
Enhancement of Phenolic and Polyacetylene Accumulation in Lobelia chinensis (Chinese lobelia) Plantlet Cultures Through Yeast Extract and Salicylic Acid Elicitation
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Culture Conditions
2.2. Elicitation
2.3. Biomass Estimation
2.4. Preparation of Plant Extract
2.5. Estimation of Total Phenolic Content
2.6. Estimation of Total Flavonoid Content
2.7. Quantification of Phenolic Compounds
2.8. Quantification of Polyacetylenes
2.9. Analysis of Antioxidant Activities
2.9.1. 2,2 Diphenyl 1 Picrylhydrazyl (DPPH) Radical Scavenging Assay
2.9.2. 2,2′-Azino-bis (3-ethybenzothiazoline-6-sulphonic acid) (ABTS) Assay
2.9.3. Ferric Reducing Antioxidant Power (FRAP) Assay
2.10. Statistical Analysis
3. Results
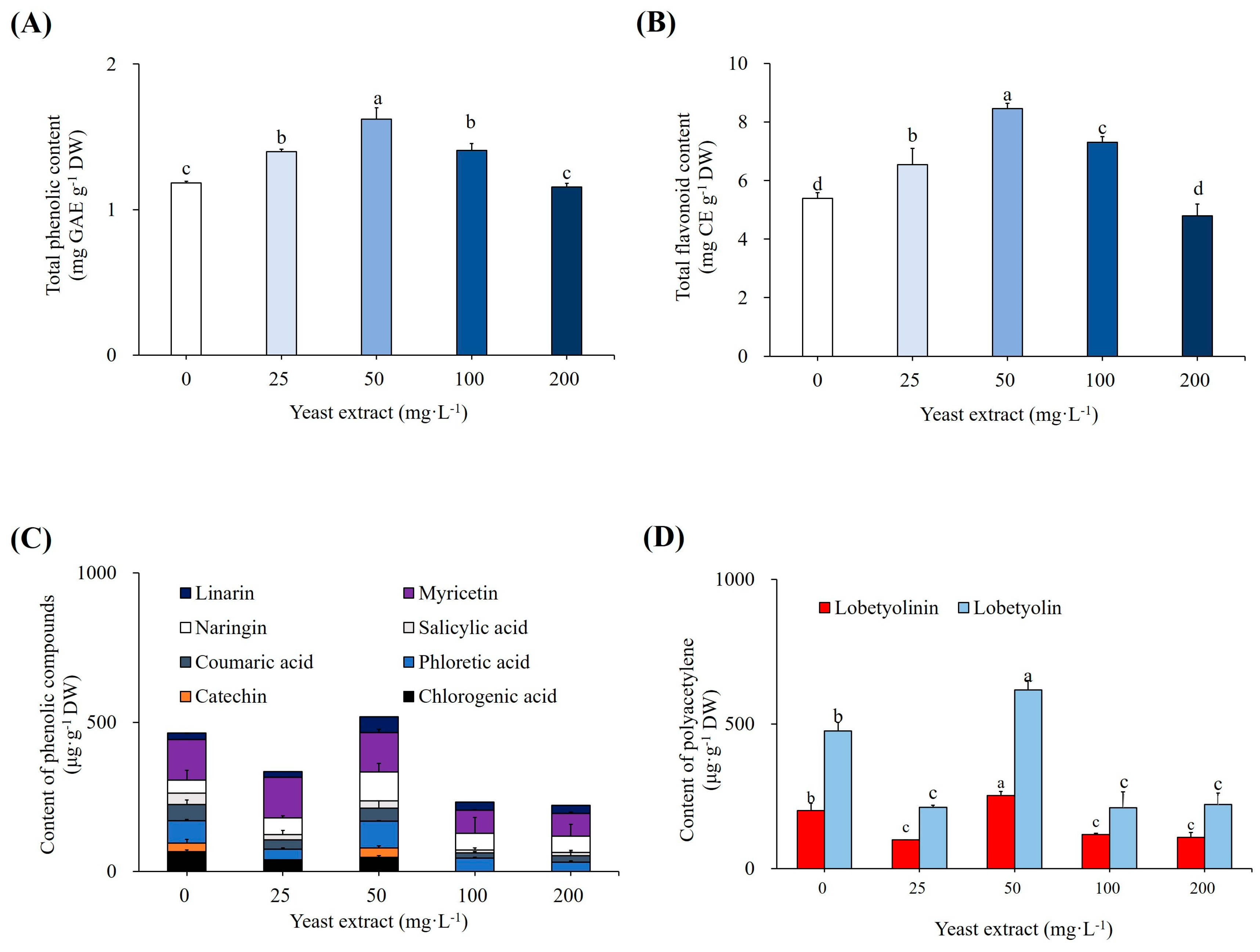
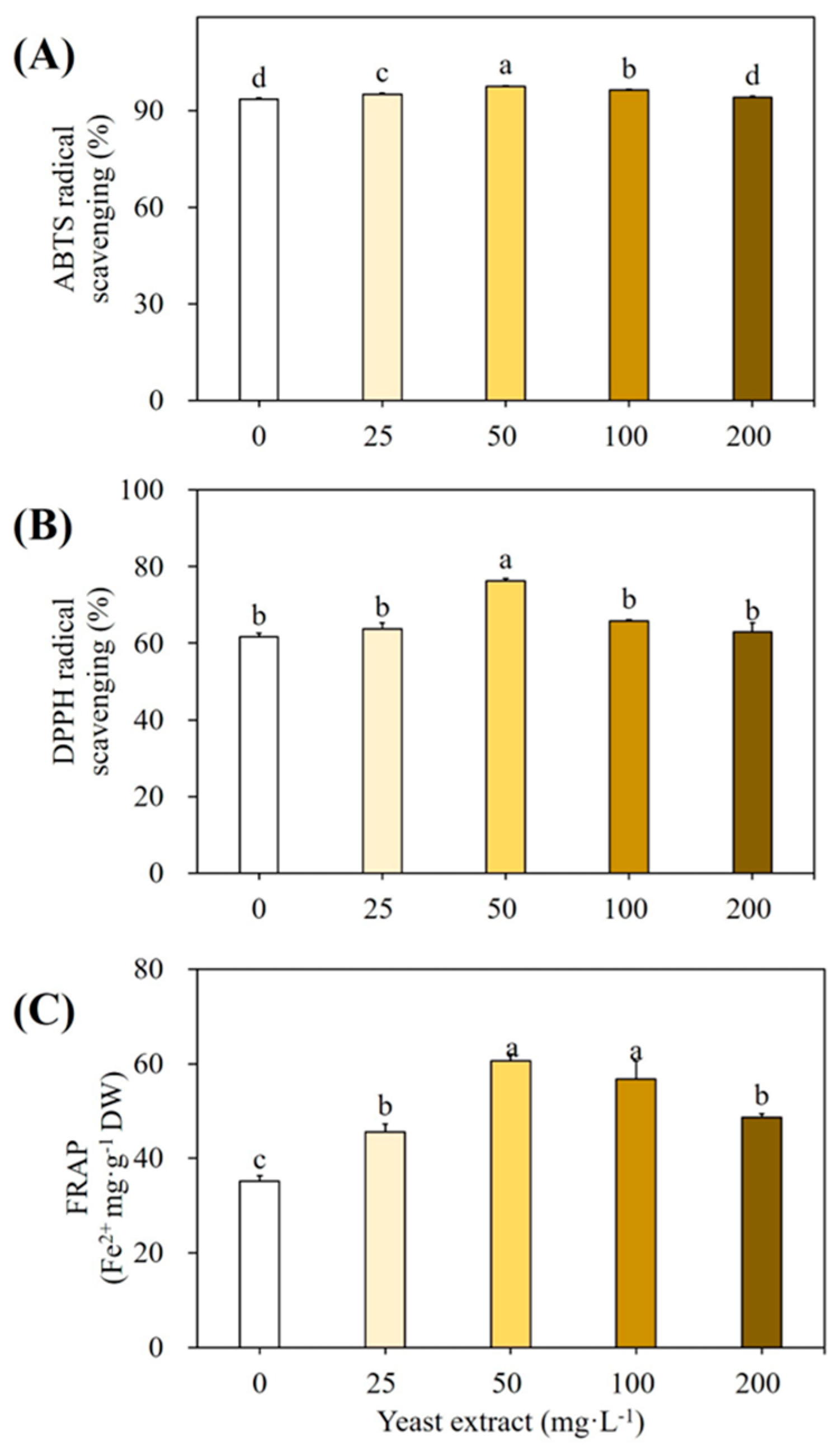
3.1. Elicitation of Cultures with Yeast Extract
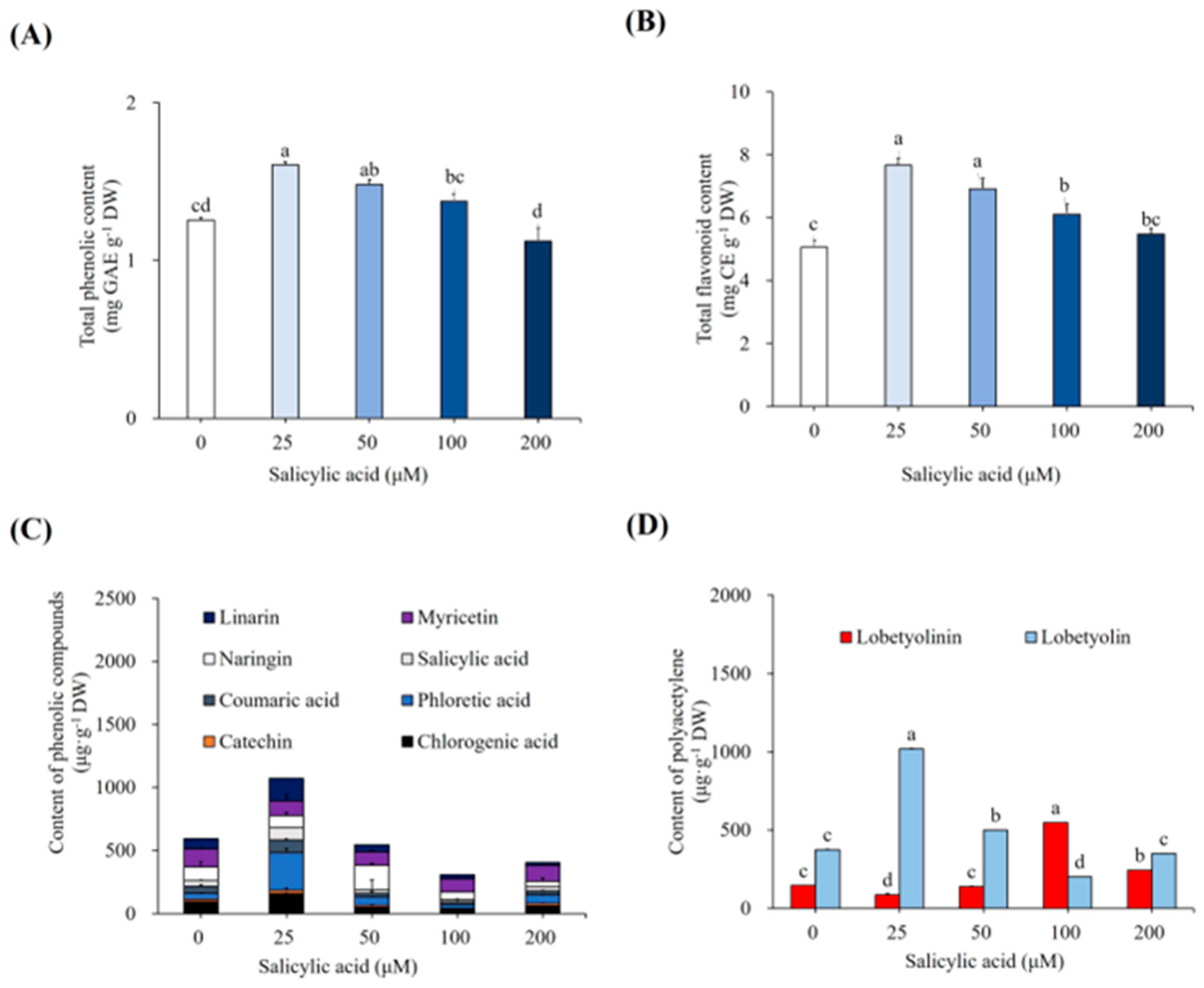
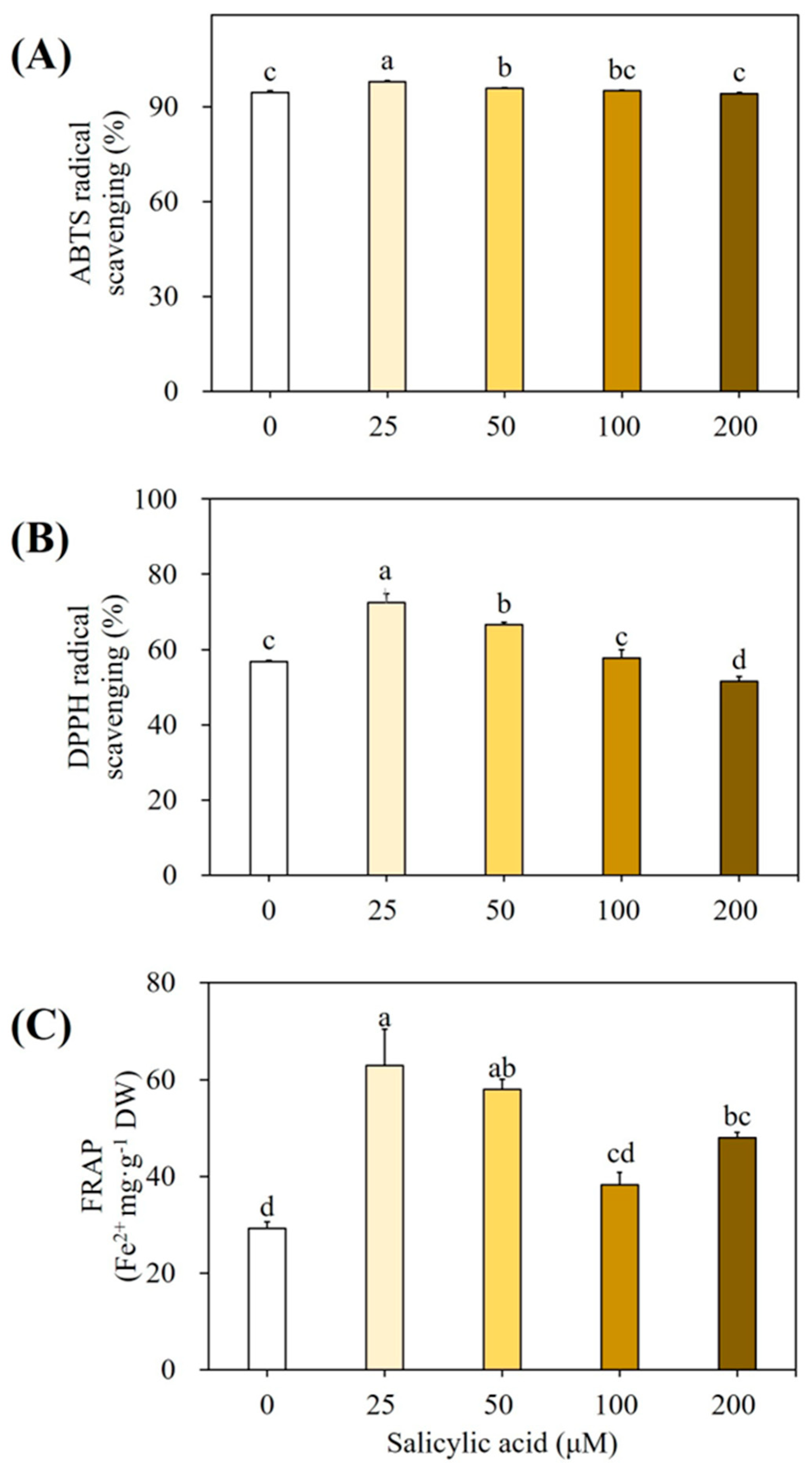
3.2. Elicitation of Cultures with Salicylic Acid
4. Discussion
4.1. Effect of YE on Growth of Plantlets
4.2. Effect of YE on Accumulation of SMs and Antioxidant Enzymes
4.3. Effect of SA on Growth of Plantlets
4.4. Effect of SA on Accumulation of SMs and Antioxidant Enzymes
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, S. The Ben Cao Gang Mu (Chinese Edition), 1st ed.; University of California Press: Oakland, CA, USA, 2016; p. 265. [Google Scholar]
- Falquitto, D.G.; Swiech, J.N.D.; Pereira, C.B.; Bobek, V.B.; Possagno, G.C.H.; Farago, P.V.; Miguel, M.D.; Durate, J.L.; Miguel, O.G. Biological activity, phytochemistry and traditional uses of genus Lobelia (Campanulaceae): A systematic review. Fitoterapia 2019, 134, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Li, K.C.; Ho, Y.L.; Huang, G.J.; Chang, Y.S. Anti-oxidative and anti-inflammatory effects of Lobelia chinensis in vitro and in vivo. Am. J. Chin. Med. 2015, 43, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Reddy, N.; Khoo, C.; Koyyalamudi, S.R.; Jone, C.E. Antioxidant and immunomodulatory activities and structural characterization of polysaccharides isolated from Lobelia chinensis Lour. Pharmacologia 2018, 9, 157–168. [Google Scholar] [CrossRef]
