
Genome-Wide Identification and Characterization of Trehalose-6-Phosphate Synthase/Phosphatases Gene Family in Petunia and Their Expression Profiling Under Abiotic Stresses



Abstract
1. Introduction
2. Materials and Methods
2.1. TPS and TPP Gene Identification in Petunia
2.2. Phylogenetic Analysis of TPS and TPP Genes
2.3. Gene Structure and Conserved Motif Analysis of TPS and TPP Genes
2.4. Cis-Element Analysis
2.5. Plant Materials, Growth Conditions, and Stress Treatment
2.6. Total RNA Extraction and qRT-PCR Analysis
2.7. Data Analysis
3. Results
3.1. Identification and Physicochemical Property Analysis of TPS and TPP Genes in Petunia
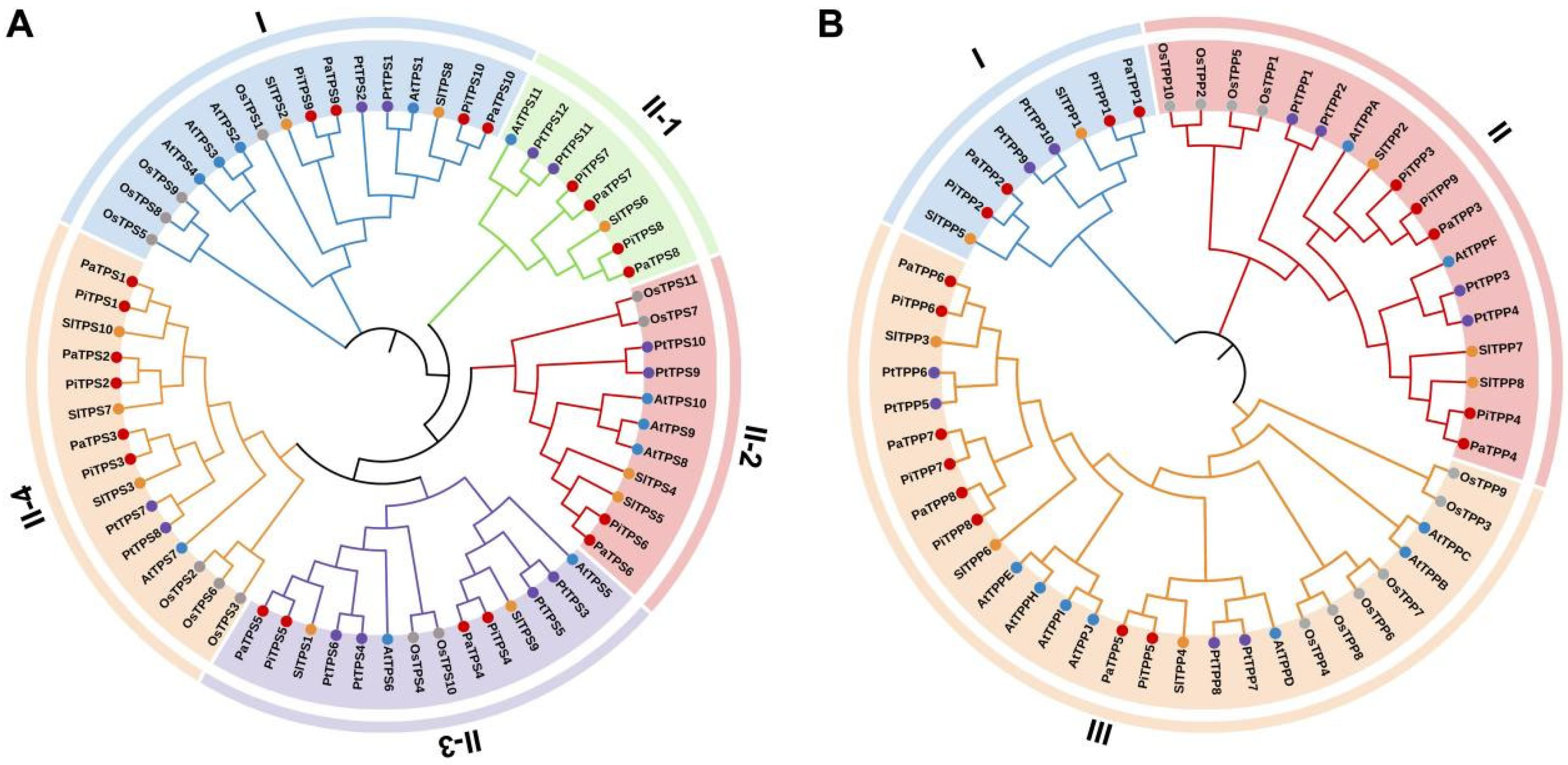
3.2. Phylogenetic Analysis of TPS and TPP Family Genes in Petunia
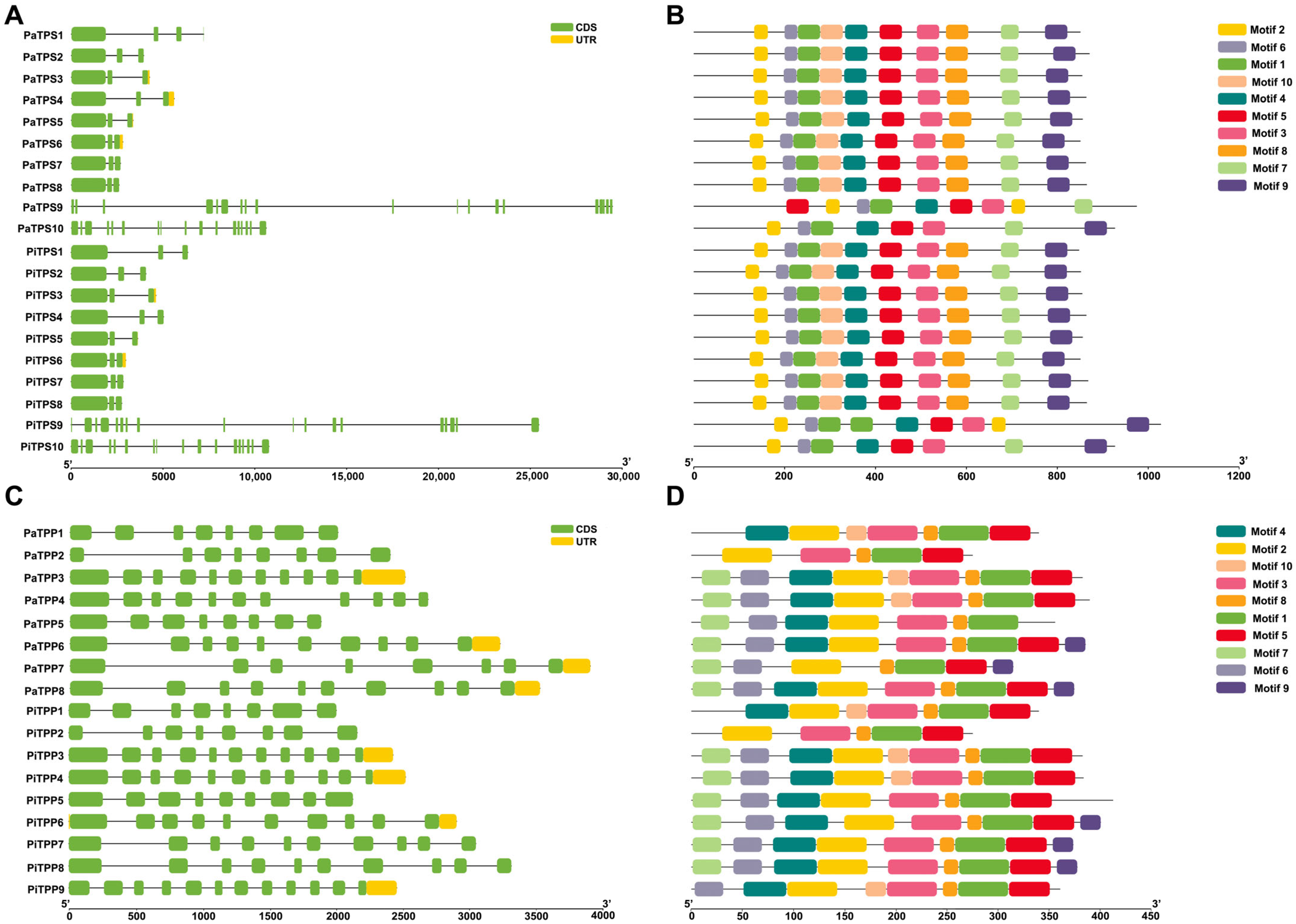
3.3. Gene Structure and Conserved Motif Analysis of TPSs and TPPs
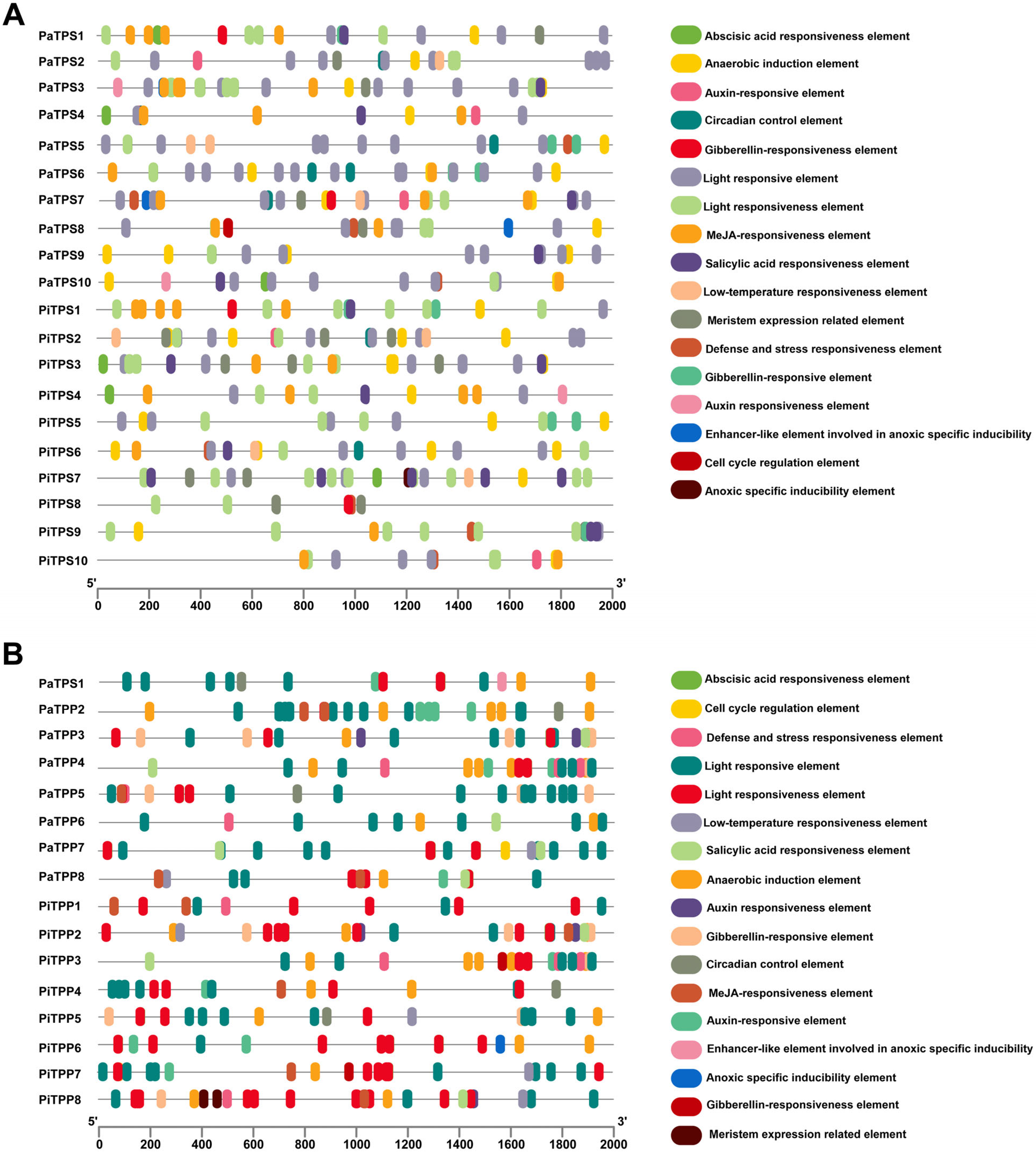
3.4. Cis-Acting Elements Analysis of TPSs and TPPs
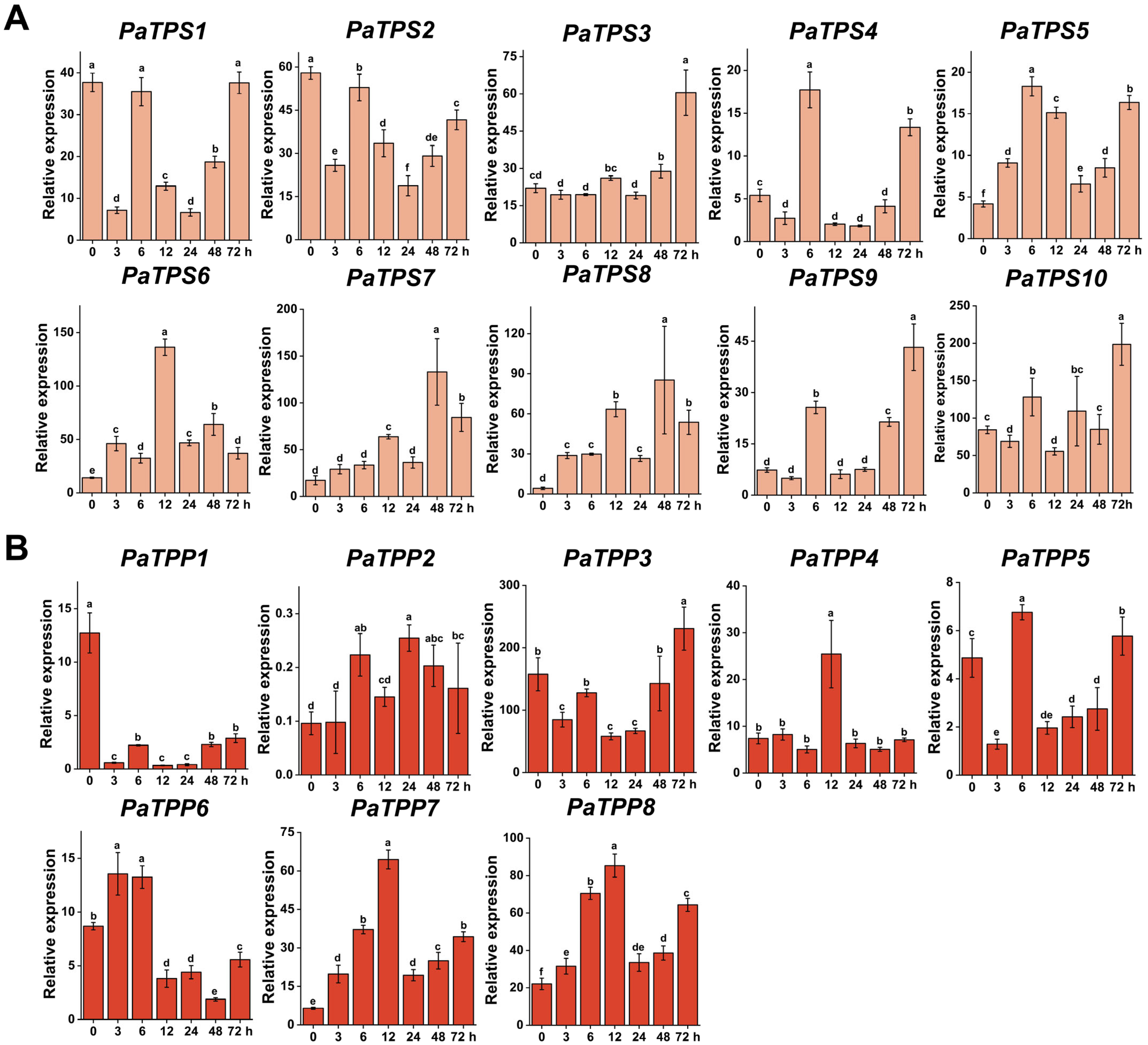
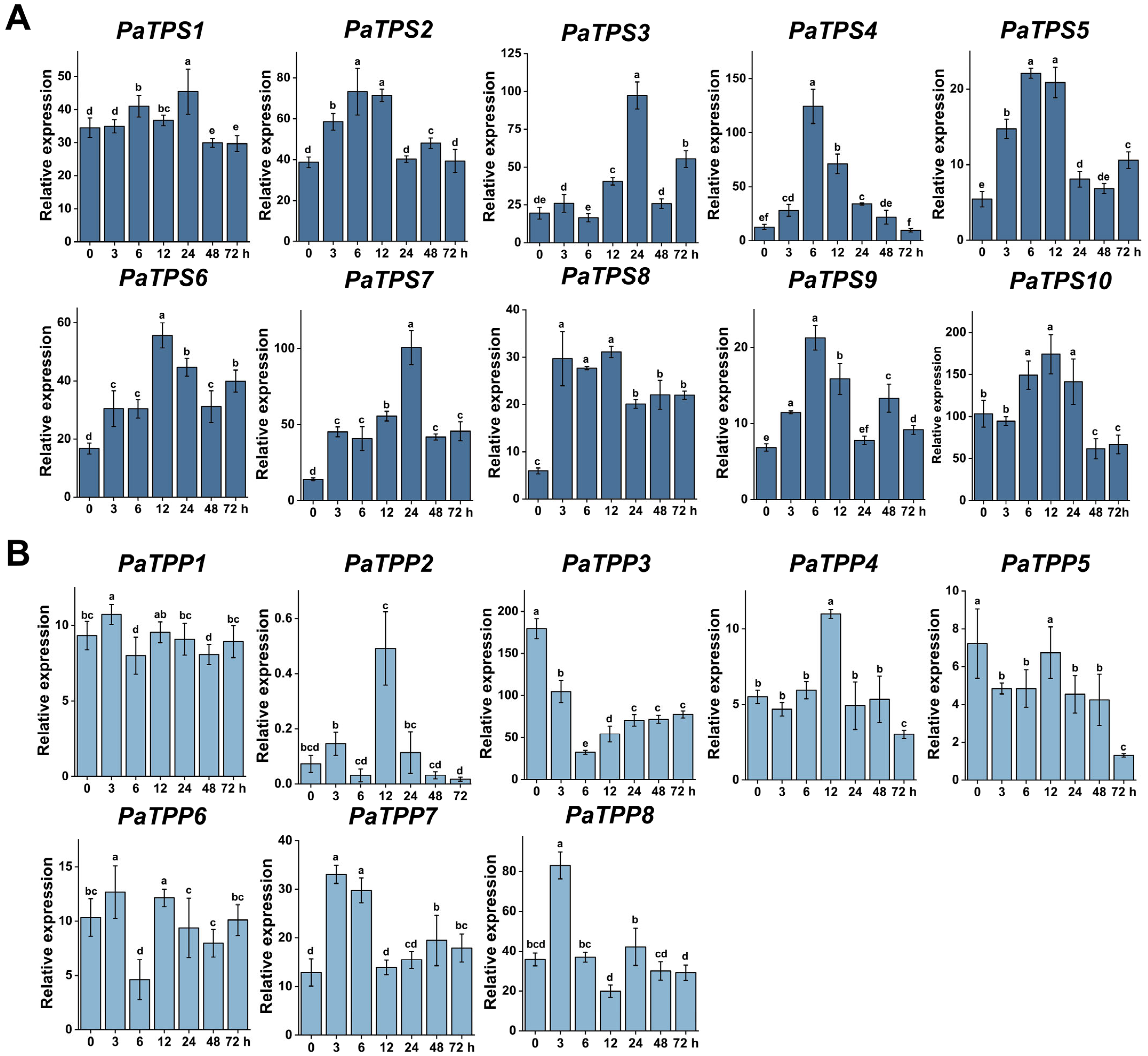
3.5. TPSs and TPPs Expression Pattern Analysis Under Different Stresses
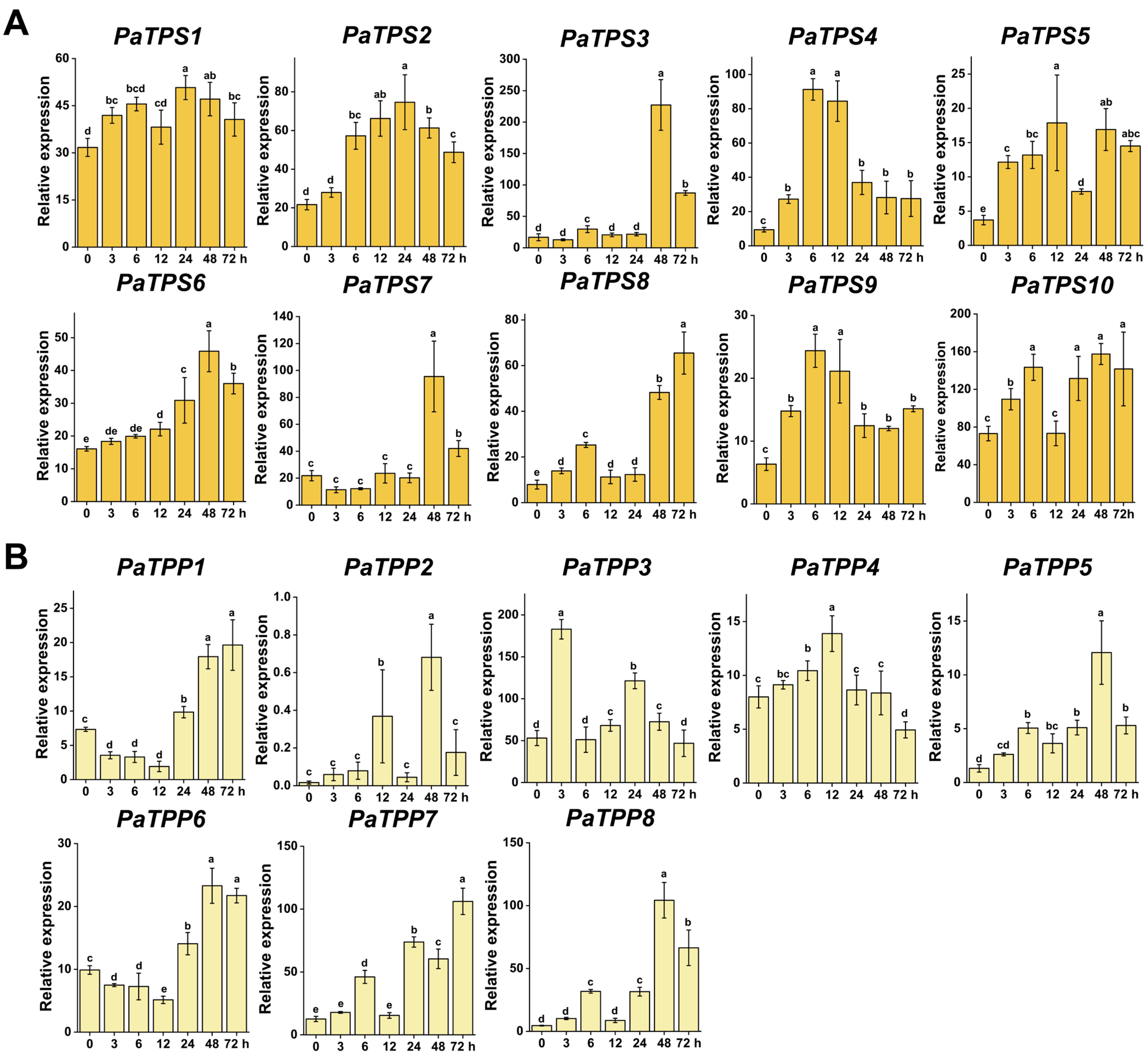
3.5.1. Drought Stress
3.5.2. Heat Stress
3.5.3. Salt Stress
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Paul, M.; Pellny, T.; Goddijn, O. Enhancing photosynthesis with sugar signals. Trends Plant Sci. 2001, 6, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Elbein, A.D.; Pan, Y.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, L.E.; Paul, M.J.; Wingler, A. How do sugars regulate plant growth and development? New insight into the role of trehalose-6-phosphate. Mol. Plant 2013, 6, 261–274. [Google Scholar] [CrossRef]
- Fernandez, O.; Bethencourt, L.; Quero, A.; Sangwan, R.S.; Clement, C. Trehalose and plant stress responses: Friend or foe? Trends Plant Sci. 2010, 15, 409–417. [Google Scholar] [CrossRef]
- Kosar, F.; Akram, N.A.; Sadiq, M.; Al-Qurainy, F.; Ashraf, M. Trehalose: A key organic osmolyte effectively involved in plant abiotic stress tolerance. J. Plant Growth Regul. 2019, 38, 606–618. [Google Scholar] [CrossRef]
