
Identification of the Grapefruit (Citrus paradisi) Isocitrate Dehydrogenase Gene and Functional Analysis of CpNADP-IDH1 in Citric Acid Metabolism



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Bioinformatics Analysis
2.3. RNA Extraction and Gene Expression Analysis
2.4. Expression Vector Construction and Genetic Transformation
2.5. Determination of IDH Enzyme Activity, Soluble Sugar, and Citric Acid Content
2.6. Data Analysis
3. Results
3.1. Determination of Sugar and Acid Content in Two Varieties of Grapefruit
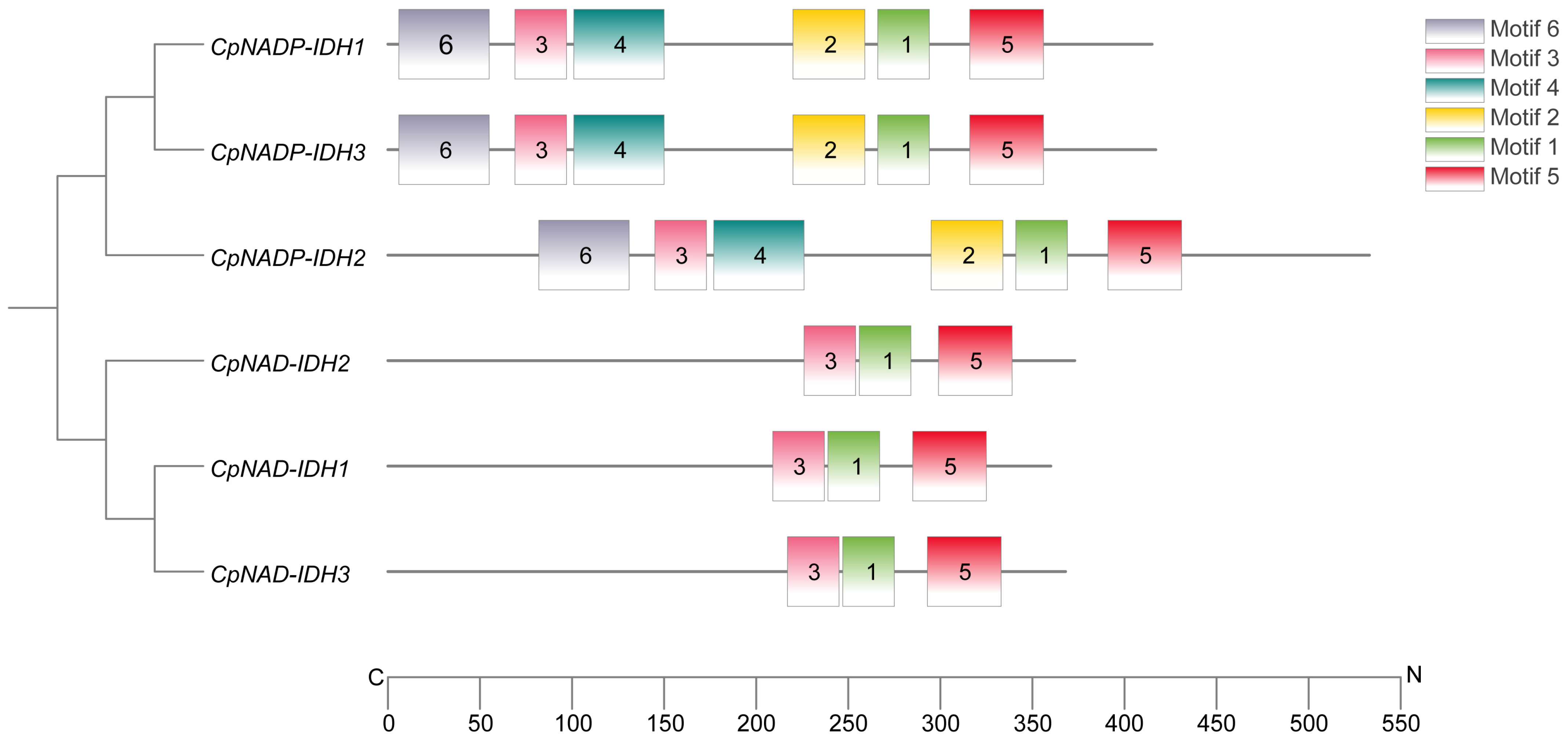
3.2. Identification and Bioinformatics Analysis of the IDH Gene Family in Citrus
3.3. Expression Analysis of IDH Genes
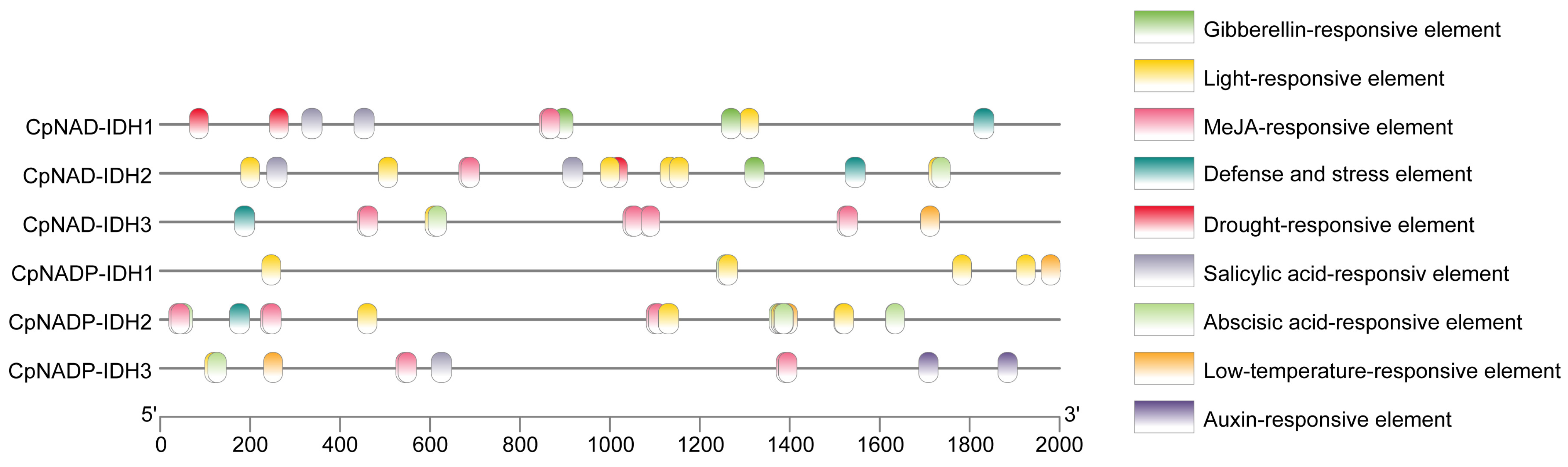
3.4. Promoter Component Analysis of IDHs
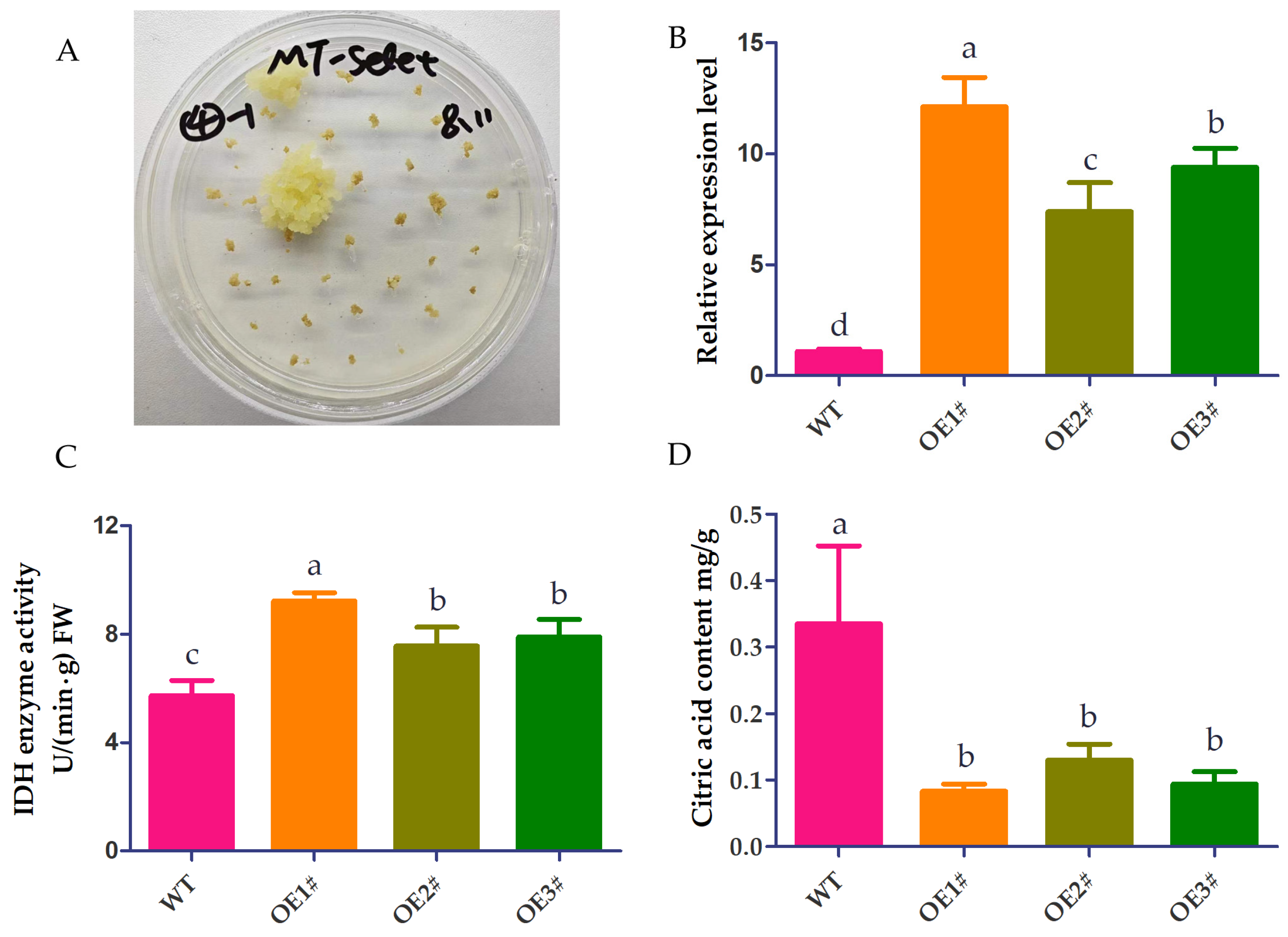
3.5. Functional Analysis of CpNADP-IDH1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Aslan, M.N.; Sukan-Karaçağıl, B.; Acar-Tek, N. Roles of citrus fruits on energy expenditure, body weight management, and metabolic biomarkers: A comprehensive review. Nutr. Rev. 2024, 82, 1292–1307. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; He, F.; Huang, Y.; He, J.; Yang, S.; Zeng, J.; Deng, C.; Jiang, X.; Fang, Y.; Wen, S.; et al. Genome of Wild Mandarin and Domestication History of Mandarin. Mol. Plant. 2018, 11, 1024–1037. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Ma, Q.; Zhou, G.; Liu, X.; Ma, Z.; Gu, Q. Identification of genes associated with soluble sugar and organic acid accumulation in ‘Huapi’ kumquat (Fortunella crassifolia Swingle) via transcriptome analysis. J. Sci. Food Agric. 2021, 101, 4321–4331. [Google Scholar] [CrossRef]
- Lin, Q.; Wang, C.; Dong, W.; Jiang, Q.; Wang, D.; Li, S.; Chen, M.; Liu, C.; Sun, C.; Chen, K. Transcriptome and metabolome analyses of sugar and organic acid metabolism in Ponkan (Citrus reticulata) fruit during fruit maturation. Gene 2015, 554, 64–74. [Google Scholar] [CrossRef]
