
Colletotrichum capsici-Induced Disease Development in Postharvest Pepper Associated with Cell Wall Metabolism and Phenylpropanoid Metabolism


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Inoculum Preparation
2.2. Plant Material and Inoculation
2.3. Firmness Measurement
2.4. Microstructure Observation of Pepper Pericarp
2.5. Lignin Content Determination and Lignin Deposition Observation
2.6. Cell Wall Materials Measurement
2.7. Cell Wall-Degrading Enzyme Activity Determination
2.8. Total Phenolic Content and Flavonoid Content Measurement
2.9. Phenylpropanoid Metabolic Pathway-Related Enzymes Determination
2.10. Chitinase and β-1,3-Glucanase Activity Assay
2.11. Statistical Analysis
3. Results
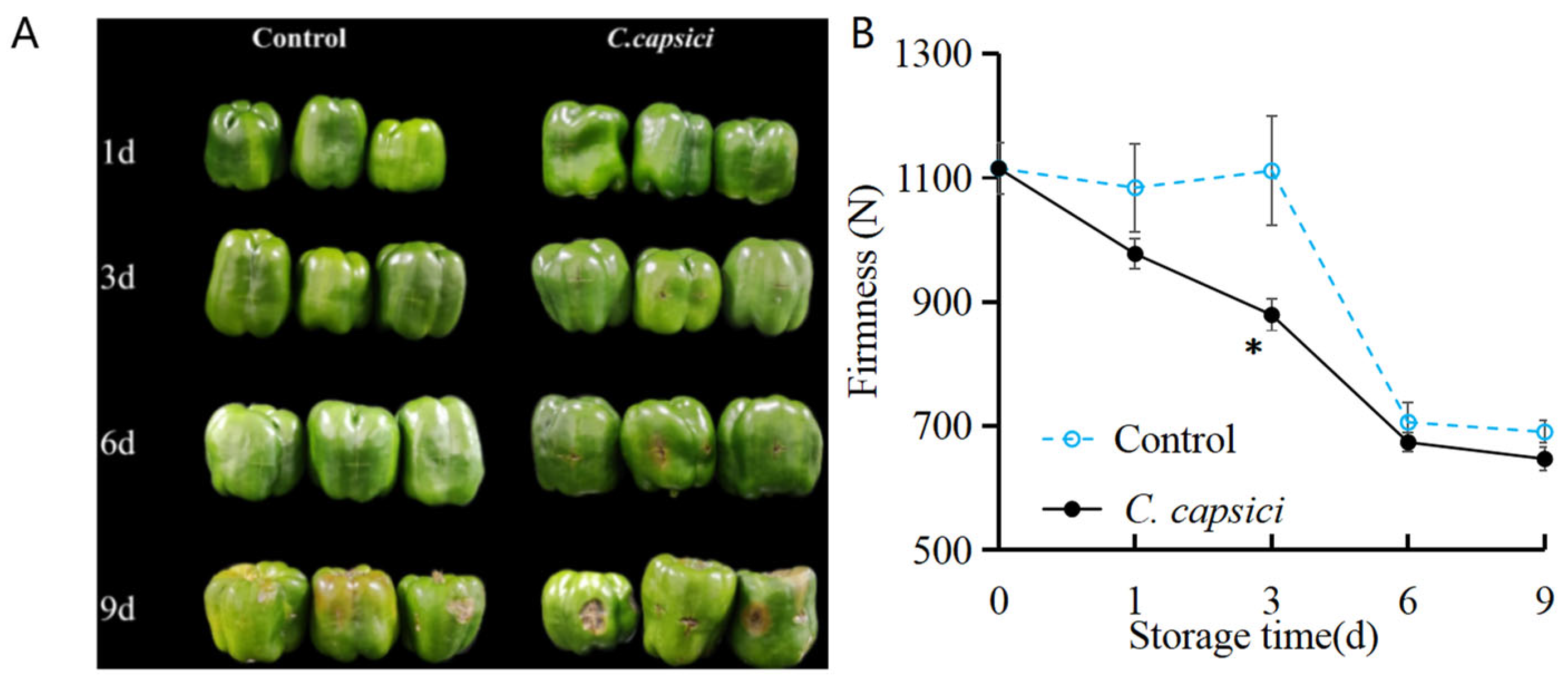
3.1. C. capsici Infestation on Appearance Quality and Firmness of Peppers
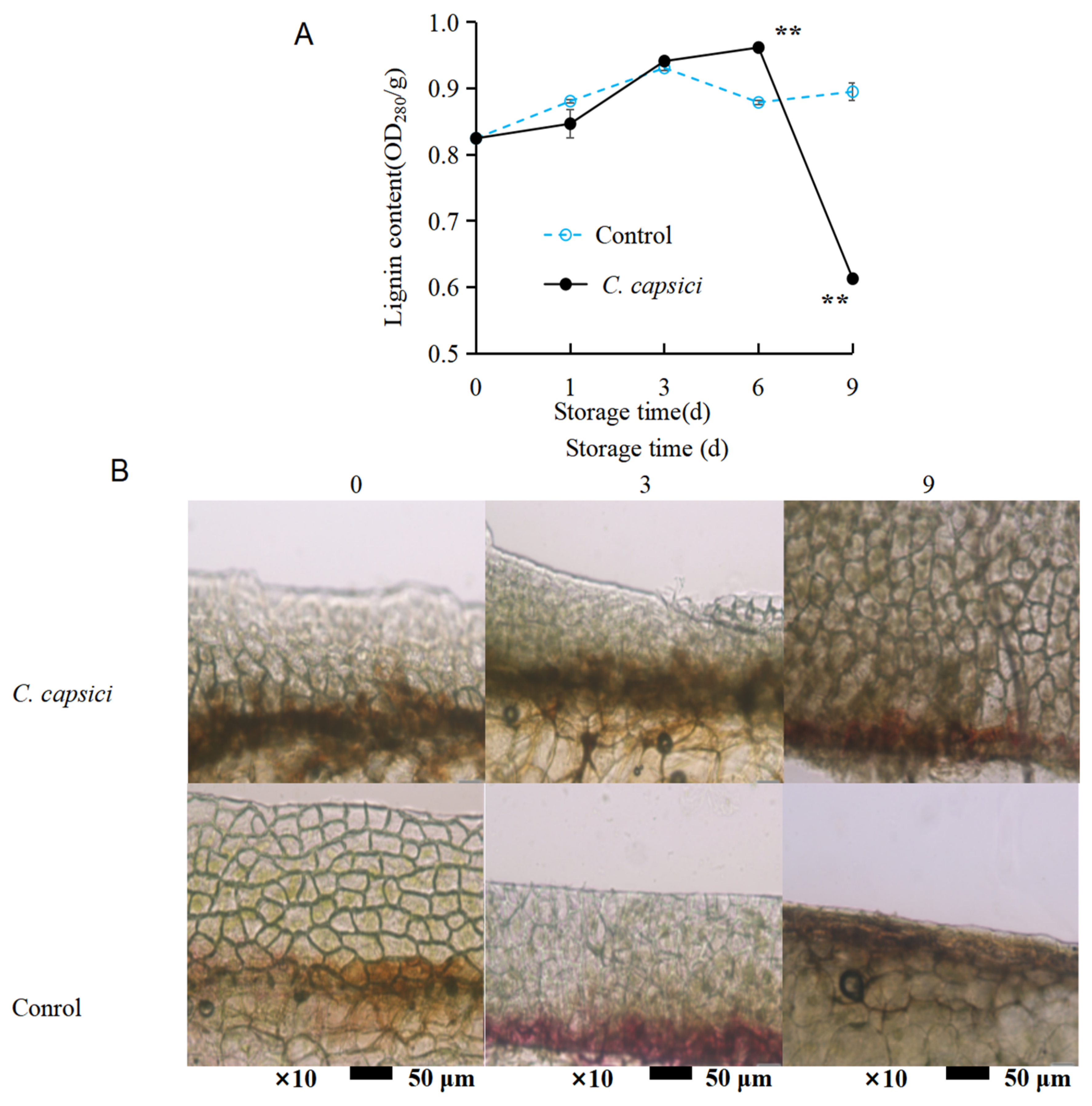
3.2. C. capsici Infestation on Microstructure of Pepper Epidermis
3.3. C. capsici Infestation on Cell-Wall Polysaccharides of Peppers
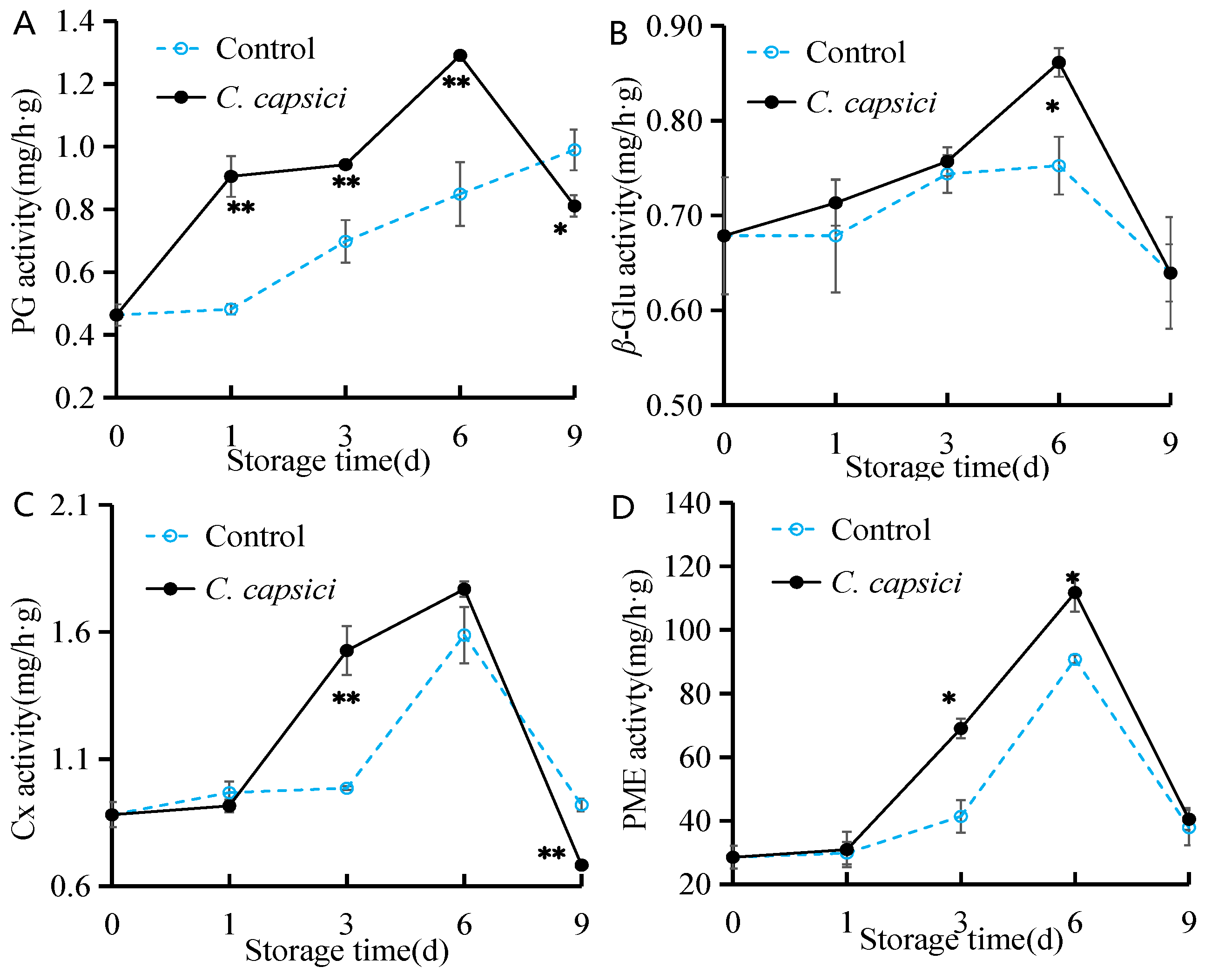
3.4. C. capsici Infestation on Activities of Cell Wall Degradation Enzymes
3.5. C. capsici Infestation on Lignin Content and Lignin Deposition of Peppers
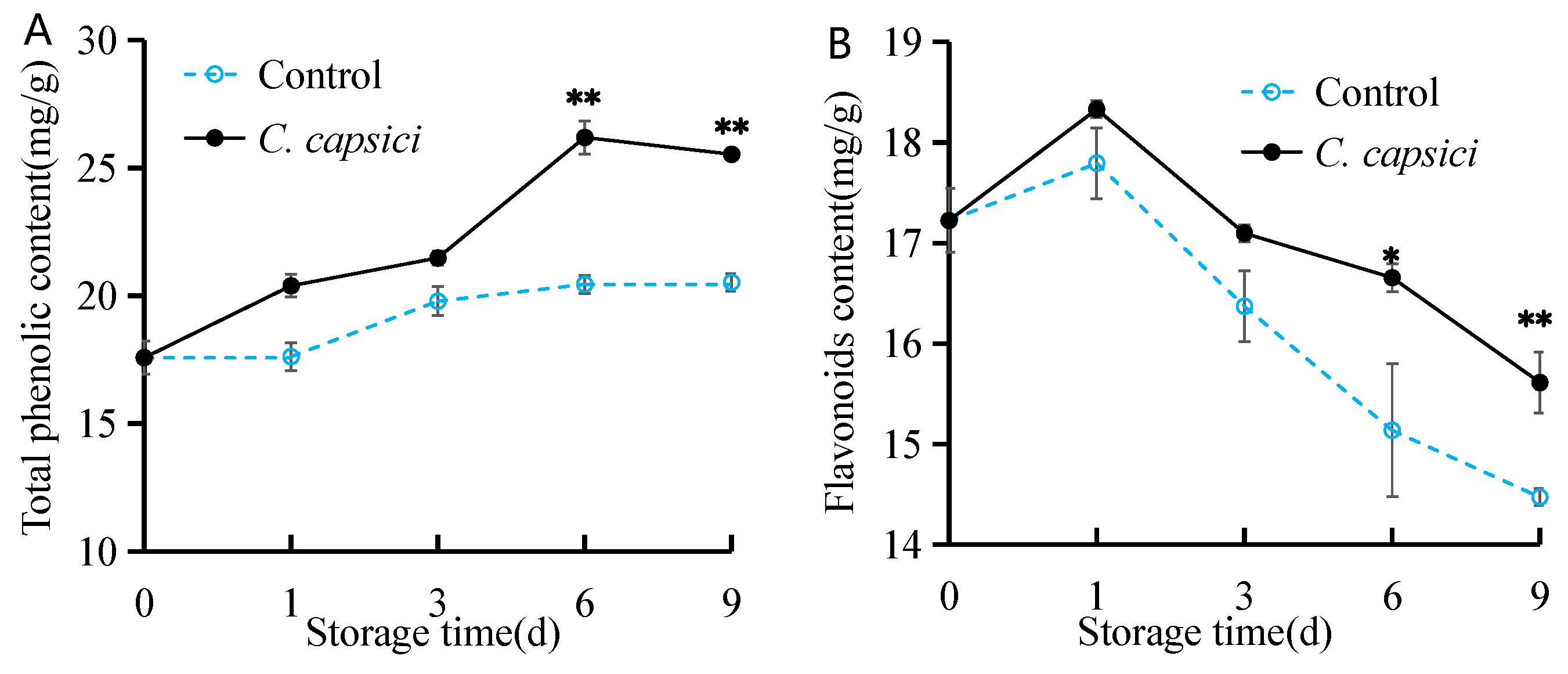
3.6. C. capsici Infestation on Total Phenolic Content and Flavonoid Content of Peppers
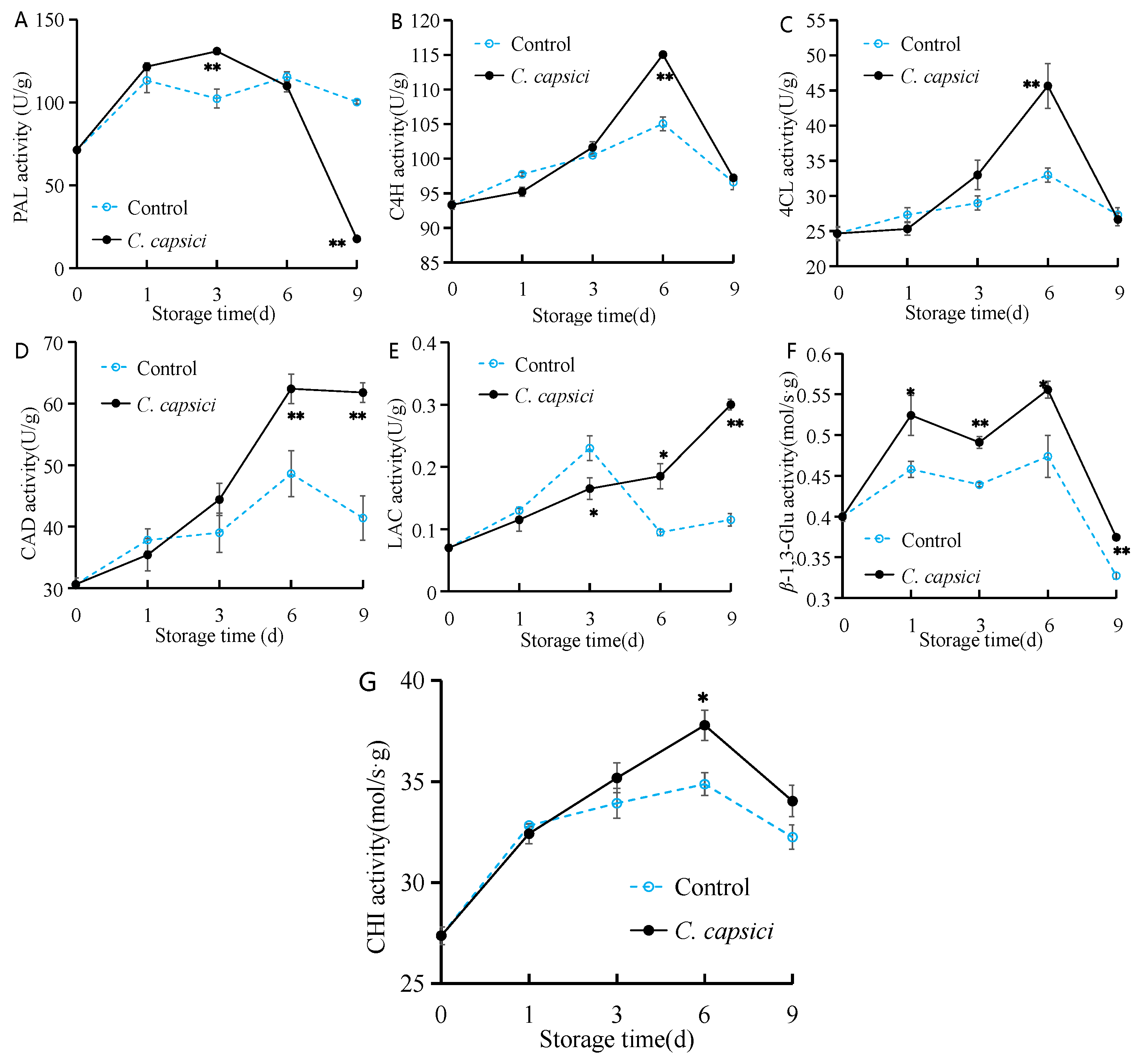
3.7. C. capsici Infestation on Activities of Disease Resistance-Related Enzymes of Peppers
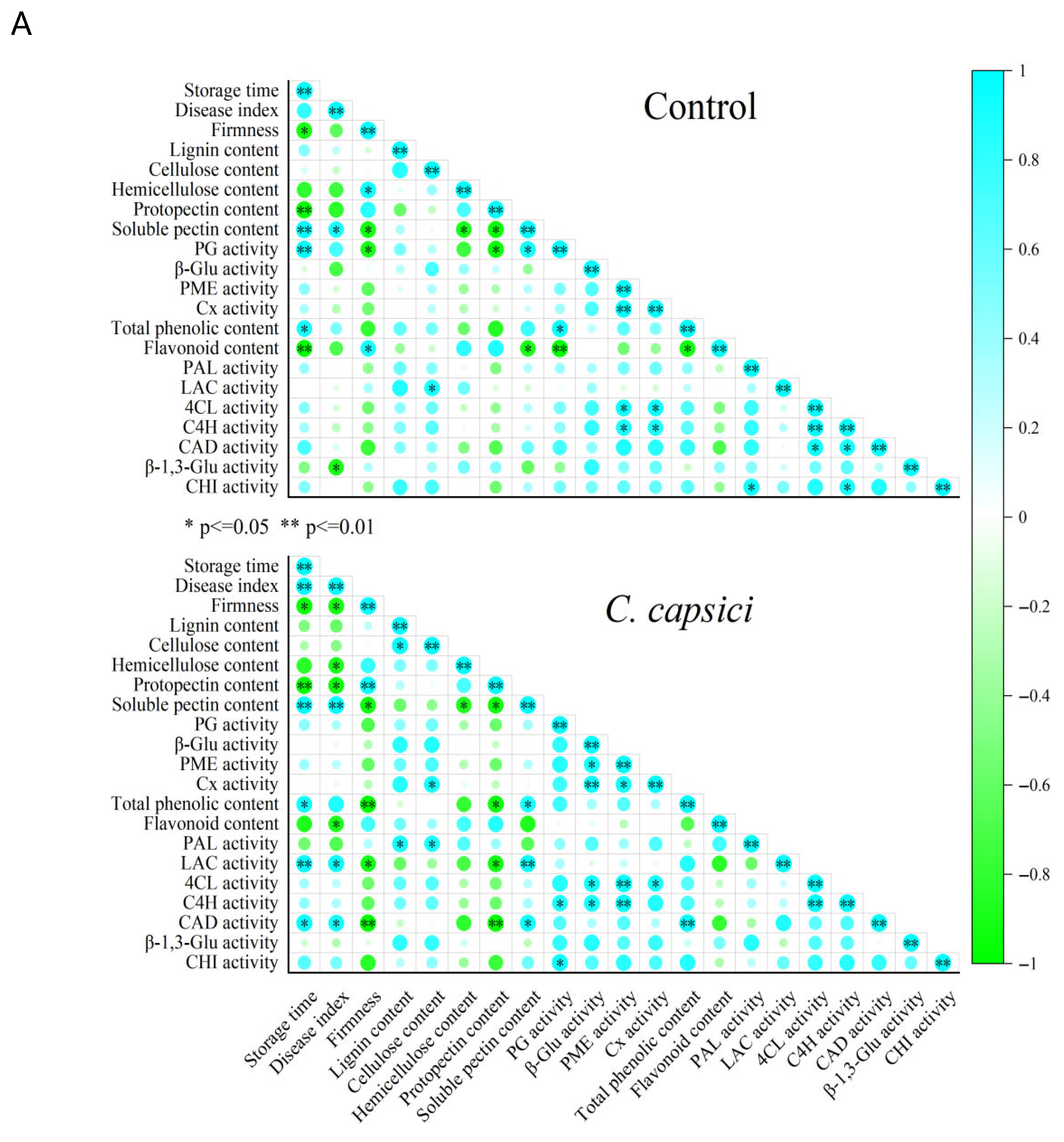
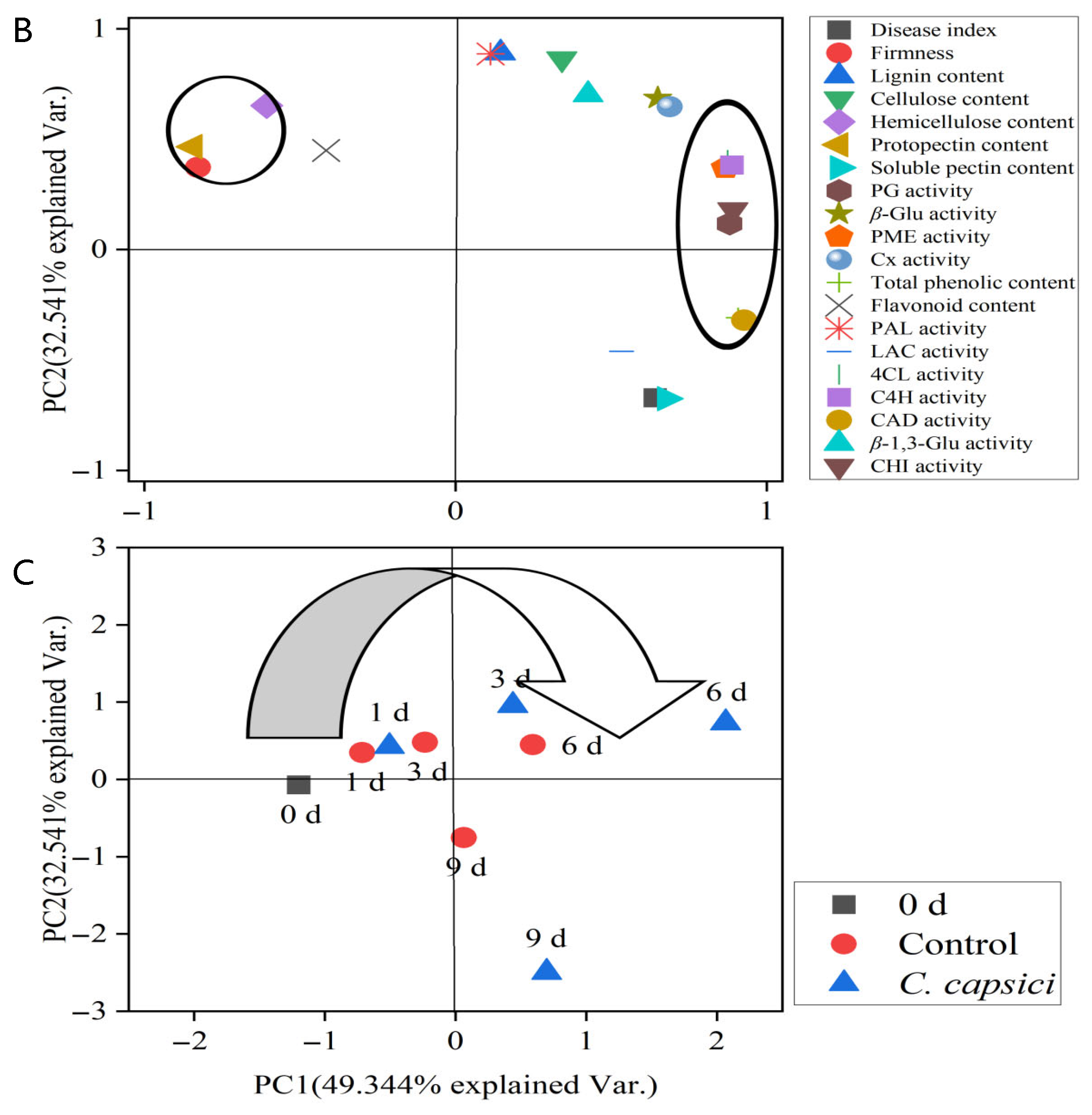
3.8. Correlation and Principal Component Analysis (PCA)
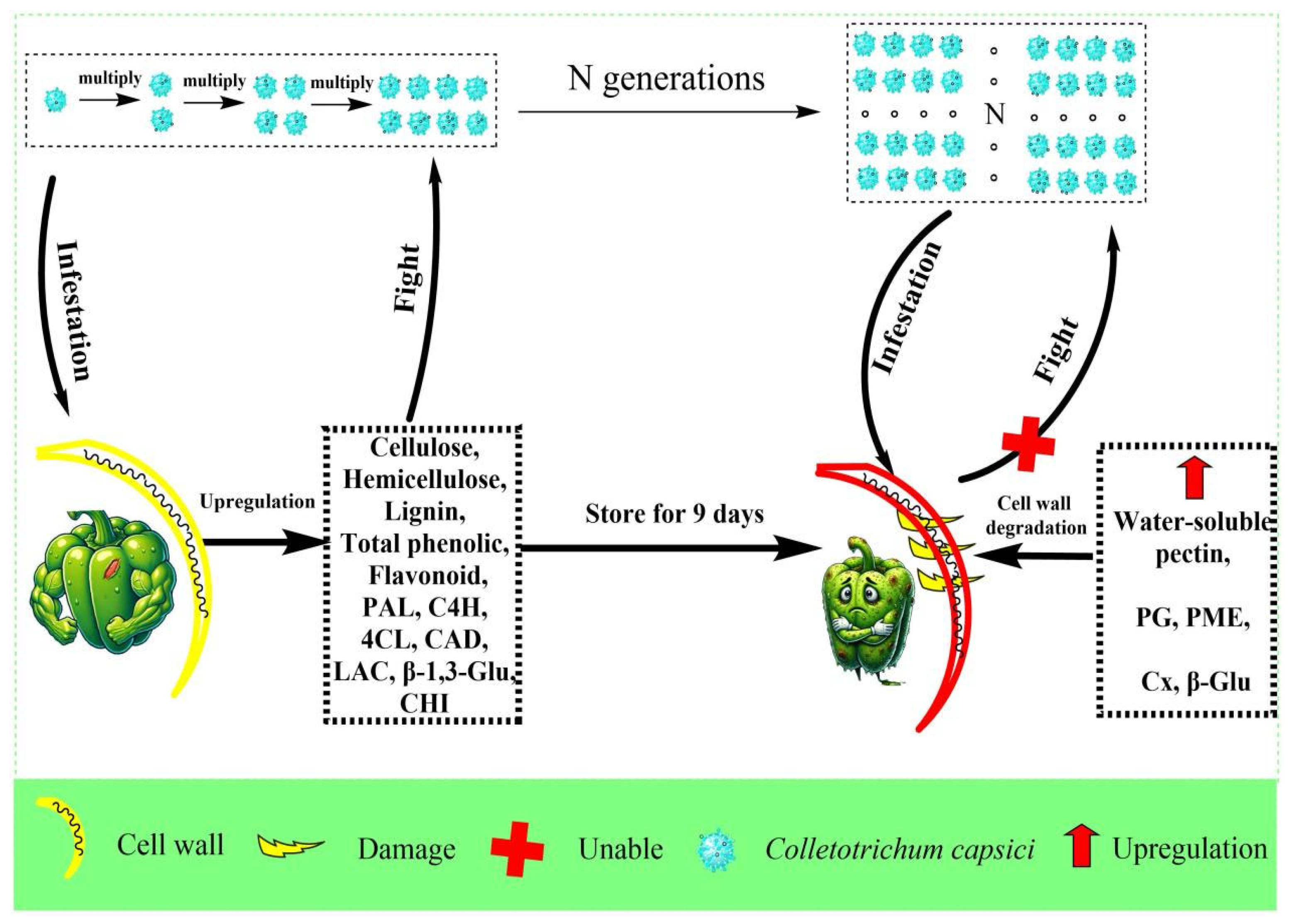
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Queiroz, A.G.D.; Ramos, E.G.; Armond, C.; Felipini, R.B.; Di Piero, R.M. Coating based on gelatin and propolis for the control of anthracnose in postharvest of bell pepper. Food Control 2024, 166, 110689. [Google Scholar] [CrossRef]