- Villarino, G.H.; Bombarely, A.; Giovannoni, J.J.; Scanlon, M.J.; Mattson, N.S. Transcriptomic analysis of Petunia hybrida in response to salt stress using high throughput RNA sequencing. PLoS ONE 2014, 9, e94651. [Google Scholar] [CrossRef]
- Villarino, G.; Hu, Q.W.; Scanlon, M.; Mueller, L.A.; Bombarely, A.; Mattson, N.S. Dissecting tissue-specific transcriptomic responses from leaf and roots under salt stress in Petunia hybrida Mitchell. Genes 2017, 8, 195. [Google Scholar] [CrossRef]
- Smeekens, S. From leaf to kernel: Trehalose-6-phosphate signaling moves carbon in the field. Plant Physiol. 2015, 169, 912–913. [Google Scholar] [CrossRef]
- Avonce, N.; Mendoza-Vargas, A.; Morett, E.; Iturriaga, G. Insights on the evolution of trehalose biosynthesis. BMC Evol. Biol. 2006, 6, 109. [Google Scholar] [CrossRef]
- Vandesteene, L.; Ramon, M.; Roy, K.L.; Dijck, P.V.; Rolland, F. A single active trehalose-6-P synthase (TPS) and a family of putative regulatory TPS-like proteins in Arabidopsis. Mol. Plant 2010, 3, 406–419. [Google Scholar] [CrossRef]
- Vandesteene, L.; Lopez-Galvis, L.; Vanneste, K.; Feil, R.; Maere, S.; Lammens, W. Expansive evolution of the trehalose-6-phosphate phosphatase gene family in Arabidopsis. Plant Physiol. 2012, 160, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.F.; Chao, D.Y.; Shi, M.; Zhu, M.Z.; Gao, J.P.; Lin, H.X. Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes. Planta 2008, 228, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Zang, B.; Li, H.; Li, W.; Deng, X.W.; Wang, X. Analysis of trehalose-6-phosphate synthase (TPS) gene family suggests the formation of TPS complexes in rice. Plant Mol. Biol. 2011, 76, 507–522. [Google Scholar] [CrossRef]
- Dijck, P.V. The cytophaga hutchinsonii chtpsp: First characterized bifunctional TPS-TPP protein as putative ancestor of all eukaryotic trehalose biosynthesis proteins. Mol. Biol. Evol. 2010, 27, 359–369. [Google Scholar]
- Shima, S.; Matsui, H.; Tahara, Y.; Imai, R. Biochemical characterization of rice trehalose-6-phosphate phosphatases supports distinctive functions of these plant enzymes. FEBS J. 2007, 274, 1192–1201. [Google Scholar] [CrossRef]
