

bHLH291 Regulates Anthocyanin Accumulation in Kiwifruit Under Bagging Treatment


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
2.2. Extraction and Quantification of Anthocyanins
2.3. Extraction of Total RNA from Kiwifruit Samples
2.4. Transcriptome Sequencing
2.5. Phylogenetic Tree Analysis
2.6. First-Strand cDNA Synthesis and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.7. Dual-Luciferase Reporter Assay
2.8. Statistical Analyses
3. Results
3.1. Anthocyanin Content in Kiwifruit at Different Developmental Stages
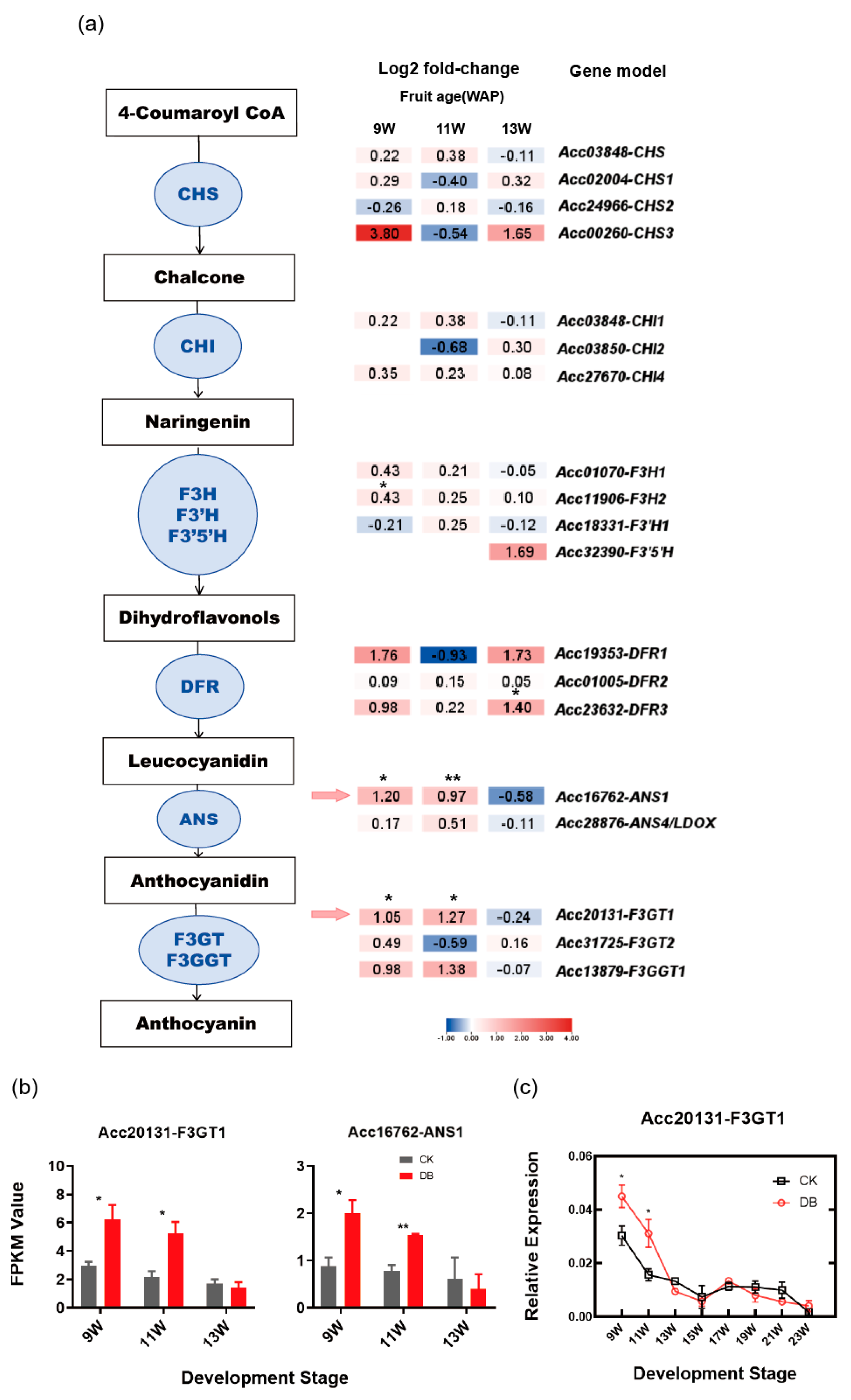
3.2. Identification of Anthocyanin Biosynthesis Structural Genes Responding to Bagging Treatment
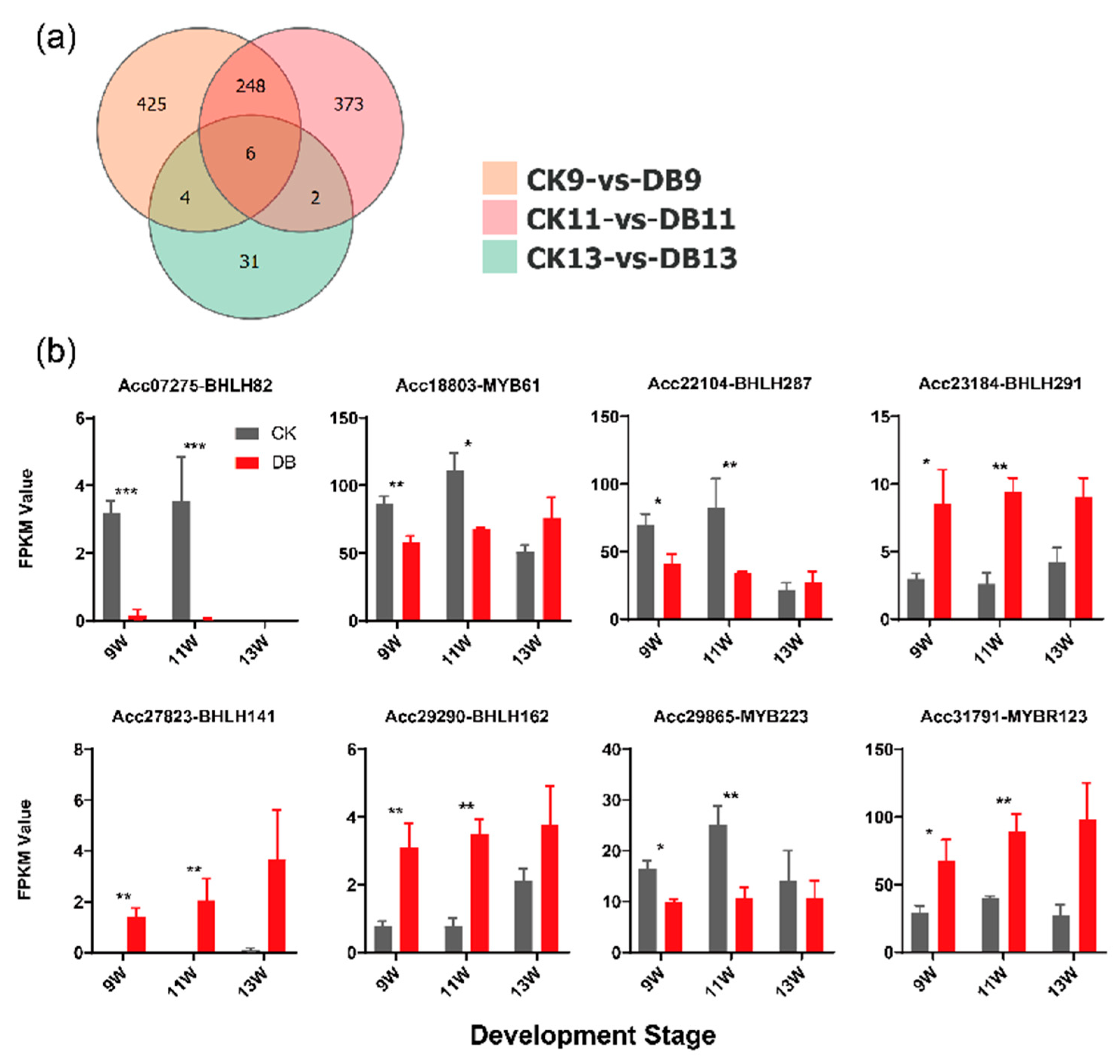
3.3. Determination of Potential Transcriptional Regulators for F3GT1
3.4. bHLH291 Positively Regulates F3GT1 Promoter Activity
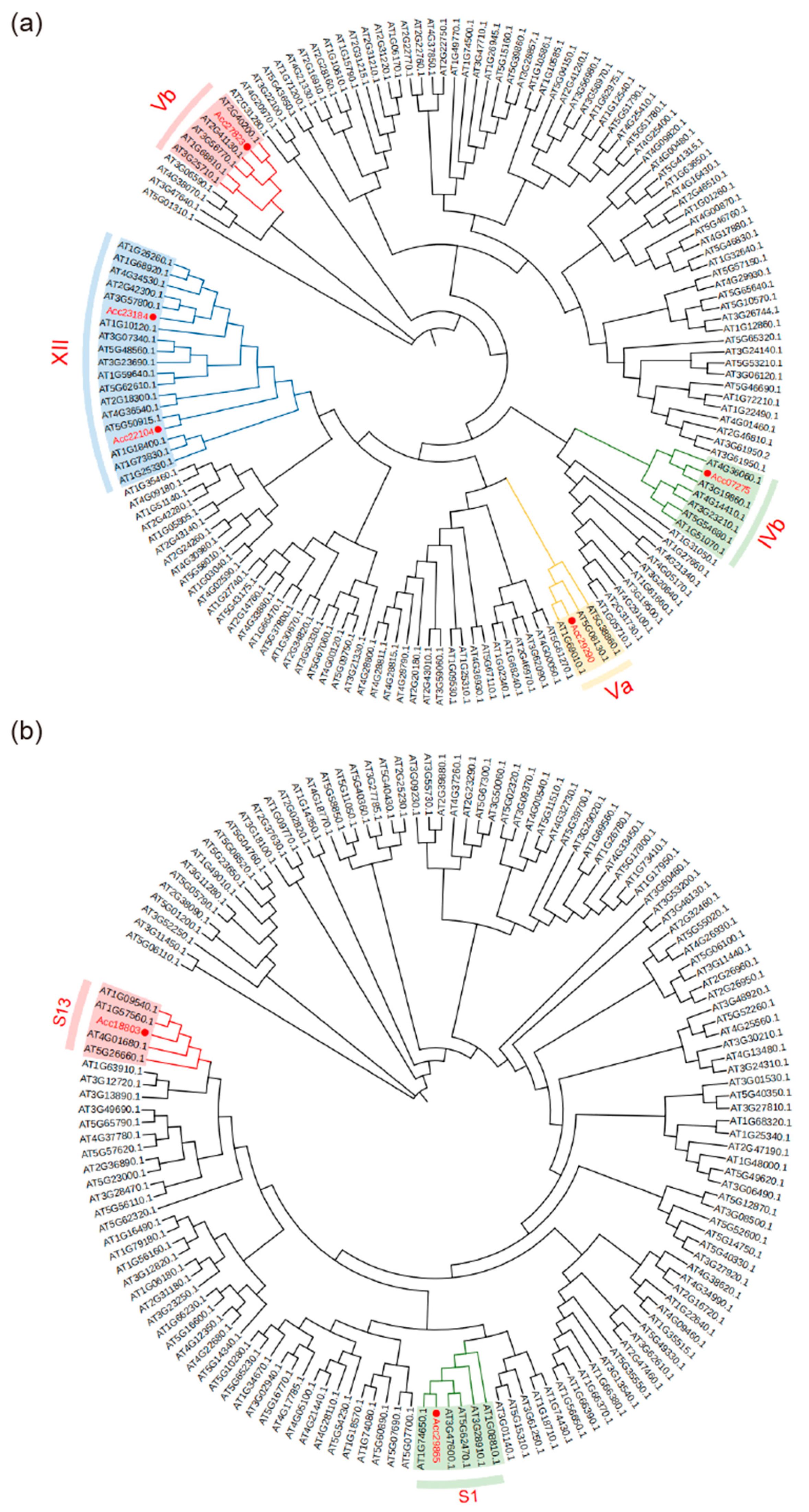
3.5. Phylogenetic Tree of bHLH and MYB Transcription Factor
4. Discussion
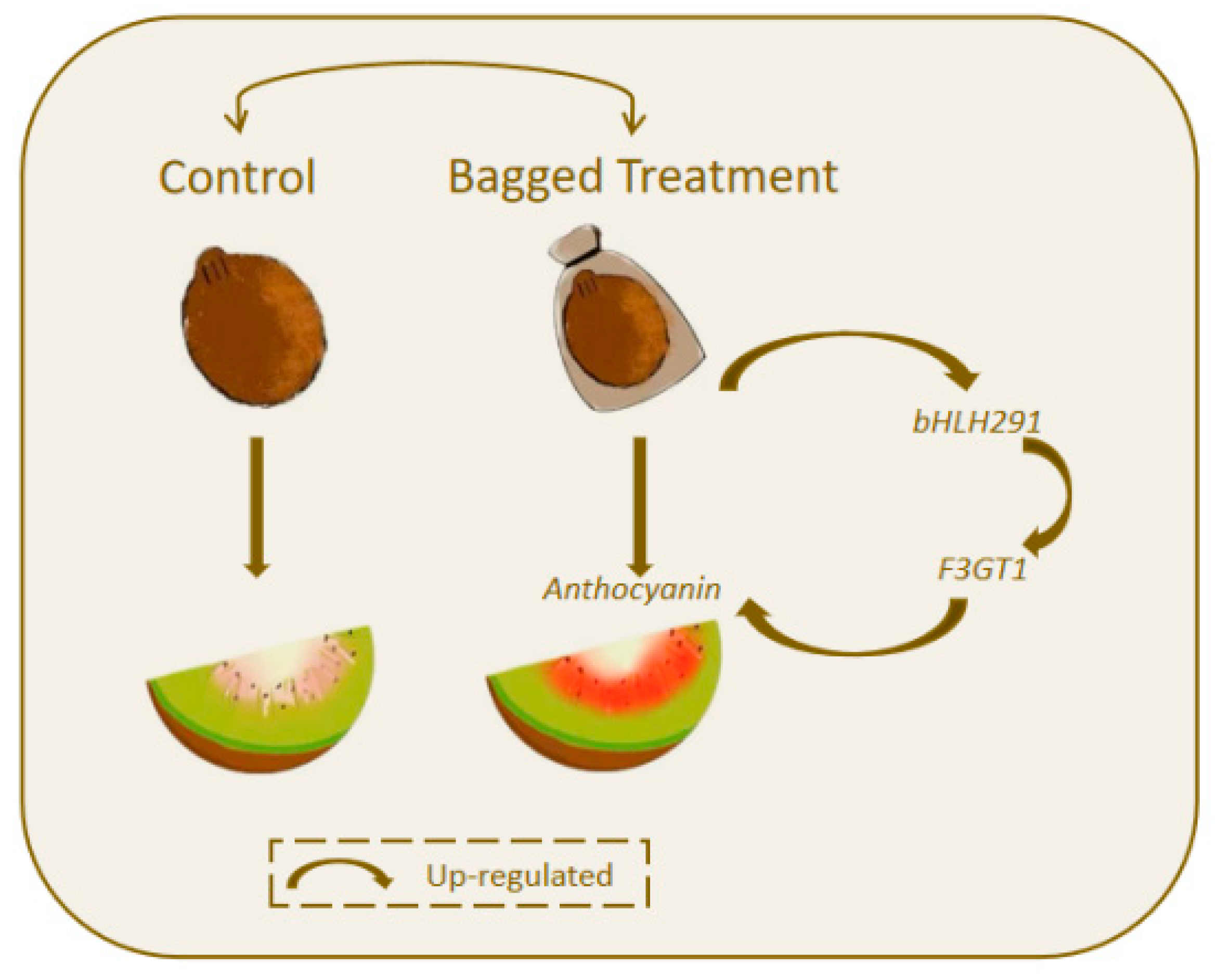
4.1. Bagging Treatment Increases Anthocyanin Content in Kiwifruit Flesh
4.2. F3GT1 Is a Candidate Structural Gene Responsive to Anthocyanin Accumulation Under Bagging Treatment
4.3. bHLH291 Targets F3GT1 and Enhances Its Expression
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Purpose | Primer Sequence 5′-3′ |
|---|---|---|
| F3GT1(Acc20131)-FP | RT-qPCR | TAGCCAAGCAGAGATCCGCTTCC |
| F3GT1(Acc20131)-RP | RT-qPCR | CAAGAATCCTTCTGGTAAGTACTGTTTCGA |
| Actin-FP | RT-qPCR | TGTCCCATGTCTGGTTGATGACT |
| Actin-RP | RT-qPCR | TGCATGAGCGATCAAGTTTCAAG |
| Purpose | Gene | Primer Sequence 5′-3′ |
|---|---|---|
| pSAK277 | bHLH82 (Acc07275)-FP | ACTAGTGGATCCAAAGAATTCATGGATCACCTAAATCACGACGC |
| pSAK277 | bHLH82 (Acc07275)-RP | GACTCTAGAAGTACTCTCGAGTCAATCATCAGCCTTCCCACC |
| pSAK277 | MYB61 (Acc18803)-FP | ACTAGTGGATCCAAAGAATTCATGGGGAGGCACTCTTGCTGC |
| pSAK277 | MYB61 (Acc18803)-RP | GACTCTAGAAGTACTCTCGAGCTAAGAATAGTGTCCAAAAGTGGC |
| pSAK277 | bHLH287 (Acc22104)-FP | ACTAGTGGATCCAAAGAATTCATGGCAGCTTTCCCAAGCACCC |
| pSAK277 | bHLH287 (Acc22104)-RP | GACTCTAGAAGTACTCTCGAGTTAATGGAATGAACACAAGTTG |
| pSAK277 | bHLH291 (Acc23184)-FP | ACTAGTGGATCCAAAGAATTCATGGAGCCAATTTCAGGAACAG |
| pSAK277 | bHLH291 (Acc23184)-RP | GACTCTAGAAGTACTCTCGAGTTAGCTAATAAAGCTGGGATTG |
| pSAK277 | bHLH141 (Acc27823)-FP | ACTAGTGGATCCAAAGAATTCATGGCATGTAAATCAGATCTTG |
| pSAK277 | bHLH141 (Acc27823)-RP | GACTCTAGAAGTACTCTCGAGCTAATGAGAAGGCAAGAAGAACC |