- Liu, C.; Wan, H.; Yang, Y.; Ye, Q.; Zhou, G.; Wang, X.; Ahammed, G.J.; Cheng, Y. Post-Harvest LED light irradiation affects firmness, bioactive substances, and amino acid compositions in chili pepper (Capsicum annum L.). Foods 2022, 11, 2712. [Google Scholar] [CrossRef] [PubMed]
- Tiamiyu, Q.O.; Adebayo, S.E.; Ibrahim, N. Recent advances on postharvest technologies of bell pepper: A review. Heliyon 2023, 9, e15302. [Google Scholar] [CrossRef]
- Barik, S.; Ponnam, N.; Reddy, A.C.; Readdy, D.C.L.; Saha, K.; Acharya, G.C.; Reddy, K.M. Breeding peppers for industrial uses: Progress and prospects. Ind. Crops Prod. 2022, 178, 114626. [Google Scholar] [CrossRef]
- González-Saucedo, A.; Barrera-Necha, L.L.; Ventura-Aguilar, R.I.; Correa-Pacheco, Z.N.; Bautista-Baños, S.; Hernández-López, M. Extension of the postharvest quality of bell pepper by applying nanostructured coatings of chitosan with Byrsonima crassifolia extract (L.) Kunth. Postharvest Biol. Technol. 2019, 149, 74–82. [Google Scholar] [CrossRef]
- de SÁ Mendes, N.; Branco De Andrade Gonçalves, É.C. The role of bioactive components found in peppers. Trends Food Sci. Technol. 2020, 99, 229–243. [Google Scholar] [CrossRef]
- Feng, P.; Zhang, X.; Godana, E.A.; Ngolong Ngea, G.L.; Dhanasekaran, S.; Gao, L.; Li, J.; Zhao, L.; Zhang, H. Control of postharvest soft rot of green peppers by Bacillus subtilis through regulating ROS metabolism. Physiol. Mol. Plant Pathol. 2024, 131, 102280. [Google Scholar] [CrossRef]
- Diao, Y.; Zhang, C.; Liu, F.; Wang, W.; Liu, L.; Cai, L.; Liu, X. Colletotrichum species causing anthracnose disease of chili in China. Persoonia 2017, 38, 20–37. [Google Scholar] [CrossRef]