- Iordachescu, M.; Imai, R. Trehalose biosynthesis in response to abiotic stresses. J. Integr. Plant Biol. 2008, 50, 1223–1229. [Google Scholar] [CrossRef]
- Li, H.W.; Zang, B.S.; Deng, X.W.; Wang, X.P. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 2011, 234, 1007–1018. [Google Scholar] [CrossRef]
- Jiang, D.; Chen, W.; Gao, J.; Yang, F.; Zhuang, C. Overexpression of the trehalose-6-phosphate phosphatase OsTPP3 increases drought tolerance in rice. Plant Biotechnol. Rep. 2019, 13, 285–292. [Google Scholar] [CrossRef]
- Lin, Q.; Wang, S.; Dao, Y.; Wang, J.; Wang, K. The Arabidopsis thaliana trehalose-6-phosphate phosphatase gene AtTPP1 enhances drought tolerance by regulating stomatal apertures. J. Exp. Bot. 2020, 71, 1795–1807. [Google Scholar] [CrossRef]
- Lin, Q.; Yang, J.; Wang, Q.; Zhu, H.; Chen, Z.; Dao, Y.; Wang, K. Overexpression of the trehalose-6-phosphate phosphatase family gene AtTPPF improves the drought tolerance of Arabidopsis thaliana. BMC Plant Biol. 2019, 19, 381. [Google Scholar] [CrossRef]
- Li, J.; Guo, H.F.; Lou, Q.J.; Zeng, Y.W.; Guo, Z.H.; Xu, P.H.; Gu, Y.; Gao, S.; Xu, B.; Han, S.; et al. Natural variation of indels in the CTB3 promoter confers cold tolerance in japonica rice. Nat. Commun. 2025, 16, 1613. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Guo, Y.F.; Yang, Q.; He, Y.H.; Fetouh, M.I.; Warner, R.M.; Deng, Z. Genome-wide identification of quantitative trait loci for important plant and flower traits in petunia using a high-density linkage map and an interspecific recombinant inbred population derived from Petunia integrifolia and P. axillaris. Hortic. Res. 2019, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Zentella, R.; Mascorro-Gallardo, J.O.; Van Dijck, P.; Folch-Mallol, J.; Bonini, B.; Van Vaeck, C.; Gaxiola, R.; Covarrubias, A.A.; Nieto-Sotelo, J.; Thevelein, J.M.; et al. A Selaginella lepidophylla trehalose-6-phosphate synthase complements growth and stress-tolerance defects in a yeast tps1 mutant. Plant Physiol. 1999, 119, 1473–1482. [Google Scholar] [CrossRef]