- Sheng, L.; Shen, D.; Luo, Y.; Sun, X.; Wang, J.; Luo, T.; Zeng, Y.; Xu, J.; Deng, X.; Cheng, Y. Exogenous γ-aminobutyric acid treatment affects citrate and amino acid accumulation to improve fruit quality and storage performance of postharvest citrus fruit. Food Chem. 2017, 216, 138–145. [Google Scholar] [CrossRef]
- Shi, C.; Hussain, S.B.; Han, H.; Alam, S.M.; Liu, D.; Liu, Y. Reduced expression of CsPH8, a P-type ATPase gene, is the major factor leading to the low citrate accumulation in citrus leaves. Plant Physiol. Biochem. 2021, 160, 211–217. [Google Scholar] [CrossRef]
- Shi, C.; Song, R.; Hu, X.; Liu, X.; Jin, L.; Liu, Y. Citrus PH5-like H+-ATPase genes: Identification and transcript analysis to investigate their possible relationship with citrate accumulation in fruits. Front. Plant Sci. 2015, 6, 135. [Google Scholar] [CrossRef]
- Li, S.; Liu, X.; Xie, X.; Sun, C.; Grierson, D.; Yin, X.; Chen, K. CrMYB73, a PH-like gene, contributes to citric acid accumulation in citrus fruit. Sci. Hortic. 2015, 197, 212–217. [Google Scholar] [CrossRef]
- Hussain, S.B.; Shi, C.; Guo, L.; Kamran, H.M.; Sadka, A.; Liu, Y. Recent Advances in the Regulation of Citric Acid Metabolism in Citrus Fruit. Crit. Rev. Plant Sci. 2017, 36, 241–256. [Google Scholar] [CrossRef]
- Sheng, L.; Shen, D.; Yang, W.; Zhang, M.; Zeng, Y.; Xu, J.; Deng, X.; Cheng, Y. GABA Pathway Rate-Limit Citrate Degradation in Postharvest Citrus Fruit Evidence from HB Pumelo (Citrus grandis) × Fairchild (Citrus reticulata) Hybrid Population. J. Agric. Food Chem. 2017, 65, 1669–1676. [Google Scholar] [CrossRef]
- Lu, X.; Cao, X.; Li, F.; Li, J.; Xiong, J.; Long, G.; Cao, S.; Xie, S. Comparative transcriptome analysis reveals a global insight into molecular processes regulating citrate accumulation in sweet orange (Citrus sinensis). Physiol. Plant. 2016, 158, 463–482. [Google Scholar] [CrossRef] [PubMed]