- Ali, A.; Bordoh, P.K.; Singh, A.; Siddiqui, Y.; Droby, S. Post-harvest development of anthracnose in pepper (Capsicum spp): Etiology and management strategies. Crop Prot. 2016, 90, 132–141. [Google Scholar] [CrossRef]
- Boukaew, S.; Chumkaew, K.; Petlamul, W.; Srinuanpan, S.; Nooprom, K.; Zhang, Z. Biocontrol effectiveness of Trichoderma asperelloides SKRU-01 and Trichoderma asperellum NST-009 on postharvest anthracnose in chili pepper. Food Control 2024, 163, 110490. [Google Scholar] [CrossRef]
- Ma, L.; Zheng, Y.; Sang, Z.; Ge, Y.; Bai, C.; Fu, A.; Wang, Q.; Watkins, C.B.; Zuo, J. Multi-omics analysis reveals the mechanism of calcium-reduced quality deterioration in mechanically injured green pepper fruit. Postharvest Biol. Technol. 2023, 204, 112437. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, X.; Liang, J.; Fu, Y.; Wang, J.; Jiang, M.; Pan, L. Cell wall and reactive oxygen metabolism responses of strawberry fruit during storage to low voltage electrostatic field treatment. Postharvest Biol. Technol. 2022, 192, 112017. [Google Scholar] [CrossRef]
- Chen, C.; Nie, Z.; Wan, C.; Gan, Z.; Chen, J. Suppression on postharvest juice sac granulation and cell wall modification by chitosan treatment in harvested pummelo (Citrus grandis L. Osbeck) stored at room temperature. Food Chem. 2021, 336, 127636. [Google Scholar] [CrossRef]
- Li, T.; Shi, D.; Wu, Q.; Yin, C.; Li, F.; Shan, Y.; Duan, X.; Jiang, Y. Mechanism of cell wall polysaccharides modification in harvested ‘Shatangju’ mandarin (Citrus reticulate Blanco) fruit caused by Penicillium italicum. Biomolecules 2019, 9, 160. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wu, W.; Fang, X.; Chen, H.; Han, Y.; Niu, B.; Gao, H. Zizania latifolia cell wall polysaccharide metabolism and changes of related enzyme activities during postharvest storage. Foods 2022, 11, 392. [Google Scholar] [CrossRef]