- Bombarely, A.; Moser, M.; Amrad, A.; Bao, M.; Bapaume, L.; Barry, C.S.; Bliek, M.; Boersma, M.R.; Borghi, L.; Bruggmann, R.; et al. Insight into the evolution of the Solanaceae from the parental genomes of Petunia hybrida. Nat. Plants 2016, 2, 16074. [Google Scholar] [CrossRef]
- Berardi, A.E.; Korinna, E.; Lea, J.; Therese, M.; Cannarozzi, G.M.; Cris, K. Complex evolution of novel red floral color in petunia. Plant Cell 2021, 33, 2273–2295. [Google Scholar] [CrossRef]
- Patrick, R.M.; Xing-Qi, H.; Natalia, D.; Ying, L. Dynamic histone acetylation in floral volatile synthesis and emission in petunia flowers. J. Exp. Bot. 2021, 72, 3704–3718. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003, 2, 2–3. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, 265–268. [Google Scholar] [CrossRef]
- Ivica, L.; Peer, B. Interactive tree of life (iTOL) v6: Recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res. 2024, 52, W78–W82. [Google Scholar]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Peer, Y.V.D.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Livak, K.L.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sarkar, A.K.; Sadhukhan, S. Imperative role of trehalose metabolism and trehalose-6-phosphate signaling on salt stress responses in plants. Physiol. Plant. 2022, 174, e13647. [Google Scholar] [CrossRef]
- Fichtner, F.; Lunn, J.E. The role of trehalose 6-phosphate (tre6p) in plant metabolism and development. Annu. Rev. Plant Biol. 2021, 72, 737–760. [Google Scholar] [CrossRef]
- Zhao, L.; Zhao, X.; Francis, F.; Liu, Y. Genome-wide identification and characterization of the TPS gene family in wheat (Triticum aestivum L.) and expression analysis in response to aphid damage. Acta Physiol. Plant. 2021, 43, 64. [Google Scholar] [CrossRef]
- Du, L.; Li, S.; Ding, L.; Cheng, X.; Kang, Z.; Mao, H. Genome-wide analysis of trehalose-6-phosphate phosphatases (TPP) gene family in wheat indicates their roles in plant development and stress response. BMC Plant Biol. 2022, 22, 120. [Google Scholar] [CrossRef]