| pSAK277 | bHLH162 (Acc29290)-FP | ACTAGTGGATCCAAAGAATTCATGGAGCTTCCTCAACCCCG |
| pSAK277 | bHLH162 (Acc29290)-RP | GACTCTAGAAGTACTCTCGAGTTAGCTAGTCTTGAGCCTCTTTAGGG |
| pSAK277 | MYB223 (Acc29865)-FP | ACTAGTGGATCCAAAGAATTCATGGGAAGACCACCGTGCTGCG |
| pSAK277 | MYB223 (Acc29865)-RP | GACTCTAGAAGTACTCTCGAGTCAAAACAGATCATCACTTCCCCC |
| pSAK277 | MYBR123 (Acc31791)-FP | ACTAGTGGATCCAAAGAATTCATGGAGATTCTCTCACCGGCG |
| pSAK277 | MYBR123 (Acc31791)-RP | GACTCTAGAAGTACTCTCGAGTCAAAAACTTCTCCATCGATCACCGG |
| LUC | F3GT1 (Acc20131)-FP | GTCGACGGTATCGATAAGCTTGACCGACTTAGATCAACAACTGATCT |
| LUC | F3GT1 (Acc20131)-RP | TGTTTTTGGCGTCTTCCATGGTTTTTTAAGGCTTTGATTTTGG |
Appendix B

References
- Rahim, M.A.; Busatto, N.; Trainotti, L. Regulation of anthocyanin biosynthesis in peach fruits. Planta 2014, 240, 913–929. [Google Scholar] [CrossRef]
- Lee, H.J.; Oh, S.K.; Choi, H.C.; Kim, S.U. Identification of anthocyanins from pigmented rice seeds. Agric. Chem. Biotechnol. 1998, 41, 257–260. [Google Scholar]
- Li, B.Q.; Xia, Y.X.; Wang, Y.Y.; Qin, G.Z.; Tian, S.P. Characterization of Genes Encoding Key Enzymes Involved in Anthocyanin Metabolism of Kiwifruit during Storage Period. Front. Plant Sci. 2017, 8, 8. [Google Scholar] [CrossRef]
- Albert, N.W.; Davies, K.M.; Lewis, D.H.; Zhang, H.B.; Montefiori, M.; Brendolise, C.; Boase, M.R.; Ngo, H.; Jameson, P.E.; Schwinn, K.E. A Conserved Network of Transcriptional Activators and Repressors Regulates Anthocyanin Pigmentation in Eudicots. Plant Cell 2014, 26, 962–980. [Google Scholar] [CrossRef]
- Sytar, O.; Kosyan, A.; Taran, N.; Smetanska, I. Anthocyanin’s as marker for selection of buckwheat plants with high rutin content. Gesunde Pflanz. 2014, 66, 165–169. [Google Scholar] [CrossRef]
- Wang, W.Q.; Moss, S.M.A.; Zeng, L.H.; Espley, R.; Wang, T.C.; Kui, L.W.; Fu, B.L.; Schwinn, K.E.; Allan, A.C.; Yin, X.R. The red flesh of kiwifruit is differentially controlled by specific activation-repression systems. New Phytol. 2022, 235, 630–645. [Google Scholar] [CrossRef]
- Tsuda, T.; Ueno, Y.; Aoki, H.; Koda, T.; Horio, F.; Takahashi, N.; Kawada, T.; Osawa, T. Anthocyanin enhances adipocytokine secretion and adipocyte-specific gene expression in isolated rat adipocytes. Biochem. Biophys. Res. Commun. 2004, 316, 149–157. [Google Scholar] [CrossRef]