- Yan, B.; Zhang, Z.; Zhang, P.; Zhu, X.; Jing, Y.; Wei, J.; Wu, B. Nitric oxide enhances resistance against black spot disease in muskmelon and the possible mechanisms involved. Sci. Hortic. 2019, 256, 108650. [Google Scholar] [CrossRef]
- Chen, C.; Cai, N.; Wan, C.; Huang, Q.; Chen, J. Cell wall modification and lignin biosynthesis involved in disease resistance against Diaporthe citri in harvested pummelo fruit elicited by carvacrol. J. Sci. Food Agric. 2022, 102, 3140–3149. [Google Scholar] [CrossRef]
- Palou, L.; Smilanick, J.L.; Droby, S. Alternatives to conventional fungicides for the control of citrus postharvest green and blue moulds. Stewart Postharvest Rev. 2008, 2, 2. [Google Scholar] [CrossRef]
- Li, N.; Chen, W.; Wang, B.; Zhang, C.; Wang, Y.; Li, R.; Yan, Y.; He, J. Arbuscular mycorrhizal fungi improve the disease resistance of Lycium barbarum to root rot by activating phenylpropane metabolism. Front. Plant Sci. 2024, 15, 1459651. [Google Scholar] [CrossRef]
- Li, C.; Wang, M.; Guo, Y.; Zhang, S.; Xu, H.; Ge, Y. Activation of the calcium signaling, mitogen-activated protein kinase cascade and phenylpropane metabolism contributes to the induction of disease resistance in pear fruit upon phenylalanine treatment. Postharvest Biol. Technol. 2024, 210, 112782. [Google Scholar] [CrossRef]
- Guo, Y.; Li, C.; Wang, M.; Xu, H.; Zhang, S.; Liu, J.; Jin, Y.; Ge, Y. Postharvest caffeic acid dipping enhances disease resistance and storage capacity of ‘Zaosu’ pear fruit via regulating phenylpropane metabolism. Postharvest Biol. Technol. 2024, 209, 112716. [Google Scholar] [CrossRef]
- Liu, Y.; Lei, X.; Deng, B.; Chen, O.; Deng, L.; Zeng, K. Methionine enhances disease resistance of jujube fruit against postharvest black spot rot by activating lignin biosynthesis. Postharvest Biol. Technol. 2022, 190, 111935. [Google Scholar] [CrossRef]