- Kuo, P.C.; Hwang, T.L.; Lin, Y.T.; Kuo, Y.C.; Leu, Y.L. Chemical constituents from Lobelia chinensis and their anti-virus and anti-inflammatory bioactivates. Arch. Pham. Res. 2011, 34, 715–722. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, P.; Zhang, X.; Li, X. Chemical structure elucidation of an inulin-type fructan isolated from Lobelia chinensis Lour. with anti-obesity on diet-induced mice. Carbohyr. Polym. 2020, 240, 116357. [Google Scholar] [CrossRef]
- Choi, W.H.; Lee, I.A. The anti-tubercular activity of Melia azaedarach L. and Lobelia chinensis Lour. and their potential as effect anti-Mycobacterium tuberculosis candidate agents. Asian Pac. J. Trop. Biomed. 2016, 6, 830–835. [Google Scholar] [CrossRef]
- Chen, M.S.; Chen, W.R.; Zhang, J.M.; Long, X.Y.; Wang, Y.T. Lobelia chinensis: Chemical constituents and anticancer activity perspective. Chin. J. Nat. Med. 2014, 12, 103–107. [Google Scholar] [CrossRef]
- Bai, X.; Lee, H.S.; Han, E.J.; Murthy, H.N.; Kwon, H.J.; Yeon, S.H.; Park, S.Y. Stimulating the synthesis of phenolics and polyacetylenes in Lobelia chinensis Lour. Plantlets using various bioreactor culture systems: A comparison of parameter effects. Plant Cell Tissue Organ Cult. 2025, 16, 36. [Google Scholar] [CrossRef]
- Murthy, H.N.; Joseph, K.S.; Hahn, J.E.; Lee, H.S.; Paek, K.Y.; Park, S.Y. Suspension culture of somatic embryos for the production of high-value secondary metabolites. Physiol. Mol. Biol. Plants 2023, 29, 1153–1177. [Google Scholar] [CrossRef]
- Murthy, H.N.; Lee, E.J.; Paek, K.Y. Production of secondary metabolites from cell and organ cultures: Strategies and approaches for biomass improvement and metabolite accumulation. Plant Cell Tissue Organ Cult. 2014, 118, 1–16. [Google Scholar] [CrossRef]
