
Effect of Combined Light Quality and Calcium Chloride Treatments on Growth and Quality of Chinese Kale Sprouts

 , ,
, , 
Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
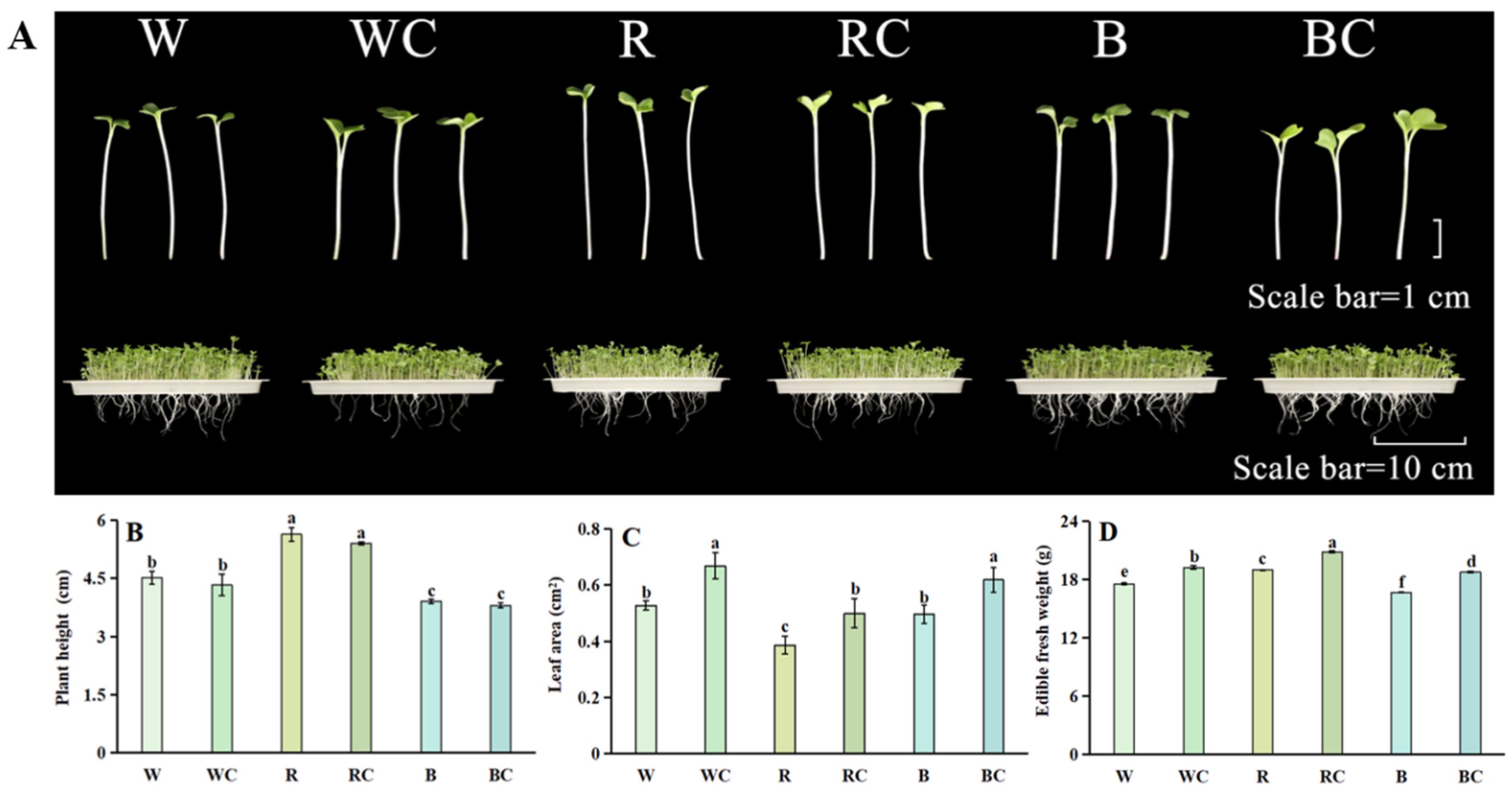
2.2. Growth Parameters
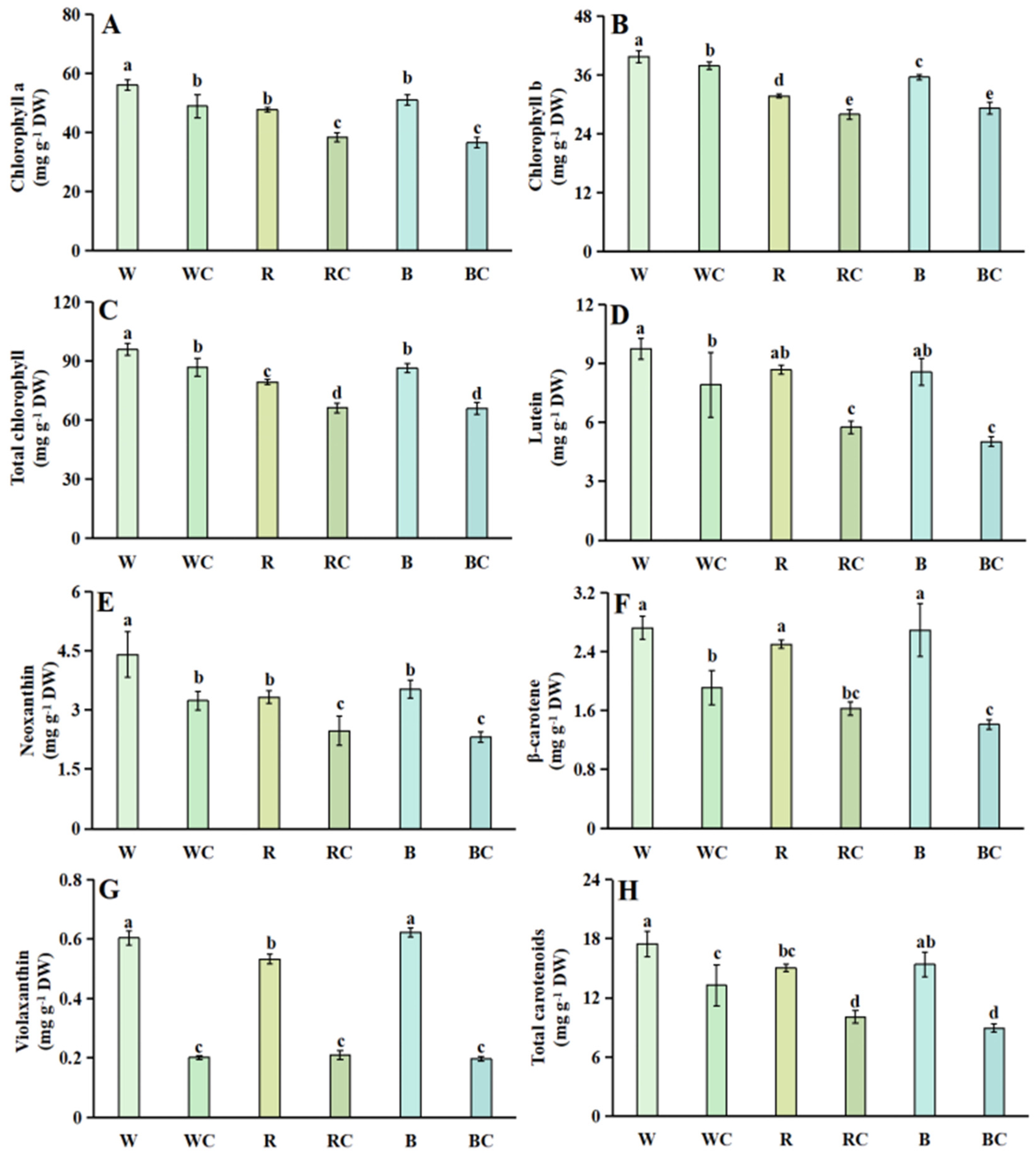
2.3. Chlorophylls and Carotenoids Content
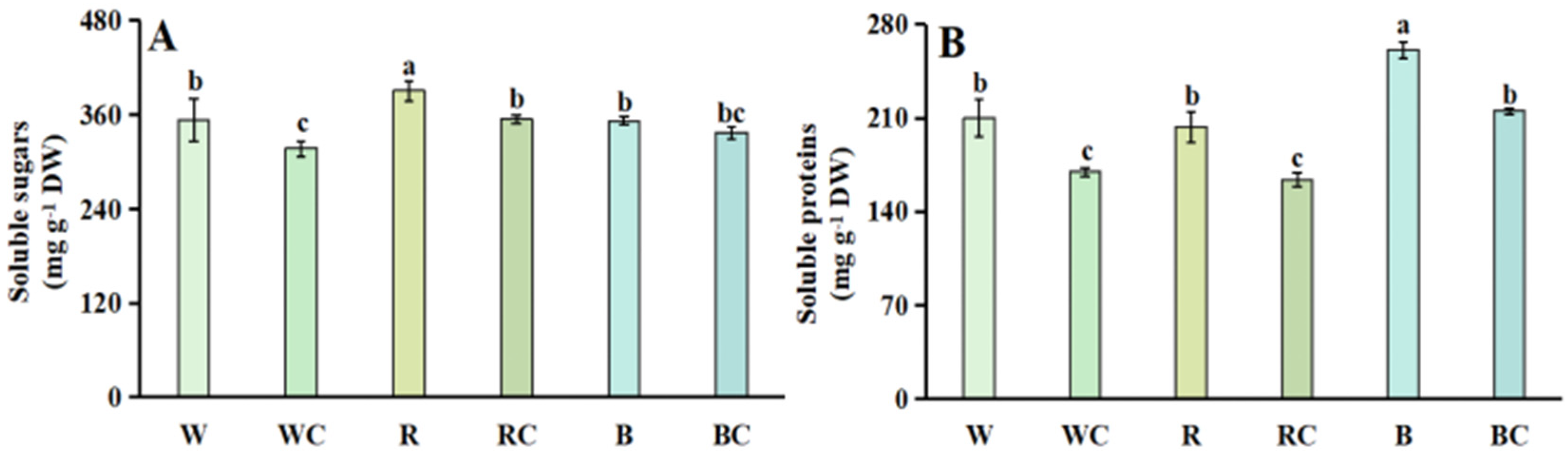
2.4. Soluble Sugar Content
2.5. Soluble Protein Content
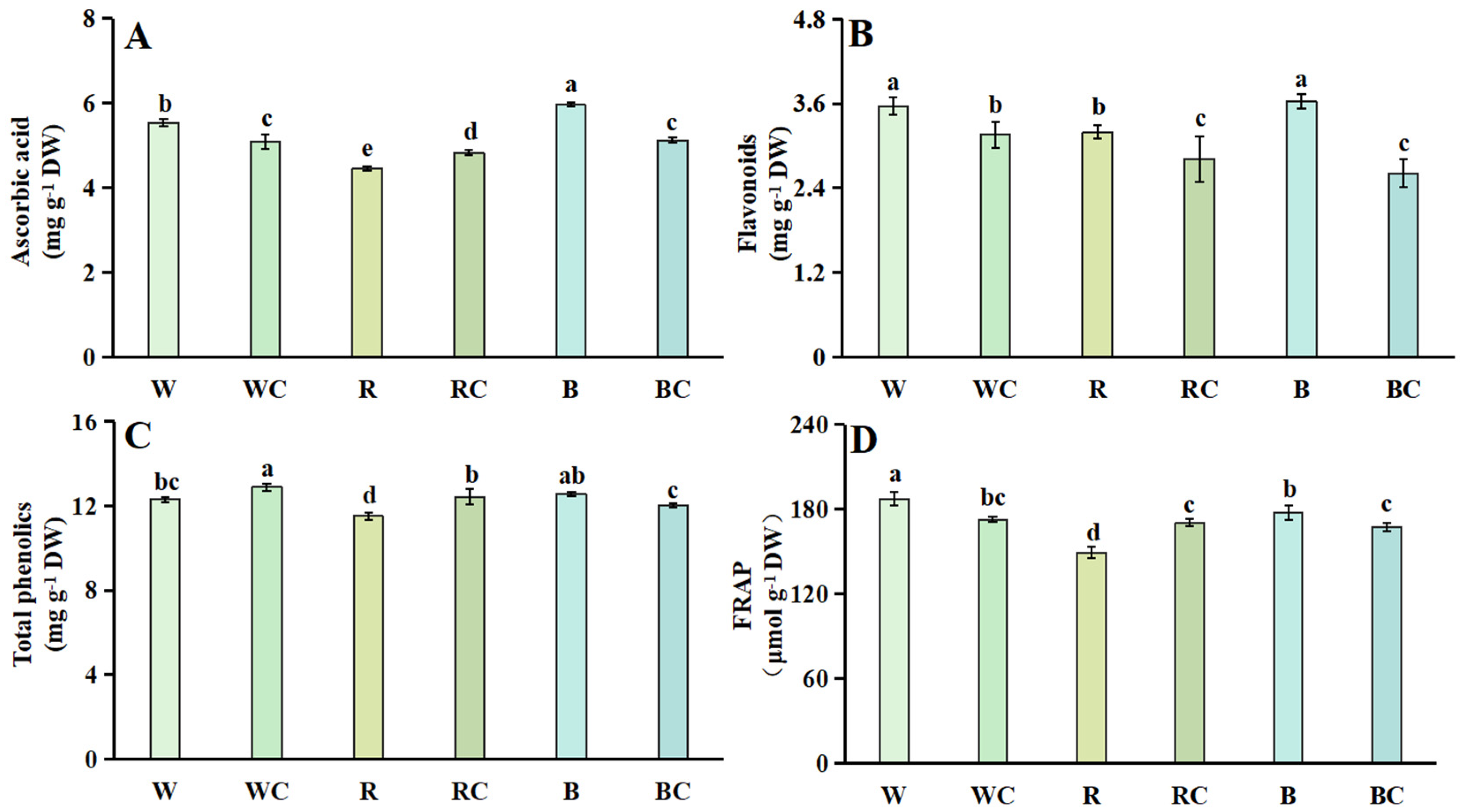
2.6. Ascorbic Acid Content
2.7. Flavonoids Content
2.8. Total Phenolics Content
2.9. Ferric Reducing Antioxidant Power (FRAP)
2.10. Glucosinolates Content
2.11. Statistical Analysis
3. Results
3.1. Sprout Growth
3.2. Chlorophylls and Carotenoids
3.3. Soluble Sugars and Soluble Proteins
3.4. Antioxidant Content and Antioxidant Capacity
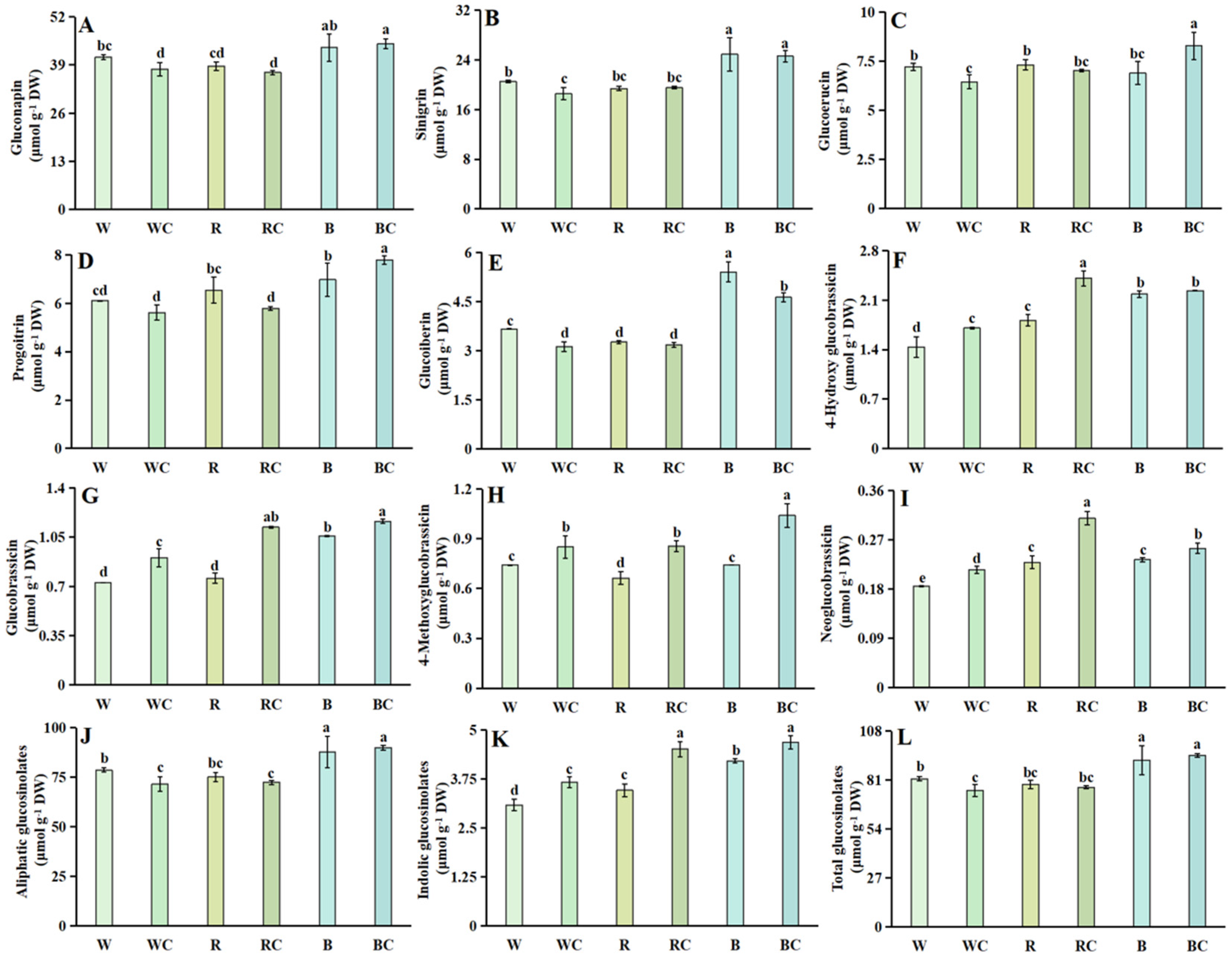
3.5. Glucosinolates
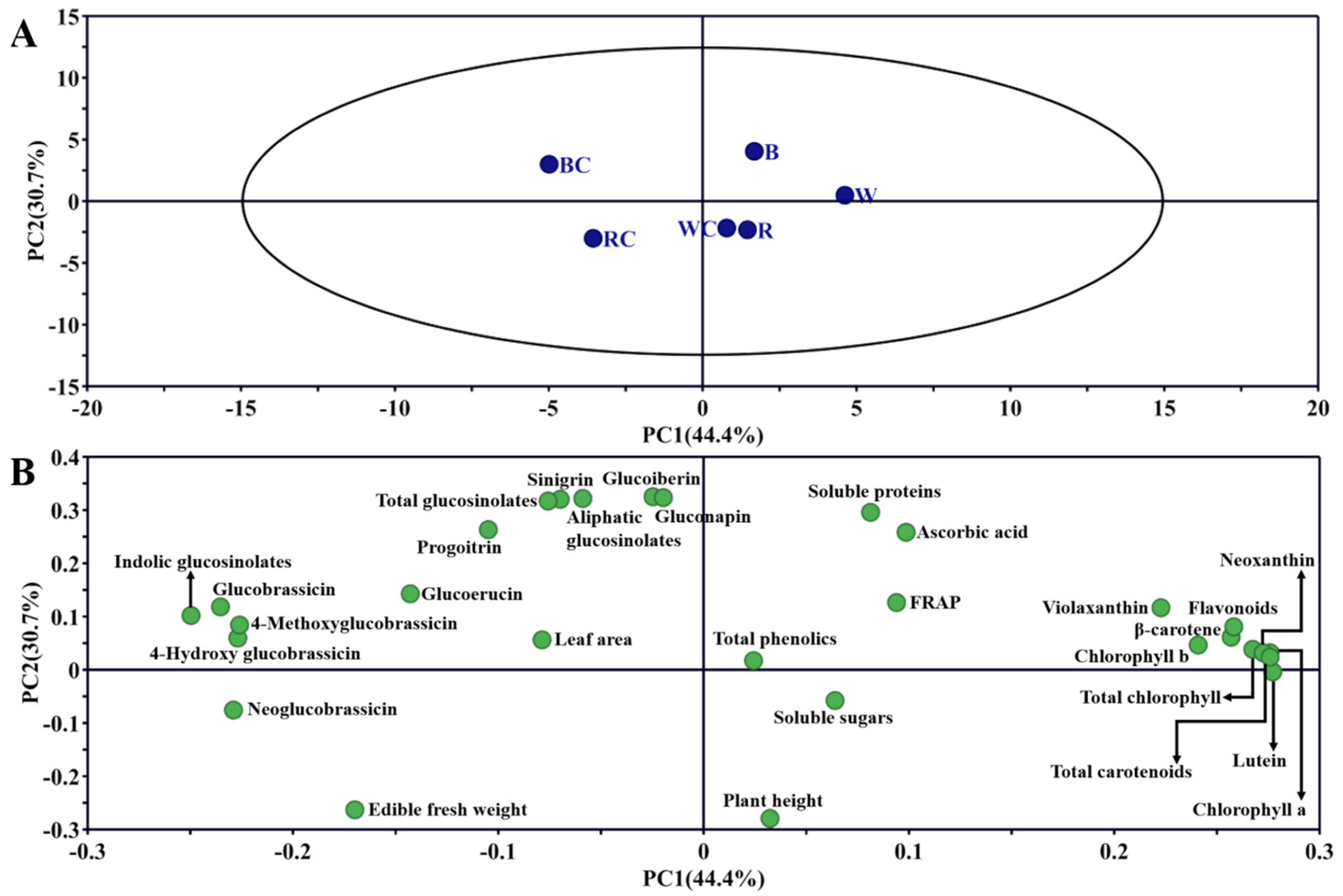
3.6. PCA
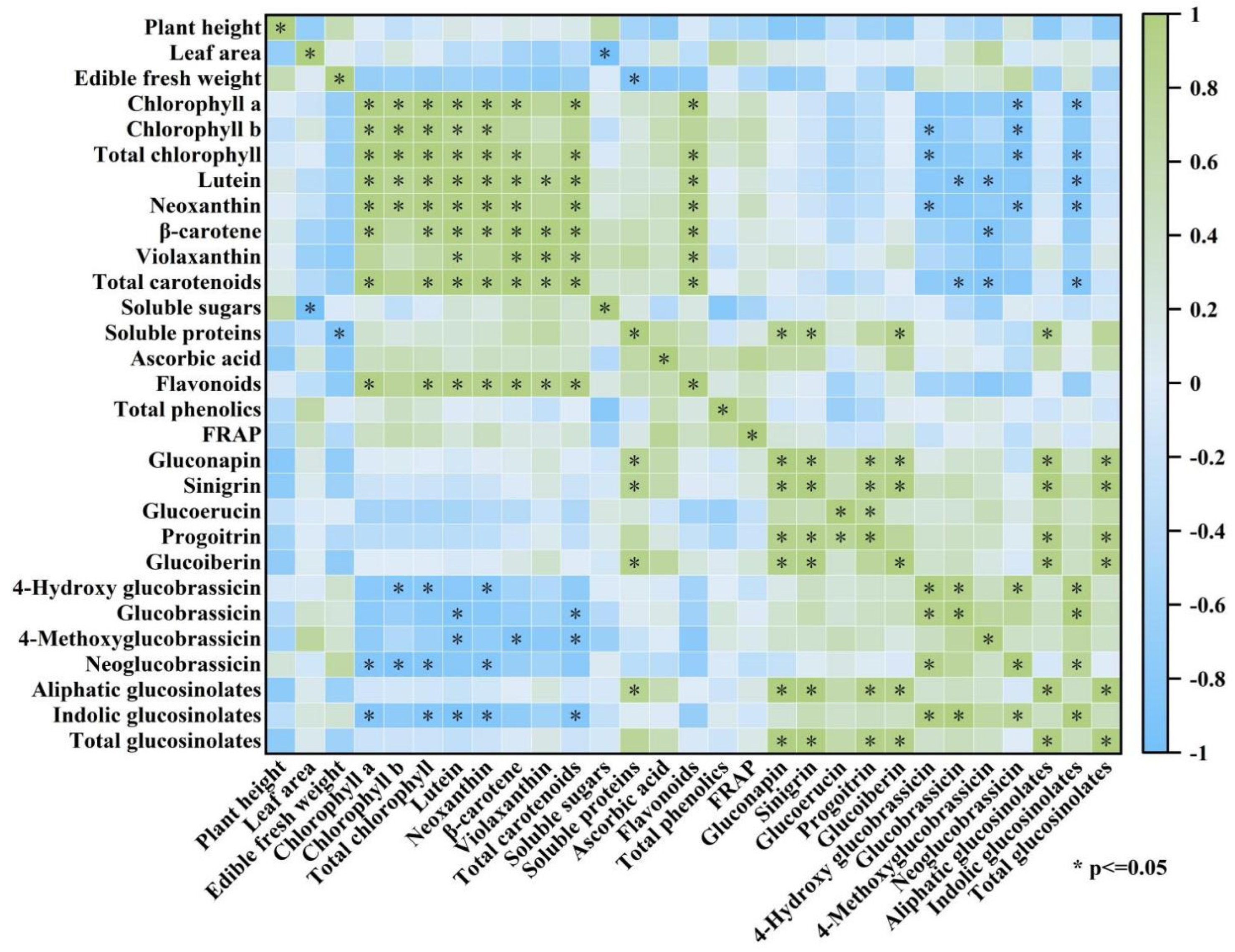
3.7. Correlation Analysis
3.8. Analysis of Variance
4. Discussion
4.1. The Effect of Different Light Qualities Alone and in Conjunction with Calcium Chloride on the Development of Chinese Kale Sprouts
4.2. The Effect of Different Light Qualities Alone and in Conjunction with Calcium Chloride on the Antioxidant Capacity of Chinese Kale Sprouts
4.3. The Effect of Different Light Qualities Alone and in Conjunction with Calcium Chloride on the Glucosinolate Content of Chinese Kale Sprouts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sun, B.; Liu, N.; Zhao, Y.T.; Yan, H.Z.; Wang, Q.M. Variation of glucosinolates in three edible parts of Chinese kale (Brassica alboglabra Bailey) varieties. Food Chem. 2011, 124, 941–947. [Google Scholar] [CrossRef]
