
The Role of Root Endophyte Pseudomonas putida A32 in the Protection of Two Pepper Genotypes from Pseudomonas syringae pv. aptata
 ,
,  ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Antibacterial Activity In Vitro Assay
2.3. Bacterial Treatment and Infection Susceptibility Assay
2.4. Quantification of Lipid Peroxidation and Hydrogen Peroxide in Pepper Leaves
2.5. Antioxidant Enzyme Activity
2.6. Determination of Phenylalanine Ammonia Lyase Activity
2.7. Statistical Analysis
3. Results
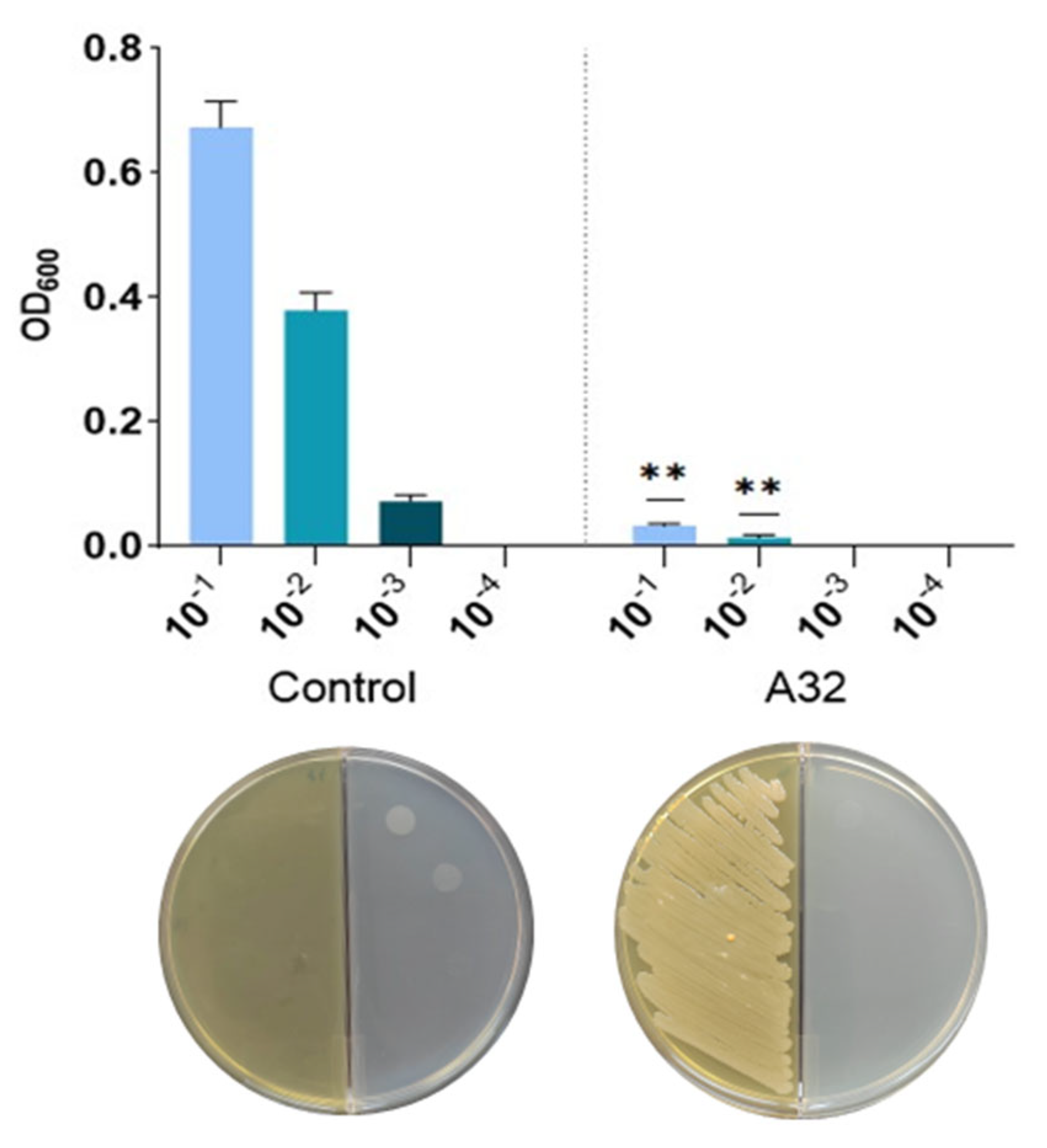
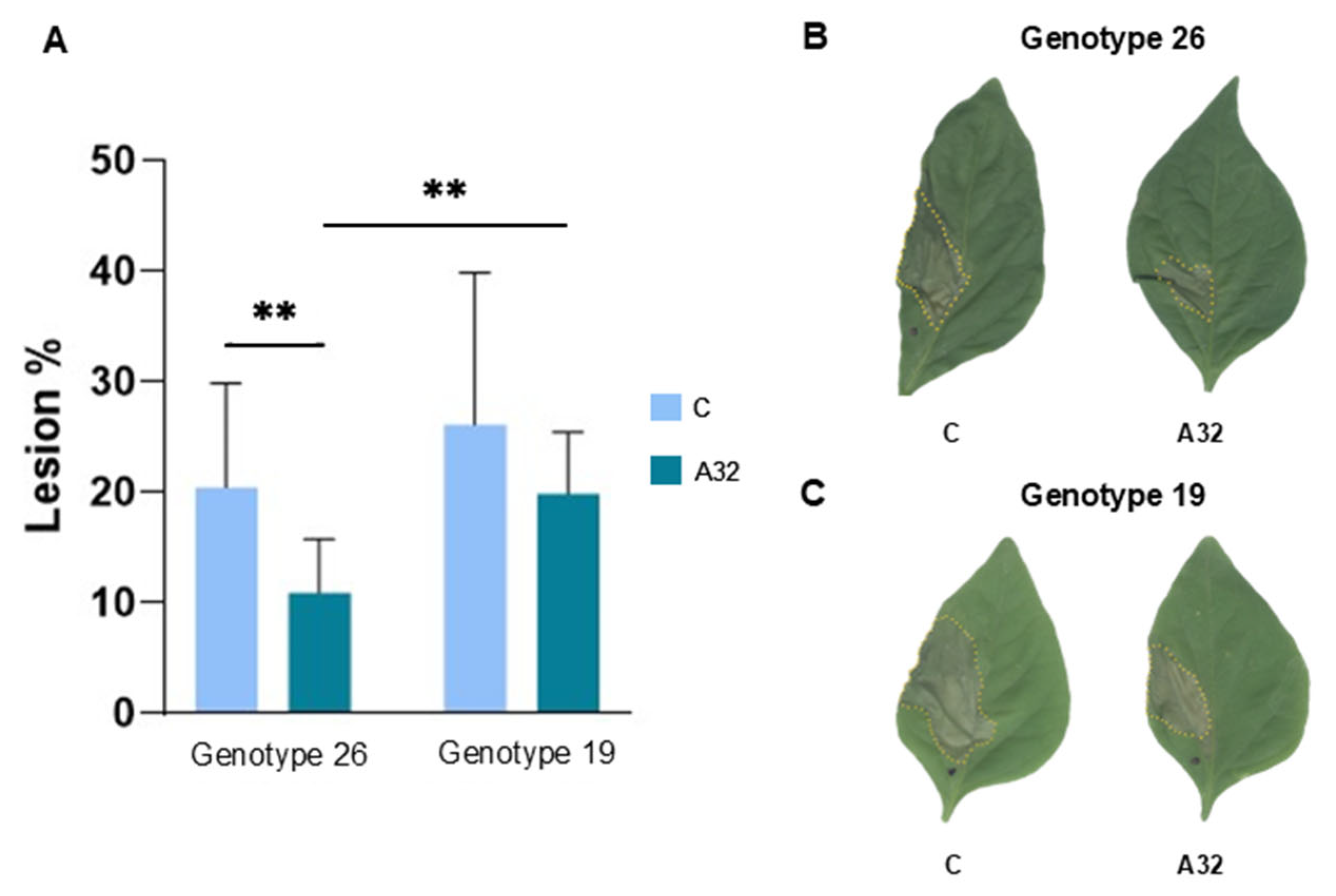
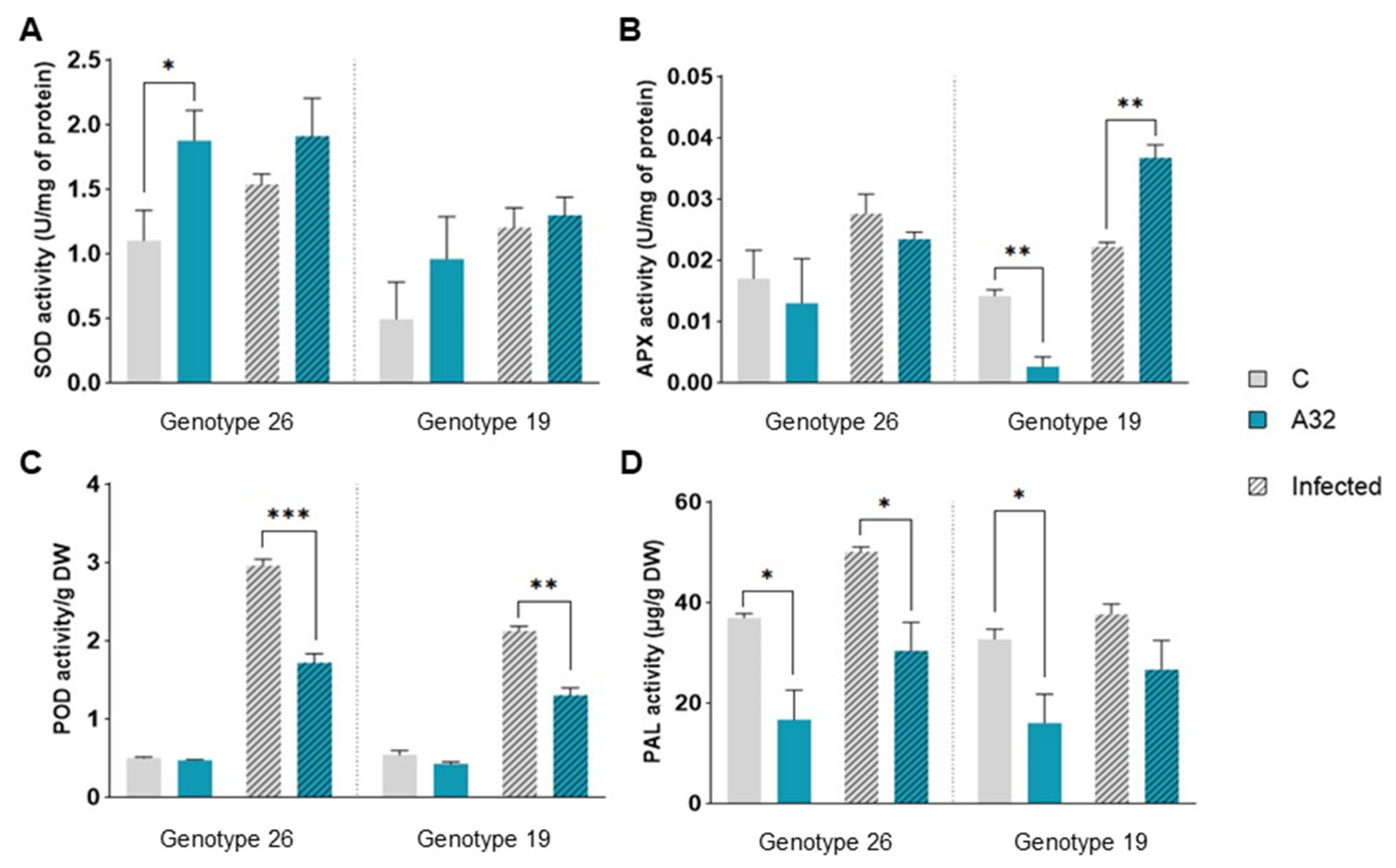
In Vitro and In Planta Antagonistic Activity of Pseudomonas Putida A32 Against Phytopathogen Bacteria Pseudomonas syringae pv. aptata P21
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lushchak, V.I.; Matviishyn, T.M.; Husak, V.V.; Storey, J.M.; Storey, K.B. Pesticide toxicity: A mechanistic approach. EXCLI J. 2018, 17, 1101–1136. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Niranjana, M.; Jha, S.; Mallick, N.; Agarwal, P. QTL detection and putative candidate gene prediction for leaf rolling under moisture stress condition in wheat. Sci. Rep. 2020, 10, 18696. [Google Scholar] [CrossRef]
- Laishram, B.; Devi, O.R.; Dutta, R.; Senthilkumar, T.; Goyal, G.; Paliwal, D.K.; Panotra, N.; Rasool, A. Plant-microbe interactions: PGPM as microbial inoculants/biofertilizers for sustaining crop productivity and soil fertility. Curr. Res. Microb. Sci. 2024, 18, 100333. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Zaib, S. Mighty Microbes: Plant Growth Promoting Microbes in Soil Health and Sustainable Agriculture. In Soil Health. Soil Biology; Giri, B., Varma, A., Eds.; Springer: Cham, Switzerland, 2020; Volume 59. [Google Scholar] [CrossRef]
- Zhao, Y.; Liao, H.; Ran, T.; Yang, H. Changes in rhizosphere soil bacterial, fungal, and protistan communities during tomato (Solanum lycopersicum) growth after reductive soil disinfestation. Plant Soil. 2025, 506, 573–589. [Google Scholar] [CrossRef]