- Mollavali, M.; Börnke, F. Characterization of trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase genes of tomato (Solanum lycopersicum L.) and analysis of their differential expression in response to temperature. Int. J. Mol. Sci. 2022, 23, 11436. [Google Scholar] [CrossRef]
- Zhong, C.; He, Z.; Liu, Y.; Li, Z.; Wang, X.; Jiang, C.; Kang, S.; Liu, X.; Zhao, S.; Wang, J.; et al. Genome-wide identification of TPS and TPP genes in cultivated peanut (Arachis hypogaea) and functional characterization of AtTPS9 in response to cold stress. Front. Plant Sci. 2024, 14, 1343402. [Google Scholar] [CrossRef]
- Doganlar, S.; Frary, A.; Daunay, M.C.; Lester, R.N.; Tanksley, S.D. Conservation of gene function in the solanaceae as revealed by comparative mapping of domestication traits in eggplant. Genetics 2002, 161, 1713–1726. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Chen, L.; Wang, C.; Zhang, S.; Yang, Y.; Liu, J.; Li, D.; Song, J.; Wang, R. Characterization of the auxin efflux transporter PIN proteins in pear. Plants 2020, 9, 349. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, D.; Luo, S.; Zhu, Y.; Jia, X.; Duan, Y.; Zhou, M. Intron-mediated regulation of β-tubulin genes expression affects the sensitivity to carbendazim in Fusarium graminearum. Curr. Genet. 2019, 65, 1057–1069. [Google Scholar] [CrossRef]
- Paul, M.J.; Primavesi, L.F.; Jhurreea, D.; Zhang, Y. Trehalose metabolism and signaling. Annu. Rev. Plant Biol. 2008, 59, 417–441. [Google Scholar] [CrossRef]
- Jain, N.K.; Roy, I. Effect of trehalose on protein structure. Protein Sci. 2010, 18, 24–36. [Google Scholar] [CrossRef]
- Sadak, M.S.; El-Bassiouny, H.M.S.; Dawood, M.G. Role of trehalose on antioxidant defense system and some osmolytes of quinoa plants under water deficit. Bull. Natl. Res. Cent. 2019, 43, 5. [Google Scholar] [CrossRef]
- Yang, Y.; Yao, Y.; Li, J.; Zhang, J.; Zhang, X.; Hu, L.; Ding, D.; Bakpa, E.P.; Xie, J. Trehalose alleviated salt stress in tomato by regulating ros metabolism, photosynthesis, osmolyte synthesis, and trehalose metabolic pathways. Front. Plant Sci. 2022, 13, 772948. [Google Scholar] [CrossRef]