- Sadka, A.; Dahan, E.; Or, E.; Cohen, L. NADP+-isocitrate dehydrogenase gene expression and isozyme activity during citrus fruit development. Plant Sci. 2000, 158, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yin, X.; Wang, W.; Liu, X.; Zhang, B.; Chen, K. Citrus CitNAC62 cooperates with CitWRKY1 to participate in citric acid degradation via up-regulation of CitAco3. J. Exp. Bot. 2017, 68, 3419–3426. [Google Scholar] [CrossRef]
- Lancien, M.; Gadal, P.; Hodges, M. Enzyme redundancy and the importance of 2-oxoglutarate in higher plant ammonium assimilation. Plant Physiol. 2000, 123, 817–824. [Google Scholar] [CrossRef]
- Zhan, S.; Zhang, Q.; Yao, Y.; Cui, Y.; Huang, T. Cytosolic isocitrate dehydrogenase regulates plant stem cell maintenance in response to nutrient deficiency. Plant Physiol. 2023, 192, 3069–3087. [Google Scholar] [CrossRef]
- Muñoz-Vargas, M.A.; González-Gordo, S.; Taboada, J.; Palma, J.M.; Corpas, F.J. Activity and gene expression analysis of the NADP-dependent isocitrate dehydrogenase (NADP-ICDH) through pepper fruit ripening and its modulation by nitric oxide (NO). Molecular characterization of the peroxisomal isozyme. Plant Sci. 2024, 349, 112269. [Google Scholar] [CrossRef]
- Nilo, R.; Saffie, C.; Lilley, K.; Baeza-Yates, R.; Cambiazo, V.; Campos-Vargas, R.; González, M.; Meisel, L.A.; Retamales, J.; Silva, H.; et al. Proteomic analysis of peach fruit mesocarp softening and chilling injury using difference gel electrophoresis (DIGE). BMC Genom. 2010, 11, 43. [Google Scholar] [CrossRef]
- Margaria, P.; Abba, S.; Palmano, S. Novel aspects of grapevine response to phytoplasma infection investigated by a proteomic and phospho-proteomic approach with data integration into functional networks. BMC Genom. 2013, 14, 38. [Google Scholar] [CrossRef]
- Castañeda, V.; González, E.M. Strategies to Apply Water-Deficit Stress: Similarities and Disparities at the Whole Plant Metabolism Level in Medicago truncatula. Int. J. Mol. Sci. 2021, 22, 2813. [Google Scholar] [CrossRef]