- Chang, J.Q.; Wang, M.Y.; Jian, Y.; Zhang, F.; Zhu, J.; Wang, Q.M.; Sun, B. Health-promoting phytochemicals and antioxidant capacity in different organs from six varieties of Chinese kale. Sci. Rep. 2019, 9, 20344. [Google Scholar] [CrossRef] [PubMed]
- Hernández, R.; Kubota, C. Physiological responses of cucumber seedlings under different blue and red photon flux ratios using LEDs. Environ. Exp. Bot. 2016, 121, 66–74. [Google Scholar] [CrossRef]
- Rehman, M.; Ullah, S.; Bao, Y.N.; Wang, B.; Peng, D.X.; Liu, L.J. Light-emitting diodes: Whether an efficient source of light for indoor plants. Environ. Sci. Pollut. Res. 2017, 24, 24743–24752. [Google Scholar] [CrossRef]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Xu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Wang, L.W.; Li, Y.; Xin, G.F.; Wei, M.; Yang, Q.C.; Mi, Q.H. Effects of red and blue light quality on nitrogen levels, activities and gene expression of key enzymes involved in nitrogen metabolism from leaves of tomato seedlings. Acta Hortic. Sin. 2017, 44, 768–776. [Google Scholar]
- Kitayama, M.; Nguyen, D.T.P.; Lu, N.; Takagaki, M. Effect of light quality on physiological disorder, growth, and secondary metabolite content of water spinach (Ipomoea aquatica Forsk) cultivated in a closed-type plant production system. Hortic. Sci. Technol. 2019, 37, 206–218. [Google Scholar] [CrossRef]
- Abdul, M.; Amana, K.; Wajahat, K.; Dilsat, B.K.; Muhammad, M.A.; Muhammad, J.; Shafiq, U.R.; Baber, A.; Alevcan, K.; Sana, W. Effect of light emitting diodes (LEDs) on growth, mineral composition, and nutritional value of wheat & lentil sprouts. Phyton-Int. J. Exp. Bot. 2024, 93, 1117–1128. [Google Scholar]