- Ali, A.; Elrys, A.S.; Liu, L.; Xia, Q.; Wang, B.; Li, Y.; Dan, X.; Iqbal, M.; Zhao, J.; Huang, X.; et al. Deciphering the synergies of reductive soil disinfestation combined with biochar and antagonistic microbial inoculation in cucumber Fusarium wilt suppression through rhizosphere microbiota structure. Microb. Ecol. 2022, 85, 980–997. [Google Scholar] [CrossRef] [PubMed]
- Pascale, A.; Proietti, S.I.; Pantelides, S.; Stringlis, I.A. Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef]
- Simon, J.C.; Marchesi, J.R.; Mougel, C.; Selosse, M.A. Host-Microbiota interactions: From holobiont theory to analysis. Microbiome 2019, 7, 5. [Google Scholar] [CrossRef]
- Scagliola, M.; Valentinuzzi, F.; Mimmo, T.; Cesco, S.; Crecchio, C.; Pii, Y. Bioinoculants as promising complement of chemical fertilizers for a more sustainable agricultural practice. Front. Sustain. Food Syst. 2021, 4, 622169. [Google Scholar] [CrossRef]
- Bonaterra, A.; Badosa, E.; Daranas, N.; Francés, J.; Roselló, G.; Montesinos, E. Bacteria as biological control agents of plant diseases. Microorganisms 2022, 10, 1759. [Google Scholar] [CrossRef]
- Sahgal, M.; Saini, N.; Jaggi, V.; Brindhaa, N.T.; Kabdwal, M.; Singh, R.P.; Prakash, A. Antagonistic potential and biological control mechanisms of Pseudomonas strains against banded leaf and sheath blight disease of maize. Sci. Rep. 2024, 14, 13580. [Google Scholar] [CrossRef]
- Pandey, S.; Gupta, S. Evaluation of Pseudomonas sp. for its multifarious plant growth promoting potential and its ability to alleviate biotic and abiotic stress in tomato (Solanum lycopersicum) plants. Sci. Rep. 2020, 10, 20951. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef]
- Nikolić, I.; Stanković, S.; Dimkić, I.; Berić, T.; Stojšin, V.; Janse, J.; Popović, T. Genetic diversity and pathogenicity of Pseudomonas syringae pv aptata isolated from sugar beet. Plant Pathol. 2018, 67, 1194–1207. [Google Scholar] [CrossRef]
- Nikolić, I.; Glatter, T.; Ranković, T.; Berić, T.; Stanković, S.; Diepold, A. Repertoire and abundance of secreted virulence factors shape the pathogenic capacity of Pseudomonas syringae pv aptata. Front. Microbiol. 2023, 14, 1205257. [Google Scholar] [CrossRef]