- Leterrier, M.; Del Río, L.A.; Corpas, F.J. Cytosolic NADP-isocitrate dehydrogenase of pea plants: Genomic clone characterization and functional analysis under abiotic stress conditions. Free Radic. Res. 2007, 41, 191–199. [Google Scholar] [CrossRef]
- Wei, N.; Zhang, Z.; Yang, H.; Hu, D.; Wu, Y.; Xue, J.; Guo, D.; Xu, S. Characterization of the Isocitrate Dehydrogenase Gene Family and Their Response to Drought Stress in Maize. Plants 2023, 12, 3466. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.M.; Gupta, P. Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Edgar, R.C.; Batzoglou, S. Multiple sequence alignment. Curr. Opin. Struc. Biol. 2006, 16, 368–373. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Long, J.; Liu, C.; Feng, M.; Liu, Y.; Wu, X.; Guo, W. miR156-SPL modules regulate induction of somatic embryogenesis in citrus callus. J. Exp. Bot. 2018, 69, 2979–2993. [Google Scholar] [CrossRef]
- Tang, M.; Bie, Z.; Wu, M.; Yi, H.; Feng, J. Changes in organic acids and acid metabolism enzymes in melon fruit during development. Sci. Hortic. 2010, 123, 360–365. [Google Scholar] [CrossRef]
- Popova, O.V.; Ismailov, S.F.; Popova, T.N.; Dietz, K.; Golldack, D. Salt-induced expression of NADP-dependent isocitrate dehydrogenase and ferredoxin-dependent glutamate synthase in Mesembryanthemum crystallinum. Planta 2002, 215, 906–913. [Google Scholar] [CrossRef]
- Wu, B.; Qi, F.; Liang, Y. Fuels for ROS signaling in plant immunity. Trends Plant Sci. 2023, 28, 1124–1131. [Google Scholar] [CrossRef]
- Zhang, W.; Gong, Z.; Wu, M.; Chan, H.; Yuan, Y.; Tang, N.; Zhang, Q.; Miao, M.; Chang, W.; Li, Z.; et al. Integrative comparative analyses of metabolite and transcript profiles uncovers complex regulatory network in tomato (Solanum lycopersicum L.) fruit undergoing chilling injury. Sci. Rep. 2019, 9, 4470. [Google Scholar] [CrossRef] [PubMed]