- Chang, H.K.; Eun, K.Y.; Muthusamy, M.; Jin, A.K.; Mi-Jeong, J.; Soo, I.L. Blue LED light irradiation enhances L-ascorbic acid content while reducing reactive oxygen species accumulation in Chinese cabbage seedlings. Sci. Hortic. 2020, 261, 108924. [Google Scholar]
- Qian, H.M.; Liu, T.Y.; Deng, M.D.; Miao, H.Y.; Cai, C.X.; Shen, W.S.; Wang, Q.M. Effects of light quality on main health-promoting compounds and antioxidant capacity of Chinese kale sprouts. Food Chem. 2016, 196, 1232–1238. [Google Scholar] [CrossRef]
- Gao, Q.Y.; Xiong, T.T.; Li, X.P.; Chen, W.X.; Zhu, X.Y. Calcium and calcium sensors in fruit development and ripening. Sci. Hortic. 2019, 253, 412–421. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, S.; Wan, S.; Li, X. The significance of calcium in photosynthesis. Int. J. Mol. Sci. 2019, 20, 1353. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.K.; Yang, R.Q.; Zhou, Y.L.; Gu, Z.X. A comparative transcriptome and proteomics analysis reveals the positive effect of supplementary Ca2+ on soybean sprout yield and nutritional qualities. J. Proteom. 2016, 143, 161–172. [Google Scholar] [CrossRef]
- Yang, R.Q.; Hui, Q.R.; Gu, Z.X.; Zhou, Y.L.; Guo, L.P.; Shen, C.; Zhang, W.H. Effects of CaCl2 on the metabolism of glucosinolates and the formation of isothiocyanates as well as the antioxidant capacity of broccoli sprouts. J. Funct. Foods 2016, 24, 156–163. [Google Scholar] [CrossRef]
- Sun, J.H.; Kou, L.P.; Geng, P.; Huang, H.L.; Yang, T.B.; Luo, Y.G.; Chen, P. Metabolomic assessment reveals an elevated level of glucosinolate content in CaCl2 treated broccoli microgreens. J. Agric. Food Chem. 2015, 63, 1863–1868. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Georg, K.; Michael, M.; Joseph, A.M.H.; Erwin, L. Stromal free calcium concentration and light-mediated activation of chloroplast fructose-1,6-bisphosphatase. Plant Physiol. 1988, 86, 423–428. [Google Scholar]
