance to Colletotrichum gloeosporioides. J. Phytomed. 2018, 60, 1–8. [Google Scholar] [CrossRef]
- Liu, H.C.; Li, M.J.; Jin, L.; Tian, D.Q.; Zhu, K.Y.; Zhang, J.Q.; Zhou, J.; Tan, C. Effect of Piriformospora indica on the growth of Oncidium orchid tissue culture seedlings. J. Zhejiang Agric. Sci. 2019, 60, 642–645. [Google Scholar] [CrossRef]
- Dong, S.; Tian, Z.; Chen, P.J.; Senthil Kumar, R.; Shen, C.H.; Cai, D.; Oelmüllar, R.; Yeh, K.W. The maturation zone is an important target of Piriformospora indica in Chinese cabbage roots. J. Exp. Bot. 2013, 64, 4529–4540. [Google Scholar] [CrossRef]
- Ding, J.T. Optimization of Bougainvillea Tissue Culture by Response Surface Methodology and the Effect of Piriformospora indica on Its Growth. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2022. [Google Scholar]
- Moreira, B.; Mendes, F.; Mendes, I.; Paula, T.; Junior, P.P.; Salomão, L.; Stürmer, S.; Otoni, W.; Kasuya, M. The interaction between arbuscular mycorrhizal fungi and Piriformospora indica improves the growth and nutrient uptake in micropropagation-derived pineapple plantlets. Sci. Hortic. 2015, 197, 183–192. [Google Scholar] [CrossRef]
- Li, J.; He, P.; Jin, Q.; Chen, J.; Chen, D.; Dai, X.; Ding, S.; Chu, L. Aeration Alleviated the Adverse Effects of Nitrogen Topdressing Reduction on Tomato Root Vigor, Photosynthetic Performance, and Fruit Development. Plants 2024, 13, 1378. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yuan, W.; Di, B.; Zhang, G.; Zhu, J.; Zhou, P.; Ding, T.; Qian, J. Relationship among electrical signals, chlorophyll fluorescence, and root vitality of strawberry seedlings under drought stress. Agronomy 2022, 12, 1428. [Google Scholar] [CrossRef]
- Ben Khaled, L.; Pérez-Gilabert, M.; Dreyer, B.; Oihabi, A.; Honrubia, M.; Morte, A. Peroxidase changes in Phoenix dactylifera palms inoculated with mycorrhizal and biocontrol fungi. Agron. Sustain. Dev. 2008, 28, 411–418. [Google Scholar] [CrossRef]
- Boorboori, M.R.; Zhang, H. The effect of cadmium on soil and plants, and the influence of Serendipita indica (Piriformospora indica) in mitigating cadmium stress. Environ. Geochem. Health 2024, 46, 426. [Google Scholar] [CrossRef]
- Gao, J.; Wang, C.; Tian, P.C.; Liu, C.; Ahsan, T.; Wei, Y.; Huang, Y.Q.; Zhang, S.H. Peanut-Colonized Piriformospora indica Enhanced Drought Tolerance by Modulating the Enzymes and Expression of Drought-Related Genes. J. Basic Microbiol. 2025, 65, e2400305. [Google Scholar] [CrossRef]
- Gaspar, T.; Kevers, C.; Hausman, J.; Berthon, J.; Ripetti, V. Practical uses of peroxidase activity as a predictive marker of rooting performance of micropropagated shoots. Agronomie 1992, 12, 757–765. [Google Scholar] [CrossRef]
- Cheng, C.; Li, D.; Wang, B.; Liao, B.; Qu, P.; Liu, W.; Zhang, Y.; Lü, P. Piriformospora indica colonization promotes the root growth of Dimocarpus longan seedlings. Sci. Hortic. 2022, 301, 111137. [Google Scholar] [CrossRef]
- Scheer, H. Chlorophylls: A Personal Snapshot. Molecules 2022, 27, 1093. [Google Scholar] [CrossRef]
- Sharma, P.; Kharkwal, A.C.; Abdin, M.; Varma, A. Piriformospora indica improves micropropagation, growth and phytochemical content of Aloe vera L. plants. Symbiosis 2014, 64, 11–23. [Google Scholar] [CrossRef]
- Mehri, M.; Ghabooli, M.; Movahedi, Z. Contribution of Serendipita indica on growth improvement, antioxidative capacity of Dracocephalum kotschyi, and its resistance against cadmium stress. Int. Microbiol. 2023, 26, 821–831. [Google Scholar] [CrossRef]