- Gamrasni, D.; Erov, M.; Saar, L.; Raz, A.; Glikman, M.; Sonawane, P.D.; Aharoni, A.; Goldway, M. The isocitrate dehydrogenase 1 gene is associated with the climacteric response in tomato fruit ripening. Postharvest Biol. Technol. 2020, 166, 111219. [Google Scholar] [CrossRef]
- Jiang, N.; Jin, L.; Da Silva, J.A.T.; Islam, M.Z.; Gao, H.; Liu, Y.; Peng, S. Activities of enzymes directly related with sucrose and citric acid metabolism in citrus fruit in response to soil plastic film mulch. Sci. Hortic. 2014, 168, 73–80. [Google Scholar] [CrossRef]
- Wang, X.; Yin, W.; Wu, J.; Chai, L.; Yi, H. Effects of exogenous abscisic acid on the expression of citrus fruit ripening-related genes and fruit ripening. Sci. Hortic. 2016, 201, 175–183. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, L.; Yue, Y.; Liu, F.; Wen, M.; Huang, B.; Wang, P. Identification of the Grapefruit (Citrus paradisi) Isocitrate Dehydrogenase Gene and Functional Analysis of CpNADP-IDH1 in Citric Acid Metabolism. Horticulturae 2025, 11, 598. https://doi.org/10.3390/horticulturae11060598
Jin L, Yue Y, Liu F, Wen M, Huang B, Wang P. Identification of the Grapefruit (Citrus paradisi) Isocitrate Dehydrogenase Gene and Functional Analysis of CpNADP-IDH1 in Citric Acid Metabolism. Horticulturae. 2025; 11(6):598. https://doi.org/10.3390/horticulturae11060598
Chicago/Turabian StyleJin, Longfei, Yang Yue, Feng Liu, Mingxia Wen, Bei Huang, and Peng Wang. 2025. "Identification of the Grapefruit (Citrus paradisi) Isocitrate Dehydrogenase Gene and Functional Analysis of CpNADP-IDH1 in Citric Acid Metabolism" Horticulturae 11, no. 6: 598. https://doi.org/10.3390/horticulturae11060598
APA StyleJin, L., Yue, Y., Liu, F., Wen, M., Huang, B., & Wang, P. (2025). Identification of the Grapefruit (Citrus paradisi) Isocitrate Dehydrogenase Gene and Functional Analysis of CpNADP-IDH1 in Citric Acid Metabolism. Horticulturae, 11(6), 598. https://doi.org/10.3390/horticulturae11060598