- Nikolić, I.; Berić, T.; Dimkić, I.; Popović, T.; Lozo, J.; Fira, D.; Stanković, S. Biological control of Pseudomonas syringae pv. aptata on sugar beet with Bacillus pumilus SS-10.7 and Bacillus amyloliquefaciens (SS-12.6 and SS-38.4) strains. J. Appl. Microbiol. 2019, 126, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Nedeljković, M.; Mesaroš, A.; Rašić, V.; Nikolić, I.; Stanković, S.; Lozo, J.; Atanasković, I. Effects of T3SS-positive Pseudomonas isolates on sugar beet growth stimulation and pathogen resistance. Plant Soil. 2024. [Google Scholar] [CrossRef]
- Mesaroš, A.; Atanasković, I.; Nedeljković, M.; Stanković, S.; Lozo, J. Differential responses of bell pepper genotypes to indigenous Pseudomonas putida A32 treatment: Implications for drought resilience. J. Appl. Microbiol. 2024, 135, lxae190. [Google Scholar] [CrossRef]
- Krstić Tomić, T.; Atanasković, I.; Nikolić, I.; Joković, N.; Stević, T.; Stanković, S.; Berić, T.; Lozo, J. Culture-dependent and metabarcoding characterization of the sugar beet (Beta vulgaris L.) microbiome for high-yield isolation of bacteria with plant growth-promoting traits. Microorganisms 2023, 11, 1538. [Google Scholar] [CrossRef]
- Létoffé, S.; Wu, Y.; Darch, S.E.; Beloin, C.; Whiteley, M.; Touqui, L.; Ghigo, J.M. Pseudomonas aeruginosa production of hydrogen cyanide leads to airborne control of Staphylococcus aureus growth in biofilm and in vivo lung environments. mBio 2022, 13, e0215422. [Google Scholar] [CrossRef]
- Mesaroš, A.; Nedeljković, M.; Danojević, D.; Medić-Pap, S.; Stanković, S.; Radović, S.; Lozo, J. Influence of growth conditions on an antioxidative system in two bell pepper genotypes differing in susceptibility to phytopathogen bacteria Xanthomonas euvesicatoria. Plant Growth Regul. 2023, 100, 609–617. [Google Scholar] [CrossRef]