- Zhu, C.G.; Hu, Z.J.; Hu, C.Y.; Ma, H.X.; Zhou, J.; Xia, X.J.; Shi, K.; Foyer, C.H.; Yu, J.Q.; Zhou, Y.H. SlCPK27 cross-links SlHY5 and SlPIF4 in brassinosteroid-dependent photo- and thermo-morphogenesis in tomato. Proc. Natl. Acad. Sci. USA 2024, 121, e2403040121. [Google Scholar] [CrossRef]
- Sun, B.; Di, H.M.; Zhang, J.Q.; Xia, P.X.; Huang, W.L.; Jian, Y.; Zhang, C.L.; Zhang, F. Effect of light on sensory quality, health-promoting phytochemicals and antioxidant capacity in post-harvest baby mustard. Food Chem. 2021, 339, 128057. [Google Scholar] [CrossRef]
- Sun, B.; Tian, Y.X.; Jiang, M.; Yuan, Q.; Chen, Q.; Zhang, Y.; Luo, Y.; Zhang, F.; Tang, H.R. Variation in the main health-promoting compounds and antioxidant activity of whole and individual edible parts of baby mustard (Brassica juncea var. gemmifera). RSC Adv. 2018, 8, 33845–33854. [Google Scholar] [CrossRef]
- Zhou, A.L.; Tang, J.Y.; Wu, S.; Di, H.M.; Ma, J.; Escalona, V.H.; Yu, X.N.; Liang, K.H.; Huang, Z.; Tang, Y.; et al. Low concentration of sodium chloride combined with melatonin improved the nutritional quality of Chinese kale sprouts. LWT-Food Sci. Technol. 2025, 225, 117903. [Google Scholar] [CrossRef]
- Zhou, A.L.; Zhang, Y.T.; Li, L.; Di, H.M.; Bian, J.L.; Ma, J.; Escalona, V.H.; Hong, H.J.; Li, H.X.; Tang, Y.; et al. Effects of combined treatment of light quality and melatonin on health-promoting compounds in Chinese kale sprouts. LWT-Food Sci. Technol. 2024, 199, 116137. [Google Scholar] [CrossRef]