- Nawaz, M.; Hassan, M.U.; Chattha, M.U.; Mahmood, A.; Shah, A.N.; Hashem, M.; Alamri, S.; Batool, M.; Rasheed, A.; Thabit, M.A.; et al. Trehalose: A promising osmo-protectant against salinity stress-physiological and molecular mechanisms and future prospective. Mol. Biol. Rep. 2022, 49, 11255–11271. [Google Scholar] [CrossRef]
- Schluepmann, H.; Berke, L.; Sanchez-Perez, G.F. Metabolism control over growth: A case for trehalose-6-phosphate in plants. J. Exp. Bot. 2012, 63, 3379–3390. [Google Scholar] [CrossRef]
- Paul, M. Trehalose 6-phosphate. Curr. Opin. Plant Biol. 2007, 3, 303–309. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus_ID | Position | Length (AA) | MW (Da) | pI | Subcellular Localization | ||
|---|---|---|---|---|---|---|---|---|
| Scaffold Location | Start | End | ||||||
| PaTPS1 | Peaxi162Scf00229g00096.1 | Peaxi162Scf00229 | 917,477 | 925,133 | 851 | 96.93 | 5.58 | Cytoplasmic |
| PaTPS2 | Peaxi162Scf00469g00632.1 | Peaxi162Scf00469 | 667,063 | 671,245 | 871 | 98.92 | 5.72 | Nuclear |
| PaTPS3 | Peaxi162Scf00202g00187.1 | Peaxi162Scf00202 | 1,815,219 | 1,819,743 | 855 | 96.39 | 5.70 | Cytoplasmic |
| PaTPS4 | Peaxi162Scf00170g01223.1 | Peaxi162Scf00170 | 1,214,499 | 1,220,448 | 864 | 97.52 | 5.78 | Cytoplasmic |
| PaTPS5 | Peaxi162Scf00275g00085.1 | Peaxi162Scf00275 | 813,026 | 816,592 | 856 | 97.16 | 5.66 | Cytoplasmic |
| PaTPS6 | Peaxi162Scf00421g00116.1 | Peaxi162Scf00421 | 104,373 | 107,349 | 851 | 96.23 | 5.91 | Cytoplasmic |
| PaTPS7 | Peaxi162Scf00297g00098.1 | Peaxi162Scf00297 | 982,826 | 985,666 | 863 | 97.97 | 6.01 | Cytoplasmic |
| PaTPS8 | Peaxi162Scf00457g00215.1 | Peaxi162Scf00457 | 252,951 | 255,711 | 865 | 98.05 | 5.96 | Cytoplasmic |
| PaTPS9 | Peaxi162Scf00022g02413.1 | Peaxi162Scf00022 | 2,390,346 | 2,421,693 | 975 | 110.65 | 6.03 | Cytoplasmic |
| PaTPS10 | Peaxi162Scf00111g01324.1 | Peaxi162Scf00111 | 1,308,982 | 1,320,265 | 927 | 104.39 | 6.29 | Cytoplasmic |
| PiTPS1 | Peinf101Scf00275g03007.1 | Peinf101Scf00275 | 406,279 | 412,631 | 848 | 96.55 | 5.67 | Cytoplasmic |
| PiTPS2 | Peinf101Scf00442g03004.1 | Peinf101Scf00442 | 320,610 | 324,681 | 852 | 96.75 | 5.77 | Nuclear |