- Hyder, S.; Gondal, A.S.; Rizvi, Z.F.; Atiq, R.; Haider, M.I.S.; Fatima, N.; Inam-ul-Haq, M. Biological control of chili damping-off disease, caused by Pythium myriotylum. Front. Microbiol. 2021, 12, 587431. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nature Meth. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Lozo, J.; Danojević, D.; Jovanović, Ž.; Nenadović, Ž.; Fira, D.; Stanković, S.; Radović, S. Genotype-dependent antioxidative response of four sweet pepper cultivars to water deficiency as affected by drought-tolerant Bacillus safensis SS-2.7 and Bacillus thuringien sis SS-29.2 strains. Horticulturae 2022, 8, 236. [Google Scholar] [CrossRef]
- Kleinhofs, A.; Haskins, F.A.; Gorz, H.J. Relationship of Phenylalanine ammonia-lyase activity to o-hydroxycinnamic acid content in Melilotus alba. Plant Physiol. 1966, 41, 1276–1279. [Google Scholar] [CrossRef]
- Soth, S.; Hampton, J.G.; Alizadeh, H.; Wakelin, S.A.; Mendoza-Mendoza, A. Microbiomes in action: Multifaceted benefits and challenges across academic disciplines. Front. Microbiol. 2025, 16, 1550749. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant-microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Tack, A.J.; Wasserman, B.; Liu, J.; Berg, G.; Norelli, J.; Droby, S.; Wisniewski, M. Evidence for host–microbiome co-evolution in apple. New Phytol. 2022, 234, 2088–2100. [Google Scholar] [CrossRef]
- Pal, G.; Saxena, S.; Kumar, K.; Verma, A.; Sahu, P.K.; Pandey, A.; White, J.F.; Verma, S.K. Endophytic Burkholderia: Multifunctional roles in plant growth promotion and stress tolerance. Microbiol. Res. 2022, 265, 127201. [Google Scholar] [CrossRef]
- Sehrawat, A.; Sindhu, S.S.; Glick, B.R. Hydrogen cyanide production by soil bacteria: Biological control of pests and promotion of plant growth in sustainable agriculture. Pedosphere 2022, 32, 15–38. [Google Scholar] [CrossRef]
- Díaz-Rueda, P.; Morales de los Ríos, L.; Romero, C.; García, I. Old poisons, new signaling molecules: The case of hydrogen cyanide. J. Exp. Bot. 2023, 74, 6040–6051. [Google Scholar] [CrossRef]
- Kandel, S.L.; Joubert, P.M.; Doty, S.L. Bacterial endophyte colonization and distribution within plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef]