- Artés-Hernández, F.; Castillejo, N.; Martínez-Zamora, L. UV and visible spectrum LED lighting as abiotic elicitors of bioactive compounds in sprouts, microgreens, and baby leaves—A comprehensive review including their mode of action. Foods 2022, 11, 265. [Google Scholar] [CrossRef]
- Akvilė, V.; Aušra, B.; Viktorija, V.K.; Jurga, M.; Julė, J.; Algirdas, N.; Kristina, L.; Giedrė, S. The distinct impact of multi-color LED light on nitrate, amino acid, soluble sugar and organic acid contents in red and green leaf lettuce cultivated in controlled environment. Food Chem. 2020, 310, 125799. [Google Scholar]
- Xu, F.; He, S.B.; Zhang, J.Y.; Mao, Z.L.; Wang, W.X.; Li, T.; Hua, J.; Du, S.S.; Xu, P.B.; Li, L.; et al. Photoactivated CRY1 and phyB interact directly with AUX/IAA proteins to inhibit auxin signaling in Arabidopsis. Mol. Plant 2018, 11, 523–541. [Google Scholar] [CrossRef]
- Adhikari, A.; Sapkota, M.; Savidya, R.N.; Tosin, A.T.; Adam, M.; Alam, M.N.; Kwon, E.-H.; Kang, S.-M.; Shaffique, S.; Lee, I.-J. Calcium enhances the effectiveness of melatonin in improving nutritional properties of soybean sprouts and germination under salt and cadmium stress. Int. J. Mol. Sci. 2025, 26, 878. [Google Scholar] [CrossRef]
- Gao, Y.; Dong, X.; Wang, R.; Hao, F.; Zhang, H.; Zhang, Y.; Lin, G. Exogenous calcium alleviates oxidative stress caused by salt stress in peanut seedling roots by regulating the antioxidant enzyme system and flavonoid biosynthesis. Antioxidants 2024, 13, 233. [Google Scholar] [CrossRef] [PubMed]
- Charlotte, M.M.G.; Eric, J.W.V.; Kate, R.S.O.; Laurentius, A.C.J.V.; Ronald, P. Shade tolerance: When growing tall is not an option. Trends Plant Sci. 2013, 18, 65–71. [Google Scholar]
- Park, Y.J.; Runkle, E.S. Far-red radiation promotes growth of seedlings by increasing leaf expansion and whole-plant net assimilation. Environ. Exp. Bot. 2017, 136, 41–49. [Google Scholar] [CrossRef]