- Maierdan, Y.S.; Nulibiya, M.H.T.; Zhou, W.T. Research progress of anthocyanins against cardiovascular diseases. J. Food Saf. Qualit. 2020, 11, 8844–8848. [Google Scholar]
- Navas, M.J.; Jiménez-Moreno, A.M.; Bueno, J.M.; Sáez-Plaza, P.; Asuero, A.G. Analysis and Antioxidant Capacity of Anthocyanin Pigments. Part IV: Extraction of Anthocyanins. Crit. Rev. Anal. Chem. 2012, 42, 313–342. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, J.L.; Cherono, S.; An, J.P.; Allan, A.C.; Han, Y.P. Colorful hues: Insight into the mechanisms of anthocyanin pigmentation in fruit. Plant Physiol. 2023, 192, 1718–1732. [Google Scholar] [CrossRef]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar] [CrossRef]
- He, J.P.; Yao, L.; Pecoraro, L.; Liu, C.X.; Wang, J.; Huang, L.Q.; Gao, W.Y. Cold stress regulates accumulation of flavonoids and terpenoids in plants by phytohormone, transcription process, functional enzyme, and epigenetics. Crit. Rev. Biotechnol. 2023, 43, 680–697. [Google Scholar] [CrossRef]
- Naing, A.H.; Kim, C.K. Abiotic stress-induced anthocyanins in plants: Their role in tolerance to abiotic stresses. Physiol. Plant. 2021, 172, 1711–1723. [Google Scholar] [CrossRef]
- Yu, M.; Man, Y.P.; Wang, Y.C. Light- and Temperature-Induced Expression of an R2R3-MYB Gene Regulates Anthocyanin Biosynthesis in Red-Fleshed Kiwifruit. Int. J. Mol. Sci. 2019, 20, 5228. [Google Scholar] [CrossRef]
- Ngoc, A.T.; Naing, A.H.; Kim, C.K. Influences of different light sources and light/dark cycles on anthocyanin accumulation and plant growth in Petunia. J. Plant Biotechnol. 2016, 43, 119–124. [Google Scholar]
- Wang, N.; Zhang, Z.Y.; Jiang, S.H.; Xu, H.F.; Wang, Y.C.; Feng, S.Q.; Chen, X.S. Synergistic effects of light and temperature on anthocyanin biosynthesis in callus cultures of red-fleshed apple (Malus sieversii f. niedzwetzkyana). Plant Cell Tissue Organ Cult. 2016, 127, 217–227. [Google Scholar] [CrossRef]
- Petrella, D.P.; Metzger, J.D.; Blakeslee, J.J.; Nangle, E.J.; Gardner, D.S. Anthocyanin Production Using Rough Bluegrass Treated with High-Intensity Light. HortScience 2016, 51, 1111–1120. [Google Scholar] [CrossRef]
- Cominelli, E.; Gusmaroli, G.; Allegra, D.; Galbiati, M.; Wade, H.K.; Jenkins, G.I.; Tonelli, C. Expression analysis of anthocyanin regulatory genes in response to different light qualities in Arabidopsis thaliana. J. Plant Physiol. 2008, 165, 886–894. [Google Scholar] [CrossRef]