- Xu, L.; Wu, C.; Oelmüller, R.; Zhang, W. Role of Phytohormones in Piriformospora indica-Induced Growth Promotion and Stress Tolerance in Plants: More Questions Than Answers. Front. Microbiol. 2018, 9, 1646. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Johnson, J.M.; Chien, C.T.; Sun, C.; Cai, D.; Lou, B.; Oelmüller, R.; Yeh, K.W. Growth promotion of Chinese cabbage and Arabidopsis by Piriformospora indica is not stimulated by mycelium-synthesized auxin. Mol. Plant-Microbe Interact. 2011, 24, 421–431. [Google Scholar] [CrossRef]
- Shi, Q.Y. Optimizing Tissue Culture System of Stem Explants and the Influence of Piriformospora indica on the Growth and Develepment in Tetrastigma hemsleyanum. Master’s Thesis, Zhejiang University, Hangzhou, China, 2015. [Google Scholar]
- Vahabi, K.; Dorcheh, S.K.; Monajembashi, S.; Westermann, M.; Reichelt, M.; Falkenberg, D.; Hemmerich, P.; Sherameti, I.; Oelmüller, R. Stress promotes Arabidopsis-Piriformospora indica interaction. Plant Signal. Behav. 2016, 11, e1136763. [Google Scholar] [CrossRef]
- Negi, S.; Sukumar, P.; Liu, X.; Cohen, J.D.; Muday, G.K. Genetic dissection of the role of ethylene in regulating auxin-dependent lateral and adventitious root formation in tomato. Plant J. 2010, 61, 3–15. [Google Scholar] [CrossRef]
- Qu, P.; Zhang, Z.; Li, R.; Liu, R.; Zhang, Y.; Cheng, C. Insights into the Rooting and Growth-Promoting Effects of endophytic Fungus Serendipita indica in Blueberry (Vaccinium corymbosum). J. Plant Growth Regul. 2024, 44, 2235–2246. [Google Scholar] [CrossRef]
- Zheng, M.; Zhong, S.; Wang, W.; Tang, Z.; Bu, T.; Li, Q. Serendipita indica promotes the growth of tartary buckwheat by stimulating hormone synthesis, metabolite production, and increasing systemic resistance. J. Fungi 2023, 9, 1114. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 d | 14 d | |||||
|---|---|---|---|---|---|---|
| CK | P+ | P++ | CK | P+ | P++ | |
| plant height (cm) | 0.83 ± 0.10 c | 1.89 ± 0.20 b | 2.26 ± 0.13 a | 1.85 ± 0.07 c | 2.83 ± 0.10 b | 3.29 ± 0.16 a |
| total root length (cm) | 0.40 ± 0.10 b | 0.86 ± 0.12 a | 1.03 ± 0.11 a | 0.74 ± 0.09 c | 1.18 ± 0.09 b | 1.64 ± 0.08 a |
| root number | 2.00 ± 0.00 b | 5.00 ± 1.00 a | 5.00 ± 1.00 a | 3.00 ± 1.00 b | 4.00 ± 1.00 b | 6.00 ± 1.00 a |
| leaf number | 3.00 ± 2.00 c | 8.00 ± 1.00 b | 14.00 ± 1.00 a | 9.00 ± 1.00 c | 14.00 ± 2.00 b | 17.00 ± 1.00 a |
| plant fresh weight (g) | 0.12 ± 0.02 c | 0.23 ± 0.03 b | 0.28 ± 0.02 a | 0.18 ± 0.02 c | 0.27 ± 0.02 b | 0.37 ± 0.02 a |
| plant dry weight (g) | 0.02 ± 0.01 a | 0.06 ± 0.01 a | 0.08 ± 0.06 a | 0.08 ± 0.01 c | 0.14 ± 0.01 b | 0.2 ± 0.02 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, L.; Cheng, J.; Liu, Y.; Guan, Y.; Jia, L.; Zhang, S.; Wang, P.; Mu, X.; Zhang, J. Promoting Effects of Piriformospora indica on Plant Growth and Development of Tissue-Cultured Cerasus humilis Seedlings. Horticulturae 2025, 11, 797. https://doi.org/10.3390/horticulturae11070797
Yin L, Cheng J, Liu Y, Guan Y, Jia L, Zhang S, Wang P, Mu X, Zhang J. Promoting Effects of Piriformospora indica on Plant Growth and Development of Tissue-Cultured Cerasus humilis Seedlings. Horticulturae. 2025; 11(7):797. https://doi.org/10.3390/horticulturae11070797
Chicago/Turabian StyleYin, Lu, JinYang Cheng, YunPeng Liu, YinTao Guan, LuTing Jia, Shuai Zhang, PengFei Wang, XiaoPeng Mu, and JianCheng Zhang. 2025. "Promoting Effects of Piriformospora indica on Plant Growth and Development of Tissue-Cultured Cerasus humilis Seedlings" Horticulturae 11, no. 7: 797. https://doi.org/10.3390/horticulturae11070797
APA StyleYin, L., Cheng, J., Liu, Y., Guan, Y., Jia, L., Zhang, S., Wang, P., Mu, X., & Zhang, J. (2025). Promoting Effects of Piriformospora indica on Plant Growth and Development of Tissue-Cultured Cerasus humilis Seedlings. Horticulturae, 11(7), 797. https://doi.org/10.3390/horticulturae11070797