| PiTPS3 | Peinf101Scf00994g03007.1 | Peinf101Scf00994 | 331,522 | 336,139 | 855 | 96.37 | 5.65 | Cytoplasmic |
| PiTPS4 | Peinf101Scf01033g01017.1 | Peinf101Scf01033 | 150,487 | 155,519 | 864 | 97.51 | 5.78 | Cytoplasmic |
| PiTPS5 | Peinf101Scf01456g02005.1 | Peinf101Scf01456g | 225,034 | 228,660 | 856 | 97.16 | 5.66 | Cytoplasmic |
| PiTPS6 | Peinf101Scf01436g11018.1 | Peinf101Scf01436 | 1,108,249 | 1,111,224 | 851 | 96.25 | 5.91 | Cytoplasmic |
| PiTPS7 | Peinf101Scf00586g01015.1 | Peinf101Scf00586 | 115,175 | 118,018 | 868 | 98.67 | 6.01 | Cytoplasmic |
| PiTPS8 | Peinf101Scf01850g03027.1 | Peinf101Scf01850 | 402,667 | 405,426 | 865 | 97.92 | 5.99 | Cytoplasmic |
| PiTPS9 | Peinf101Scf00304g01004.1 | Peinf101Scf00304 | 68,073 | 93,541 | 1028 | 116.45 | 7.19 | Cytoplasmic |
| PiTPS10 | Peinf101Scf00665g07025.1 | Peinf101Scf00665 | 722,849 | 733,623 | 927 | 104.39 | 6.29 | Cytoplasmic |
| Gene Name | Locus_ID | Position | Length (AA) | MW (Da) | pI | Subcellular Localization | ||
|---|---|---|---|---|---|---|---|---|
| Scaffold Location | Start | End | ||||||
| PaTPP1 | Peaxi162Scf00904g00003.1 | Peaxi162Scf00904 | 81,668 | 83,679 | 341 | 38.06 | 6.04 | Cytoplasmic |
| PaTPP2 | Peaxi162Scf00711g00011.1 | Peaxi162Scf00711 | 115,934 | 118,338 | 276 | 31.56 | 6.62 | Cytoplasmic |
| PaTPP3 | Peaxi162Scf00241g00319.1 | Peaxi162Scf00241 | 378,645 | 381,161 | 384 | 42.76 | 6.72 | Chloroplast |
| PaTPP4 | Peaxi162Scf00002g00816.1 | Peaxi162Scf00002 | 860,185 | 862,872 | 391 | 44.18 | 7.66 | Mitochondrial |
| PaTPP5 | Peaxi162Scf00041g00810.1 | Peaxi162Scf00041 | 797,885 | 799,770 | 357 | 40.06 | 9.13 | Chloroplast |
| PaTPP6 | Peaxi162Scf00838g00003.1 | Peaxi162Scf00838 | 86,470 | 89,698 | 387 | 43.66 | 9.4 | Chloroplast |
| PaTPP7 | Peaxi162Scf00295g00075.1 | Peaxi162Scf00295 | 757,573 | 761,478 | 316 | 35.94 | 9.65 | Mitochondrial |
| PaTPP8 | Peaxi162Scf00295g00510.1 | Peaxi162Scf00295 | 578,702 | 582,229 | 376 | 42.29 | 9.22 | Chloroplast |
| PiTPP1 | Peinf101Scf01632g02002.1 | Peinf101Scf01632 | 229,134 | 231,134 | 341 | 38.03 | 6.04 | Cytoplasmic |
| PiTPP2 | Peinf101Scf03315g00014.1 | Peinf101Scf03315 | 71,337 | 73,493 | 276 | 31.47 | 6.41 | Cytoplasmic |