- Wang, J.D.; Zhao, Y.Q.; Sun, B.; Yang, Y.T.; Wang, S.P.; Feng, Z.R.; Li, J.Y. The structure of anthocyanins and the copigmentation by common micromolecular copigments: A review. Food Res. Int. 2024, 176, 15. [Google Scholar] [CrossRef]
- Zhao, Z.C.; Hu, G.B.; Hu, F.C.; Wang, H.C.; Yang, Z.Y.; Lai, B. The UDP glucose: Flavonoid-3-O-glucosyltransferase (UFGT) gene regulates anthocyanin biosynthesis in litchi (Litchi chinesis Sonn.) during fruit coloration. Mol. Biol. Rep. 2012, 39, 6409–6415. [Google Scholar] [CrossRef]
- Li, X.J.; Zhang, J.Q.; Wu, Z.C.; Lai, B.; Huang, X.M.; Qin, Y.H.; Wang, H.C. Hu Functional characterization of a glucosyltransferase gene, LcUFGT1, involved in the formation of cyanidin glucoside in the pericarp of Litchi chinensis. Physiol. Plant. 2016, 156, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Ma, K.X.; Qi, Y.W.; Lv, G.W.; Ren, X.L.; Liu, Z.D.; Ma, F.W. Transcriptional Regulation of Anthocyanin Synthesis by MYB-bHLH-WDR Complexes in Kiwifruit (Actinidia chinensis). J. Agric. Food Chem. 2021, 69, 3677–3691. [Google Scholar] [CrossRef]
- Wang, W.; Yang, C.; Jiang, Y.B.; Tang, G.C.; Guo, L.Y.; Qiao, G.Z.; Liu, S.H.; Tan, B.; Cheng, J.; Zhang, L.L.; et al. The basic helix-loop-helix transcription factor PpeUNE12 regulates peach ripening by promoting polyamine catabolism and anthocyanin synthesis. Plant Physiol. Biochem. 2025, 220, 109537. [Google Scholar] [CrossRef] [PubMed]
- Ning, G.X.; Li, W.F.; Chu, M.Y.; Ma, Z.H.; Wang, P.; Mao, J.; Chen, B.H. MdbHLH51 plays a positive role in anthocyanin accumulation in ‘Red Delicious’ apples. Trees 2022, 36, 1687–1695. [Google Scholar] [CrossRef]
- Hao, Y.Q.; Zong, X.M.; Ren, P.; Qian, Y.Q.; Fu, A.G. Basic Helix-Loop-Helix (bHLH) Transcription Factors Regulate a Wide Range of Functions in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 7152. [Google Scholar] [CrossRef]
- Wang, N.; Qu, C.Z.; Jiang, S.H.; Chen, Z.J.; Xu, H.F.; Fang, H.C.; Su, M.Y.; Zhang, J.; Wang, Y.C.; Liu, W.J.; et al. The proanthocyanidin-specific transcription factor MdMYBPA1 initiates anthocyanin synthesis under low-temperature conditions in red-fleshed apples. Plant J. 2018, 96, 39–55. [Google Scholar] [CrossRef]
- An, J.P.; Wang, X.F.; Zhang, X.W.; Xu, H.F.; Bi, S.Q.; You, C.X.; Hao, Y.J. An apple MYB transcription factor regulates cold tolerance and anthocyanin accumulation and undergoes MIEL1-mediated degradation. Plant Biotechnol. J. 2020, 18, 337–353. [Google Scholar] [CrossRef]