- Weraduwage, S.M.; Chen, J.; Anozie, F.C.; Morales, A.; Weise, S.E.; Sharkey, T.D. The relationship between leaf area growth and biomass accumulation in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 167. [Google Scholar] [CrossRef]
- Gao, Y.F.; Zhou, X.; Huang, H.T.; Wang, C.; Xiao, X.X.; Wen, J.; Wu, J.M.; Zhou, S.; Víctor, R.D.; Lucas, G.R.; et al. ORANGE proteins mediate adaptation to high light and resistance to Pseudomonas syringae in tomato by regulating chlorophylls and carotenoids accumulation. Int. J. Biol. Macromol. 2025, 306, 141379. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.L.; Tang, J.Y.; Cheng, W.J.; Yao, X.W.; Escalona, V.H.; Qian, G.P.; Ma, J.; Yu, X.N.; Li, H.X.; Huang, Z.; et al. Combination of light quality and melatonin regulates the quality in mustard sprouts. Food Chem. 2024, 23, 101560. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.Y.; Xiang, N.; Chen, H.L.; Zhao, Y.H.; Wang, L.X.; Cheng, X.Z.; Guo, X.B. The modulation of light quality on carotenoid and tocochromanol biosynthesis in mung bean (Vigna radiata) sprouts. Food Chem.-Mol. Sci. 2023, 6, 100170. [Google Scholar] [CrossRef]
- Weinl, S.; Held, K.; Schlücking, K.; Steinhorst, L.; Kuhlgert, S.; Hippler, M.; Kudla, J. A plastid protein crucial for Ca2+-regulated stomatal responses. New Phytol. 2008, 179, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.K.; Chen, Y.Y.; Hu, T.T.; Zhang, S.J.; Zhang, Y.H.; Zhao, T.Y.; Yu, H.E.; Kang, Y.F. The influence of light-emitting diodes on the phenolic compounds and antioxidant activities in pea sprouts. J. Funct. Foods 2016, 25, 459–465. [Google Scholar] [CrossRef]