- Fang, R.; Lin, J.; Yao, S.; Wang, Y.; Wang, J.; Zhou, C.; Wang, H.; Xiao, M. Promotion of plant growth, biological control and induced systemic resistance in maize by Pseudomonas aurantiaca JD37. Ann. Microbiol. 2013, 63, 1177–1185. [Google Scholar] [CrossRef]
- Ghadamgahi, F.; Tarighi, S.; Taheri, P.; Saripella, G.V.; Anzalone, A.; Kalyandurg, P.B.; Catara, V.; Ortiz, R.; Vetukuri, R.R. Plant growth-promoting activity of Pseudomonas aeruginosa FG106 and its ability to act as a biocontrol agent against potato, tomato and taro pathogens. Biology 2022, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Karpinska, B.; Foyer, C.H. Superoxide signalling and antioxidant processing in the plant nucleus. J. Exp. Bot. 2024, 75, 4599–4610. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 6. [Google Scholar] [CrossRef]
- Popržen, T.; Jevremović, S.; Milošević, S.; Ðurić, M.; Uzelac, B.; Stanković, S.; Radulović, O. Antioxidative response of duckweed (Lemna minor L.) to rhizosphere- associated Pseudomonas strains and exogenous indole-3-acetic acid. Horticulturae 2024, 10, 562. [Google Scholar] [CrossRef]
- Popržen, T.; Antonić Reljin, D.; Uzelac, B.; Milovančević, M.; Paunović, D.; Trifunović-Momčilov, M.; Marković, M.; Raspor, M.; Nikolić, I.; Stanković, S.; et al. Pseudomonas oryzihabitans D1-104/3 and P. gessardii C31-106/3 differentially modulate the antioxidative response of duckweed (Lemna minor L.) to salt stress. Front. Microbiol. 2024, 15, 1481437. [Google Scholar] [CrossRef] [PubMed]
- Ajijah, N.; Fiodor, A.; Dziewit, L.; Pranaw, K. Biological amelioration of water stress in rapeseed (Brassica napus L.) by exopolysaccharides-producing Pseudomonas protegens ML15. Physiol. Plant. 2024, 176, e70012. [Google Scholar] [CrossRef]
- Ayaz, M.; Ali, Q.; Zhao, W.; Chi, Y.-K.; Ali, F.; Rashid, K.A.; Cao, S.; He, Y.-q.; Bukero, A.A.; Huang, W.-K.; et al. Exploring plant growth promoting traits and biocontrol potential of new isolated Bacillus subtilis BS-2301 strain in suppressing Sclerotinia sclerotiorum through various mechanisms. Front. Plant Sci. 2024, 15, 1444328. [Google Scholar] [CrossRef]