- Lu, D.; Ren, Y.; Yan, T.; Jia, X.; Xu, H.; Yang, B.; Zhang, X.; He, J. Melatonin improves the postharvest anthracnose resistance of mango fruit by regulating antioxidant activity, the phenylpropane pathway and cell wall metabolism. Eur. J. Plant Pathol. 2025, 171, 17–36. [Google Scholar] [CrossRef]
- Wu, Y.; Fan, W.; Li, X.; Chen, H.; Takac, T.; Samajova, O.; Fabrice, M.R.; Xie, L.; Ma, J.; Samaj, J.; et al. Expression and distribution of extensins and AGPs in susceptible and resistant banana cultivars in response to wounding and Fusarium oxysporum. Sci. Rep. 2017, 7, 42400. [Google Scholar] [CrossRef]
- Jia, S.; Zhang, N.; Dong, C.; Zheng, P.; Ji, H.; Yu, J.; Yan, S.; Chen, C.; Liang, L. Effect of cold plasma treatment on the softening of winter jujubes (Ziziphus jujuba Mill. cv. Dongzao). Horticulturae 2023, 9, 986. [Google Scholar] [CrossRef]
- Lu, M.; Wen, T.; Guo, M.; Li, Q.; Peng, X.; Zhang, Y.; Lu, Z.; Wang, J.; Xu, Y.; Zhang, C. Regulation of intracellular reactive oxygen species levels after the development of Phallus rubrovolvatus rot disease due to Trichoderma koningii Mycoparasitism. J. Fungi 2023, 9, 525. [Google Scholar] [CrossRef]
- Qu, L.; Xu, Z.; Huang, W.; Han, D.; Dang, B.; Ma, X.; Liu, Y.; Xu, J.; Jia, W. Selenium-molybdenum interactions reduce chromium toxicity in Nicotiana tabacum L. by promoting chromium chelation on the cell wall. J. Hazard. Mater. 2024, 461, 132641. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhu, N.; Zhu, C.; Wu, D.; Chen, K. Morphology and cell wall composition changes in lignified cells from loquat fruit during postharvest storage. Postharvest Biol. Technol. 2019, 157, 110975. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, X.; Wu, C.; Fan, G.; Li, T.; Dong, C. Retardation of postharvest softening of blueberry fruit by methyl jasmonate is correlated with altered cell wall modification and energy metabolism. Sci. Hortic. 2021, 276, 109752. [Google Scholar] [CrossRef]