- Ramirez-Estrda, K.; Limon, H.V.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cudio, R.M.; Palazon, J. Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef] [PubMed]
- Halder, M.; Sarkar, S.; Jha, S. Elicitation: A biotechnological tool for enhanced production of secondary metabolites in hairy root cultures. Eng. Life Sci. 2019, 19, 880–895. [Google Scholar] [CrossRef] [PubMed]
- Pathak, A.R.; Patel, S.R.; Joshi, A.G.; Shrivastava, N.; Sindhav, G.; Sharma, S.; Ansari, H. Elicitor mediated enhancement of shoot biomass and lupeol production in Hemidesmus indicus (L.) R.Br. ex. Schult and Tylophora indica (Burm. F.) Merrill using yeast extract and salicylic acid. Nat. Prod. Res. 2023, 37, 1167–1773. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.Y.; Wang, M.; Wu, X.H.; Fan, M.Z.; Li, H.Z.; Guo, Y.Q.; Jiang, J.; Yin, C.R.; Lian, M.L. Improving flavonoid accumulation of bioreactor-cultured adventitious roots in Oplapanx elatus using yeast extract. Plants 2023, 12, 2174. [Google Scholar] [CrossRef]
- Laezza, C.; Imbimbo, P.; D’Amelia, V.; Marzocchi, A.; Monti, D.M.; di Loria, A.; Monti, S.M.; Novellino, E.; Tenore, G.C.; Rigano, M.M. Use of yeast extract to elicit a pulp-derived callus cultures from Annurca apple and potentiate its biological activity. J. Funct. Foods 2024, 112, 105988. [Google Scholar] [CrossRef]
- Zanella, L.; Gismondi, A.; Di Marco, G.; Barglia, R.; Scuderi, F.; Redi, E.L.; Galgani, A.; Canini, A. Induction of antioxidant metabolites in Moringa oleifera callus by abiotic stresses. J. Nat. Prod. 2019, 82, 23792386. [Google Scholar] [CrossRef]
- Moharramnejad, S.; Azam, A.T.; Panahandeh, J.; Dehghnian, Z.; Ashraf, M. Effect of methyl jasmonate and salicylic acid in vitro growth, sativoside production, and oxidative defense system in Stevia rebaudiana. Sugar Tech. 2019, 21, 1031–1038. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 373–497. [Google Scholar] [CrossRef]