- Xiong, Y.; He, J.; Li, M.; Du, K.; Lang, H.; Gao, P.; Xie, Y. Integrative Analysis of Metabolome and Transcriptome Reveals the Mechanism of Color Formation in Yellow-Fleshed Kiwifruit. Int. J. Mol. Sci. 2023, 24, 1573. [Google Scholar] [CrossRef]
- Wang, W.Q.; Liu, X.F.; Zhu, Y.J.; Zhu, J.Z.; Liu, C.; Wang, Z.Y.; Shen, X.X.; Allan, A.C.; Yin, X.R. Identification of miRNA858 long-loop precursors in seed plants. Plant Cell 2024, 36, 1637–1654. [Google Scholar] [CrossRef]
- Pilkington, S.M.; Crowhurst, R.; Hilario, E.; Nardozza, S.; Fraser, L.; Peng, Y.Y.; Gunaseelan, K.; Simpson, R.; Tahir, J.; Deroles, S.C.; et al. A manually annotated Actinidia chinensis var. chinensis (kiwifruit) genome highlights the challenges associated with draft genomes and gene prediction in plants. BMC Genom. 2018, 19, 257. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Ferradás, Y.; Rey, L.; Martínez, Ó.; Rey, M.; González, M.V. Identification and validation of reference genes for accurate normalization of real-time quantitative PCR data in kiwifruit. Plant Physiol. Biochem. 2016, 102, 27–36. [Google Scholar] [CrossRef]
- Kamo, K.; Blowers, A.; McElroy, D. Effect of the cauliflower mosaic virus 35S, actin, and ubiquitin promoters on uidA expression from a bar-uidA-fusion gene in transgenic Gladiolus plants. In Vitro Cell. Dev. Biol.-Plant 2000, 36, 13–20. [Google Scholar] [CrossRef]
- Hellens, R.P.; Allan, A.C.; Friel, E.N.; Bolitho, K.; Grafton, K.; Templeton, M.D.; Karunairetnam, S.; Gleave, A.P.; Laing, W.A. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods 2005, 1, 14. [Google Scholar] [CrossRef]
- Fu, B.L.; Wang, W.Q.; Liu, X.F.; Duan, X.W.; Allan, A.C.; Grierson, D.; Yin, X.R. An ethylene-hypersensitive methionine sulfoxide reductase regulated by NAC transcription factors increases methionine pool size and ethylene production during kiwifruit ripening. New Phytol. 2021, 232, 237–251. [Google Scholar] [CrossRef]
- Montefiori, M.; Espley, R.V.; Stevenson, D.; Cooney, J.; Datson, P.M.; Saiz, A.; Atkinson, R.G.; Hellens, R.P.; Allan, A.C. Identification and characterisation of F3GT1 and F3GGT1, two glycosyltransferases responsible for anthocyanin biosynthesis in red-fleshed kiwifruit (Actinidia chinensis). Plant J. 2011, 65, 106–118. [Google Scholar] [CrossRef]