- Song, X.; Dai, H.; Wang, S.; Ji, S.; Zhou, X.; Li, J.; Zhou, Q. Putrescine treatment delayed the softening of postharvest blueberry fruit by inhibiting the expression of cell wall metabolism key gene VcPG1. Plants 2022, 11, 1356. [Google Scholar] [CrossRef]
- Yang, W.; Liu, Y.; Sang, Y.; Ma, Y.; Guo, M.; Bai, G.; Cheng, S.; Chen, G. Influences of ice-temperature storage on cell wall metabolism and reactive oxygen metabolism in Xinjiang (Diaogan) apricot. Postharvest Biol. Technol. 2021, 180, 111614. [Google Scholar] [CrossRef]
- Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Bi, Y.; Jiang, Q.; Mao, R.; Liu, Z.; Huang, Y.; Zhang, M.; Prusky, D.B. Induction of defense response against Alternaria rot in Zaosu pear fruit by exogenous L-lysine through regulating ROS metabolism and activating defense-related proteins. Postharvest Biol. Technol. 2021, 179, 111567. [Google Scholar] [CrossRef]
- Li, Y.; He, H.; Hou, Y.; Kelimu, A.; Wu, F.; Zhao, Y.; Shi, L.; Zhu, X. Salicylic acid treatment delays apricot (Prunus armeniaca L.) fruit softening by inhibiting ethylene biosynthesis and cell wall degradation. Sci. Hortic. 2022, 300, 111061. [Google Scholar] [CrossRef]
- Chen, Y.; Hung, Y.; Chen, M.; Lin, H. Effects of acidic electrolyzed oxidizing water on retarding cell wall degradation and delaying softening of blueberries during postharvest storage. LWT 2017, 84, 650–657. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Lin, H.; Lu, W.; Wang, H.; Chen, Y.; Lin, Y.; Fan, Z. The role of cell wall polysaccharides disassembly in Lasiodiplodia theobromae-induced disease occurrence and softening of fresh longan fruit. Food Chem. 2021, 351, 129294. [Google Scholar] [CrossRef] [PubMed]