- Harborne, A.J. Phytochemical Methods: A Guide to Modern Techniques of Plant Analysis, 1st ed.; Springer: Dordrecht, The Netherlands, 1994. [Google Scholar]
- Burin, V.M.; Arari, S.G.; Costa, L.L.F.; Bordingon-Luiz, A.M.T. Determination of some phenolic compounds in red wind by RP-HPLC: Method development and validation. J. Chromatogr. Sci. 2011, 49, 647–651. [Google Scholar] [CrossRef]
- Qiao, C.F.; He, Z.D.; Han, Q.B.; Hu, H.X.; Jiang, R.W.; Li, S.L.; Zhang, Y.B.; But, P.P.H.; Shaw, P.C. The use of lobetyolin and HPLC-UV fingerprints for quality assessment of radix codonopsis. J. Food Drug Anal. 2007, 15, 258–264. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical methods used in determining antioxidant activity: A review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Jalota, K.; Sharma, V.; Agarwal, C.; Jindal, S. Eco-friendly approaches to phytochemical production: Elicitation and beyond. Nat. Prod. Bioproct. 2024, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Joseph, K.S.; Paek, K.Y.; Park, S.Y. Light as an elicitor for enhanced production of secondary metabolites in plant cell, tissue, and organ cultures. Plant Growth Regul. 2024, 104, 31–49. [Google Scholar] [CrossRef]
- Murthy, H.N.; Joseph, K.S.; Paek, K.Y.; Park, S.Y. Production of specialized metabolites in plant cell and organo-cultures: The role of gamma radiation in eliciting secondary metabolites. Int. J. Rad. Biol. 2024, 100, 678–688. [Google Scholar] [CrossRef]
- Murthy, H.N.; Joseph, K.S.; Paek, K.Y.; Park, S.Y. Nanomaterials as novel elicitors of pharmacologically active plant specialized metabolites in cell and organ cultures: Current status and future outlooks. Plant Growth Regul. 2024, 104, 5–30. [Google Scholar] [CrossRef]