| PiTPP3 | Peinf101Scf03555g01022.1 | Peinf101Scf03555 | 91,639 | 94,062 | 384 | 42.73 | 6.72 | Chloroplast |
| PiTPP4 | Peinf101Scf00674g06014.1 | Peinf1011Scf00674 | 660,658 | 663,175 | 385 | 43.56 | 7.66 | Mitochondrial |
| PiTPP5 | Peinf101Scf00173g02008.1 | Peinf1011Scf00173 | 222,878 | 225,001 | 414 | 46.45 | 9.15 | Cytoplasmic |
| PiTPP6 | Peinf101Scf00966g00008.1 | Peinf101Scf00966 | 73,267 | 76,165 | 402 | 45.63 | 9.51 | Mitochondrial |
| PiTPP7 | Peinf101Scf01178g05006.1 | Peinf101Scf01178 | 522,267 | 525,309 | 375 | 42.50 | 9.18 | Nuclear |
| PiTPP8 | Peinf101Scf01178g06005.1 | Peinf101Scf01178 | 666,203 | 669,510 | 379 | 42.76 | 9.01 | Nuclear |
| PiTPP9 | Peinf101Scf07373g00001.1 | Peinf101Scf07373 | 3 | 2454 | 362 | 40.58 | 6.12 | Chloroplast |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, R.; Liu, D.; Villarino, G.H.; Mattson, N.S. Genome-Wide Identification and Characterization of Trehalose-6-Phosphate Synthase/Phosphatases Gene Family in Petunia and Their Expression Profiling Under Abiotic Stresses. Horticulturae 2025, 11, 695. https://doi.org/10.3390/horticulturae11060695
Huang R, Liu D, Villarino GH, Mattson NS. Genome-Wide Identification and Characterization of Trehalose-6-Phosphate Synthase/Phosphatases Gene Family in Petunia and Their Expression Profiling Under Abiotic Stresses. Horticulturae. 2025; 11(6):695. https://doi.org/10.3390/horticulturae11060695
Chicago/Turabian StyleHuang, Renwei, Daofeng Liu, Gonzalo H. Villarino, and Neil S. Mattson. 2025. "Genome-Wide Identification and Characterization of Trehalose-6-Phosphate Synthase/Phosphatases Gene Family in Petunia and Their Expression Profiling Under Abiotic Stresses" Horticulturae 11, no. 6: 695. https://doi.org/10.3390/horticulturae11060695
APA StyleHuang, R., Liu, D., Villarino, G. H., & Mattson, N. S. (2025). Genome-Wide Identification and Characterization of Trehalose-6-Phosphate Synthase/Phosphatases Gene Family in Petunia and Their Expression Profiling Under Abiotic Stresses. Horticulturae, 11(6), 695. https://doi.org/10.3390/horticulturae11060695