- Gwanpua, S.G.; Van Buggenhot, S.; Verlinden, B.E.; Christiaens, S.; Shpigelman, A.; Vicent, V.; Kermani, Z.J.; Nicolai, B.M.; Hendrickx, M.; Geeraerd, A. Pectin modifications and the role of pectin-degrading enzymes during postharvest softening of Jonagold apples. Food Chem. 2014, 1582, 83–91. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Lin, H.; Sun, J.; Lin, Y.; Wang, H.; Lin, M.; Shi, J. Phomopsis longanae Chi-induced changes in activities of cell wall-degrading enzymes and contents of cell call components in pericarp of harvested longan fruit and its relation to disease development. Front. Microbiol. 2018, 9, 1051. [Google Scholar] [CrossRef]
- Wu, Y.; Duan, X.; Jing, G.; Ouyang, Q.; Tao, N. Cinnamaldehyde inhibits the mycelial growth of Geotrichum citri-aurantii and induces defense responses against sour rot in citrus fruit. Postharvest Biol. Technol. 2017, 129, 23–28. [Google Scholar] [CrossRef]
- Savatin, D.V.; Gramegna, G.; ModestiI, V.; Cervone, F. Wounding in the plant tissue: The defense of a dangerous passage. Front. Plant Sci. 2014, 5, 470. [Google Scholar] [CrossRef]
- Narváez-Barragán, D.A.; Tovar-Herrera, O.E.; Guevara-García, A.; Serrano, M.; Martinez-Anaya, C. Mechanisms of plant cell wall surveillance in response to pathogens, cell wall-derived ligands and the effect of expansins to infection resistance or susceptibility. Front. Plant Sci. 2022, 13, 969343. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Deng, L.; Zhou, Y.; Ming, J.; Yao, S.; Zeng, K. Wound healing in citrus fruit is promoted by chitosan and Pichia membranaefaciens as a resistance mechanism against Colletotrichum gloeosporioides. Postharvest Biol. Technol. 2018, 145, 134–143. [Google Scholar] [CrossRef]