- Tao, Z.; Yuan, H.; Liu, M.; Liu, Q.; Zhang, S.; Liu, H.; Jiang, Y.; Huang, D.; Wang, T. Yeast extract: Characteristics, production, applications and future perspectives. J. Micbiol. Biotechnol. 2023, 33, 151–166. [Google Scholar] [CrossRef]
- Klotz, S.; Kuenz, A.; Prüße, U. Nutritional requirements and the impact of yeast extract on the D-lactic acid production by Sporolactobacillus inulinus. Green Chem. 2017, 19, 4633–4641. [Google Scholar] [CrossRef]
- Rasouli, D.; Werbrouck, S.; Maleki, B.; Jafary, H.; Schurdi-Levraud, V. Elicitor-induced in vitro shoot multiplication and steviol glycosides production in Stevia rebaudiana. S. Afr. J. Bot. 2021, 137, 265–271. [Google Scholar] [CrossRef]
- Petrova, M.; Geneva, M.; Trendafilova, A.; Miladinova-Georgieva, K.; Dimitrova, L.; Sichanowa, M.; Nikolova, M.; Ivanova, V.; Dimitrova, M.; Sozoniuk, M. Antioxidant capacity and accumulation of caffeoylquinic acids in Arnica montana L. in vitro shoots after elicitation with yeast extract or salicylic acid. Plants 2025, 14, 967. [Google Scholar] [CrossRef]
- Jirakiattikul, Y.; Ruangnoo, S.; Sangmukdee, K.; Chamchusri, K.; Rithichai, P. Enhancement of plumbagin production through elicitation in in vitro-regenerated shoots of Plumbago indica L. Plants 2024, 13, 1450. [Google Scholar] [CrossRef]
- Abraham, F.; Bhatt, A.; Keng, C.L.; Indrayanto, G.; Sulaiman, S.F. Effect of yeast extract and chitosan on shoot proliferation, morphology and antioxidant activity of Curcuma mangga in vitro plantlets. Afr. J. Biotechnol. 2011, 10, 7787–7795. [Google Scholar] [CrossRef]
- Gonçalves, S.; Mansinhos, I.; Rodríguez-Solana, R.; SantínE, P.; Coelho, N.; Romano, A. Elicitation improves rosmarinic acid content and antioxidant activity in Thymus lotocephalus shoot cultures. Ind. Crops Prod. 2019, 137, 214–220. [Google Scholar] [CrossRef]
- Karalija, E.; Zeljkovic’, S.C.; Paric’, A. Harvest time-related changes in biomass, phenolics and antioxidant potential in Knautia sarajevensis shoot cultures after elicitation with salicylic acid and yeast. In Vitro Cell Dev. Biol. Plant 2019, 56, 177–183. [Google Scholar] [CrossRef]