- Céline, B.; Kentaro, M.; Paul, D.; Guillaume, D.; Tim, B.; Jean-Philippe, M.; Joana, J.; Stéphanie, G.; Cédric, C.; Mickael, M.; et al. Blue light promotes ascorbate synthesis by deactivating the PAS/LOV photoreceptor that inhibits GDP-L-galactose phosphorylase. Plant Cell 2023, 35, 2615–2634. [Google Scholar]
- Zhao, Y.; Shi, H.; Pan, Y.; Lyu, M.H.; Yang, Z.X.; Kou, X.X.; Deng, X.W.; Zhong, S.W. Sensory circuitry controls cytosolic calcium-mediated phytochrome B phototransduction. Cell 2023, 186, 1230–1243. [Google Scholar] [CrossRef]
- Bansal, S.; Lakra, N.; Mishra, S.; Ahlawat, Y.K. Unraveling the potential of glucosinolates for nutritional enhancement and stress tolerance in Brassica crops. Veg. Res. 2024, 4, e015. [Google Scholar] [CrossRef]
- Mao, P.P.; Li, Q.M.; Li, Y.M.; Xu, Y.L.; Yang, Q.C.; Bian, Z.H.; Wang, S.; He, L.M.; Xu, Z.G.; Zheng, Y.J.; et al. The beneficial functions of blue light supplementary on the biosynthesis of glucosinolates in pakchoi (Brassica rapa L. ssp. chinensis) under greenhouse conditions. Environ. Exp. Bot. 2022, 197, 104834. [Google Scholar]
- Köksal, D.; Gölge, S.; Gamze, Ç.S. Effect of LED lights on the growth, nutritional quality and glucosinolate content of broccoli, cabbage and radish microgreens. Food Chem. 2023, 401, 134088. [Google Scholar]
- Xue, A.H.; Liang, W.J.; Wen, S.D.; Gao, Y.Y.; Huang, X.Y.; Tong, Y.Z.; Hao, Y.B.; Luo, L.P. Metabolomic analysis based on EESI-MS indicate blue LED light promotes aliphatic-glucosinolates biosynthesis in broccoli sprouts. J. Food Compos. Anal. 2021, 97, 103777. [Google Scholar] [CrossRef]
- Agerbirk, N.; De, V.M.; Kim, J.H. Indole glucosinolate breakdown and its biological effects. Phytochem. Rev. 2009, 8, 101–120. [Google Scholar] [CrossRef]