- Nardozza, S.; Boldingh, H.L.; Kashuba, M.P.; Feil, R.; Jones, D.; Thrimawithana, A.H.; Ireland, H.S.; Philippe, M.; Wohlers, M.W.; McGhie, T.K.; et al. Carbon starvation reduces carbohydrate and anthocyanin accumulation in red-fleshed fruit via trehalose 6-phosphate and MYB27. Plant Cell Environ. 2020, 43, 819–835. [Google Scholar] [CrossRef]
- Wang, L.; Tang, W.; Hu, Y.; Zhang, Y.; Sun, J.; Guo, X.; Lu, H.; Yang, Y.; Fang, C.; Niu, X.; et al. A MYB/bHLH complex regulates tissue-specific anthocyanin biosynthesis in the inner pericarp of red-centered kiwifruit Actinidia chinensis cv. Hongyang. Plant J. 2019, 99, 359–378. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Sun, L.X.; Xi, D.D.; Li, X.F.; Gao, L.; Miao, L.M.; Luo, Y.; Tian, M.M.; Zhu, H.F. Methyl jasmonate-induced bHLH42 mediates tissue-specific accumulation of anthocyanins via regulating flavonoid metabolism-related pathways in Caitai. Physiol. Plant. 2024, 176, 13. [Google Scholar] [CrossRef]
- Castillejo, C.; Waurich, V.; Wagner, H.; Ramos, R.; Oiza, N.; Muñoz, P.; Triviño, J.C.; Caruana, J.; Liu, Z.C.; Cobo, N.; et al. Allelic Variation of MYB10 Is the Major Force Controlling Natural Variation in Skin and Flesh Color in Strawberry (Fragaria spp.) Fruit. Plant Cell 2020, 32, 3723–3749. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.Y.; Kui, L.W.; Cooney, J.M.; Wang, T.; Espley, R.V.; Allan, A.C. Differential regulation of the anthocyanin profile in purple kiwifruit (Actinidia species). Hortic. Res. 2019, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Pu, R.; Wu, Y.W.; Bai, T.; Li, Y.; Li, X.J.; Li, N.B.; Zhou, Y.; Zhang, J.L. Molecular and Metabolic Regulation of Flavonoid Biosynthesis in Two Varieties of Dendrobium devonianum. Curr. Issues Mol. Biol. 2024, 46, 14270–14290. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Dubos, C. The arabidopsis bHLH transcription factor family. Trends Plant Sci. 2024, 29, 13. [Google Scholar] [CrossRef]
- Baudry, A.; Caboche, M.; Lepiniec, L. TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell-specific accumulation of flavonoids in Arabidopsis thaliana. Plant J. 2006, 46, 768–779. [Google Scholar] [CrossRef]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef]
- Wang, X.C.; Wu, J.; Guan, M.L.; Zhao, C.H.; Geng, P.; Zhao, Q. Arabidopsis MYB4 plays dual roles in flavonoid biosynthesis. Plant J. 2020, 101, 637–652. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Zhou, H.; He, J.; Zhang, Y.; Zhao, H.; Sun, X.; Chen, X.; Liu, X.; Zheng, Y.; Lin, H. RHA2b-mediated MYB30 degradation facilitates MYB75-regulated, sucrose-induced anthocyanin biosynthesis in Arabidopsis seedlings. Plant Commun. 2024, 5, 100744. [Google Scholar] [CrossRef]