- Ishikawa, A.; Kitamura, Y.; Ozeki, Y.; Watanabe, M. Different responses of shoot and root cultures of Glehnia littoralis to yeast extract. J. Nat. Med. 2006, 61, 30–37. [Google Scholar] [CrossRef]
- Zaman, G.; Farooq, U.; Bajwa, M.N.; Jan, H.; Shah, M.; Ahmad, R.; Andleeb, A.; Drouet, S.; Hano, C.; Abbasi, B.H. Effects of yeast extract on the production of phenylpropanoid metabolites in callus culture of purple basil (Ocimum basilicum L. var purpurascens) and their in-vitro evaluation for antioxidant potential. Plant Cell Tissue Organ Cult. 2022, 150, 543–553. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Thiruvengadam, M.; Chung, I.M. Identification of elicitors enhances the polyphenolic compounds and pharmacological potential in hairy root cultures of Aster scaber. S. Afr. J. Bot. 2019, 125, 92–101. [Google Scholar] [CrossRef]
- Custodio, L.; Cziaky, Z.; Castaneda-Loaiza, V.; Rodrigues, M.J. Establishment and elicitation of liquid adventitious root cultures of Inula crithmoides L. for increased caffeoylquinic acid production and hepatoprotective properties. Plant Cell Tissue Organ Cult. 2024, 156, 59. [Google Scholar] [CrossRef]
- Zaker, A.; Sykora, C.; Gossnitzer, F.; Abrishamchi, P.; Aili, J.; Mousavi, S.H.; Wawrosch, C. Effects of some elicitors on tanshinone production in adventitious root cultures of Perovskia arbrotanoides Karel. Ind. Crops Prod. 2015, 67, 97–102. [Google Scholar] [CrossRef]
- Ho, T.T.; Lee, J.D.; Jeong, C.S.; Paek, K.Y.; Park, K.Y. Improvement of biosynthesis and accumulation of bioactive compounds by elicitation in adventitious root cultures of Polygonum multiflorum. Appl. Microbiol. Biotechnol. 2018, 102, 199–209. [Google Scholar] [CrossRef]
- Moharramnejad, S.; Sofalian, O.; Valizadeh, M.; Asgari, A.; Shiri, M. Response of antioxidant defense system to osmotic stress in maize seedlings. Fresenius Environ. Bull. 2016, 25, 805–811. [Google Scholar]