- Wang, C.; Tang, R.J.; Kou, S.H.; Xu, X.S.; Lu, Y.; Rauscher, K.; Voelfer, A.; Luan, S. Mechanisms of calcium homeostasis orchestrate plant growth and immunity. Nature 2024, 627, 382–388. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Each Treatment in the Experiment | White Light | White Light + 10 mM Calcium Chloride Solution | Red Light | Red Light + 10 mM Calcium Chloride Solution | Blue Light | Blue Light + 10 mM Calcium Chloride Solution |
|---|---|---|---|---|---|---|
| Abbrevaition | W | WC | R | RC | B | BC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Cheng, W.; Yu, X.; Liang, K.; Tang, Y.; Huang, Z.; Li, H.; Zhang, F.; Chen, C.; Sun, B. Effect of Combined Light Quality and Calcium Chloride Treatments on Growth and Quality of Chinese Kale Sprouts. Horticulturae 2025, 11, 780. https://doi.org/10.3390/horticulturae11070780
Li H, Cheng W, Yu X, Liang K, Tang Y, Huang Z, Li H, Zhang F, Chen C, Sun B. Effect of Combined Light Quality and Calcium Chloride Treatments on Growth and Quality of Chinese Kale Sprouts. Horticulturae. 2025; 11(7):780. https://doi.org/10.3390/horticulturae11070780
Chicago/Turabian StyleLi, Hongxia, Wenjuan Cheng, Xuena Yu, Kehao Liang, Yi Tang, Zhi Huang, Huanxiu Li, Fen Zhang, Cunkun Chen, and Bo Sun. 2025. "Effect of Combined Light Quality and Calcium Chloride Treatments on Growth and Quality of Chinese Kale Sprouts" Horticulturae 11, no. 7: 780. https://doi.org/10.3390/horticulturae11070780
APA StyleLi, H., Cheng, W., Yu, X., Liang, K., Tang, Y., Huang, Z., Li, H., Zhang, F., Chen, C., & Sun, B. (2025). Effect of Combined Light Quality and Calcium Chloride Treatments on Growth and Quality of Chinese Kale Sprouts. Horticulturae, 11(7), 780. https://doi.org/10.3390/horticulturae11070780