- Gamalero, E.; Glick, B.R. Recent advances in bacterial amelioration of plant drought and salt Stress. Biology 2022, 11, 437. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.D.; Franco, O.L. Pathogenesis-related proteins (PRs) with enzyme activity activating plant defense responses. Plants 2023, 12, 2226. [Google Scholar] [CrossRef] [PubMed]
- Anuar, M.S.K.; Hashim, A.M.; Ho, C.L.; Wong, M.Y.; Sundram, S.; Saidi, N.B.; Yusof, M.T. Synergism: Biocontrol agents and biostimulants in reducing abiotic and biotic stresses in crop. World J. Microbiol. Biotechnol. 2023, 39, 123. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesaroš, A.; Nedeljković, M.; Atanasković, I.; Anđelković, M.; Danojević, D.; Stanković, S.; Lozo, J. The Role of Root Endophyte Pseudomonas putida A32 in the Protection of Two Pepper Genotypes from Pseudomonas syringae pv. aptata. Horticulturae 2025, 11, 536. https://doi.org/10.3390/horticulturae11050536
Mesaroš A, Nedeljković M, Atanasković I, Anđelković M, Danojević D, Stanković S, Lozo J. The Role of Root Endophyte Pseudomonas putida A32 in the Protection of Two Pepper Genotypes from Pseudomonas syringae pv. aptata. Horticulturae. 2025; 11(5):536. https://doi.org/10.3390/horticulturae11050536
Chicago/Turabian StyleMesaroš, Aleksandra, Marija Nedeljković, Iva Atanasković, Marija Anđelković, Dario Danojević, Slaviša Stanković, and Jelena Lozo. 2025. "The Role of Root Endophyte Pseudomonas putida A32 in the Protection of Two Pepper Genotypes from Pseudomonas syringae pv. aptata" Horticulturae 11, no. 5: 536. https://doi.org/10.3390/horticulturae11050536
APA StyleMesaroš, A., Nedeljković, M., Atanasković, I., Anđelković, M., Danojević, D., Stanković, S., & Lozo, J. (2025). The Role of Root Endophyte Pseudomonas putida A32 in the Protection of Two Pepper Genotypes from Pseudomonas syringae pv. aptata. Horticulturae, 11(5), 536. https://doi.org/10.3390/horticulturae11050536