- Zhao, P.; Lu, G.H.; Yang, Y.H. Salicylic acid signaling and its role in responses to stresses in plants. Mech. Plant Horm. Signal Under Stress 2017, 39, 413–441. [Google Scholar]
- Ali, B. Salicylic acid: An efficient elicitor of secondary metabolite production in plants. Biocatal. Agric. Biotechnol. 2021, 31, 101884. [Google Scholar] [CrossRef]
- Li, A.; Sun, X.; Liu, L. Action of salicylic acid on plant growth. Front. Plant Sci. 2022, 13, 878076. [Google Scholar] [CrossRef]
- Kováčik, J.; Grúz, J.; Bačkor, M.; Strnad, M.; Repčák, M. Salicylic acid-induced changes to growth and phenolic metabolism in Matricaria chamomilla plants. Plant Cell Rep. 2009, 28, 135–143. [Google Scholar] [CrossRef]
- Bayraktar, M.; Naziri, E.; Akgun, I.H.; Karabey, F.; Ilhan, E.; Akyol, B.; Gurel, A. Elicitor induced stevioside production, in vitro shoot growth, and biomass accumulation in micropropagated Stevia rebaudiana. Plant Cell Tissue Organ Cult. 2016, 127, 289–300. [Google Scholar] [CrossRef]
- Ejtahed, R.; Radjabian, T.; Hoseini Tafreshi, S.A. Expression analysis of phenylalanine ammonia lyase gene and rosmarinic acid production in Salvia officinalis and Salvia virgata shoots under salicylic acid elicitation. Appl. Biochem. Biotechnol. 2015, 176, 1846–1858. [Google Scholar] [CrossRef]
- Dong, J.; Wan, G.; Liang, Z. Accumulation of salicylic acid-induced phenolic compounds and raised activities of secondary metabolic and antioxidative enzymes in Salvia miltiorrhiza cell culture. J. Biotech. 2010, 148, 99–104. [Google Scholar] [CrossRef]
- Gadzovska, S.; Maury, S.; Delaunay, A.; Spasenoski, M.; Hagège, D.; Courtois, D.; Joseph, C. The influence of salicylic acid elicitation of shoots, callus, and cell suspension cultures on production of naphtodianthrones and phenylpropanoids in Hypericum perforatum L. Plant Cell Tissue Organ Cult. 2012, 113, 25–39. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Extract x (mg·L−1) | Fresh Weight | Dry Weight | Growth Ratio |