- Li, J.; Wu, Z.; Zhu, Z.; Xu, L.; Wu, B.; Li, J. Botrytis cinerea mediated cell wall degradation accelerates spike stalk browning in Munage grape. J. Food Biochem. 2022, 46, e14271. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, H.; Philippe, F.; Domon, J.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Gigli-Bisceglia, N.; Engelsdorf, T.; Hamann, T. Plant cell wall integrity maintenance in model plants and crop species-relevant cell wall components and underlying guiding principles. Cell. Mol. Life Sci. 2020, 77, 2049–2077. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Song, Q.; Yin, F.; Liang, Y.; Song, M.; He, M.; Shuai, L. Colletotrichum capsici-Induced Disease Development in Postharvest Pepper Associated with Cell Wall Metabolism and Phenylpropanoid Metabolism. Horticulturae 2025, 11, 794. https://doi.org/10.3390/horticulturae11070794
Liu Y, Song Q, Yin F, Liang Y, Song M, He M, Shuai L. Colletotrichum capsici-Induced Disease Development in Postharvest Pepper Associated with Cell Wall Metabolism and Phenylpropanoid Metabolism. Horticulturae. 2025; 11(7):794. https://doi.org/10.3390/horticulturae11070794
Chicago/Turabian StyleLiu, Yunfen, Qian Song, Feilong Yin, Yuanli Liang, Mubo Song, Meiying He, and Liang Shuai. 2025. "Colletotrichum capsici-Induced Disease Development in Postharvest Pepper Associated with Cell Wall Metabolism and Phenylpropanoid Metabolism" Horticulturae 11, no. 7: 794. https://doi.org/10.3390/horticulturae11070794
APA StyleLiu, Y., Song, Q., Yin, F., Liang, Y., Song, M., He, M., & Shuai, L. (2025). Colletotrichum capsici-Induced Disease Development in Postharvest Pepper Associated with Cell Wall Metabolism and Phenylpropanoid Metabolism. Horticulturae, 11(7), 794. https://doi.org/10.3390/horticulturae11070794