- Liu, Y.; Liang, Q.; Tang, D.B.; Chen, Y.Q.; Zang, J.W.; Zhao, W.J.; Chen, J.G.; Zhang, Q.F.; Yin, Z.P. Development of suspension culture technology and hormone effects on anthocyanin biosynthesis for red Cyclocarya paliurus cells. Plant Cell Tissue Organ Cult. 2022, 149, 175–195. [Google Scholar] [CrossRef]
- Meng, X.C.; Wang, X.J. Regulation of flower development and anthocyanin accumulation in Gerbera hybrida. J. Horticult. Sci. Biotechnol. 2004, 79, 131–137. [Google Scholar] [CrossRef]
- An, X.; Tan, T.; Zhang, X.; Guo, X.; Zhu, Y.; Song, Z.; Wang, D. Effects of Light Intensity on Endogenous Hormones and Key Enzyme Activities of Anthocyanin Synthesis in Blueberry Leaves. Horticulturae 2023, 9, 618. [Google Scholar] [CrossRef]
- Ju, Z.G. Fruit bagging, a useful method for studying anthocyanin synthesis and gene expression in apples. Sci. Hortic. 1998, 77, 155–164. [Google Scholar] [CrossRef]
- Zhou, S.H.; Guo, R.R.; Wei, R.F.; Liu, J.B.; Yu, H.; Shi, X.F.; Zhang, Y.; Xie, T.L.; Cheng, G. Effects of bagging or the combination of umbrella and bag treatments on anthocyanin accumulation in the berry skin of ‘Kyoho’ (Petunia Vitis labruscana) Grape. Food Sci. Technol. 2020, 40, 394–400. [Google Scholar] [CrossRef]
- Pazmiño-Durán, E.A.; Giusti, M.M.; Wrolstad, R.E.; Glória, M.B.A. Anthocyanins from Oxalis triangularis as potential food colorants. Food Chem. 2001, 75, 211–216. [Google Scholar] [CrossRef]
- Kotepong, P.; Ketsa, S.; van Doorn, W.G. A white mutant of Malay apple fruit (Syzygium malaccense) lacks transcript expression and activity for the last enzyme of anthocyanin synthesis, and the normal expression of a MYB transcription factor. Funct. Plant Biol. 2011, 38, 75–86. [Google Scholar] [CrossRef]
- Kovinich, N.; Saleem, A.; Arnason, J.T.; Miki, B. Functional characterization of a UDP-glucose:flavonoid 3-O-glucosyltransferase from the seed coat of black soybean (Glycine max (L.) Merr.). Phytochemistry 2010, 71, 1253–1263. [Google Scholar] [CrossRef]
- An, X.L.; Tan, T.Y.; Song, Z.J.; Guo, X.L.; Zhang, X.Y.; Zhu, Y.Z.; Wang, D.L. Physiological response of anthocyanin synthesis to different light intensities in blueberry. PLoS ONE 2023, 18, e0283284. [Google Scholar] [CrossRef]