|---|---|---|---|
| (g·L−1) | (g·L−1) | ||
| 0 | 84.0 c y | 8.5 d | 27.0 c |
| 25 | 89.0 b | 9.2 bc | 28.7 b |
| 50 | 97.3 a | 9.8 a | 31.4 a |
| 100 | 91.2 b | 9.3 b | 29.4 b |
| 200 | 88.3 d | 8.8 c | 28.4 b |
| Salicylic Acid x (μM) | Fresh Weight | Dry Weight | Growth Ratio |
|---|---|---|---|
| (g·L−1) | (g·L−1) | ||
| 0 | 84.0 b y | 8.5 a | 27.0 b |
| 25 | 93.0 a | 9.3 a | 30.0 a |
| 50 | 85.0 b | 9.0 a | 27.3 b |
| 100 | 68.0 c | 7.2 b | 21.7 c |
| 200 | 65.8 c | 6.3 b | 20.9 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, X.; Lee, H.-S.; Han, J.-E.; Murthy, H.N.; Park, S.-Y. Enhancement of Phenolic and Polyacetylene Accumulation in Lobelia chinensis (Chinese lobelia) Plantlet Cultures Through Yeast Extract and Salicylic Acid Elicitation. Horticulturae 2025, 11, 612. https://doi.org/10.3390/horticulturae11060612
Bai X, Lee H-S, Han J-E, Murthy HN, Park S-Y. Enhancement of Phenolic and Polyacetylene Accumulation in Lobelia chinensis (Chinese lobelia) Plantlet Cultures Through Yeast Extract and Salicylic Acid Elicitation. Horticulturae. 2025; 11(6):612. https://doi.org/10.3390/horticulturae11060612
Chicago/Turabian StyleBai, Xinlei, Han-Sol Lee, Jong-Eun Han, Hosakatte Niranjana Murthy, and So-Young Park. 2025. "Enhancement of Phenolic and Polyacetylene Accumulation in Lobelia chinensis (Chinese lobelia) Plantlet Cultures Through Yeast Extract and Salicylic Acid Elicitation" Horticulturae 11, no. 6: 612. https://doi.org/10.3390/horticulturae11060612
APA StyleBai, X., Lee, H.-S., Han, J.-E., Murthy, H. N., & Park, S.-Y. (2025). Enhancement of Phenolic and Polyacetylene Accumulation in Lobelia chinensis (Chinese lobelia) Plantlet Cultures Through Yeast Extract and Salicylic Acid Elicitation. Horticulturae, 11(6), 612. https://doi.org/10.3390/horticulturae11060612