- Wang, F.H.; Ahammed, G.J.; Li, G.Y.; Bai, P.T.; Jiang, Y.; Wang, S.X.; Chen, S.C. Ethylene is involved in red light-induced anthocyanin biosynthesis in cabbage (Brassica oleracea). Int. J. Agric. Biol. 2019, 21, 955–963. [Google Scholar]
- Fraser, L.G.; Seal, A.G.; Montefiori, M.; McGhie, T.K.; Tsang, G.K.; Datson, P.M.; Hilario, E.; Marsh, H.E.; Dunn, J.K.; Hellens, R.P.; et al. An R2R3 MYB transcription factor determines red petal colour in an Actinidia (kiwifruit) hybrid population. BMC Genom. 2013, 14, 28. [Google Scholar] [CrossRef]
- Li, Y.; Xu, P.B.; Chen, G.Q.; Wu, J.; Liu, Z.C.; Lian, H.L. FvhHLH9 Functions as a positive regulator of anthocyanin biosynthesis by forming a HY5-bHLH9 Transcription Complex in Strawberry Fruits. Plant Cell Physiol. 2020, 61, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.B.; Li, S.; Zhang, R.F.; Zhao, J.; Chen, Y.C.; Zhao, Q.; Yao, Y.X.; You, C.X.; Zhang, X.S.; Hao, Y.J. The bHLH transcription factor MdbHLH3 promotes anthocyanin accumulation and fruit colouration in response to low temperature in apples. Plant Cell Environ. 2012, 35, 1884–1897. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Zhou, L.; Han, L.L.; Zou, H.Z.; Miao, K.; Wang, Y. PsbHLH1, a novel transcription factor involved in regulating anthocyanin biosynthesis in tree peony (Paeonia suffruticosa). Plant Physiol. Biochem. 2020, 154, 396–408. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, X.-Y.; Zhang, K.-Y.; Tu, M.-Y.; Yin, X.-R.; Wang, W.-Q. bHLH291 Regulates Anthocyanin Accumulation in Kiwifruit Under Bagging Treatment. Horticulturae 2025, 11, 666. https://doi.org/10.3390/horticulturae11060666
Kang X-Y, Zhang K-Y, Tu M-Y, Yin X-R, Wang W-Q. bHLH291 Regulates Anthocyanin Accumulation in Kiwifruit Under Bagging Treatment. Horticulturae. 2025; 11(6):666. https://doi.org/10.3390/horticulturae11060666
Chicago/Turabian StyleKang, Xin-Yun, Ke-Yu Zhang, Mei-Yan Tu, Xue-Ren Yin, and Wen-Qiu Wang. 2025. "bHLH291 Regulates Anthocyanin Accumulation in Kiwifruit Under Bagging Treatment" Horticulturae 11, no. 6: 666. https://doi.org/10.3390/horticulturae11060666
APA StyleKang, X.-Y., Zhang, K.-Y., Tu, M.-Y., Yin, X.-R., & Wang, W.-Q. (2025). bHLH291 Regulates Anthocyanin Accumulation in Kiwifruit Under Bagging Treatment. Horticulturae, 11(6), 666. https://doi.org/10.3390/horticulturae11060666