
MIKC-Type MADS-box Genes Regulate Phytohormone-Dependent Fruit Ripening in Tomatoes



Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of the MIKC-Type MADS-box Genes in Tomatoes
2.2. Phylogenetic Relationship
2.3. Analysis of Exon–Intron Structures and Conserved Motifs
2.4. Analysis of Promoter Cis-Acting Elements
2.5. Expression Patterns of MIKC-Type MADS-box Genes at Different Growth Stages
2.6. Plant Materials and Growth Conditions
2.7. Virus-Induced Gene Silencing (VIGS)
2.8. Carotenoid (Car) Content Measurement
2.9. Extraction and Determination of Phytohormones
2.10. Statistical Analysis
3. Results
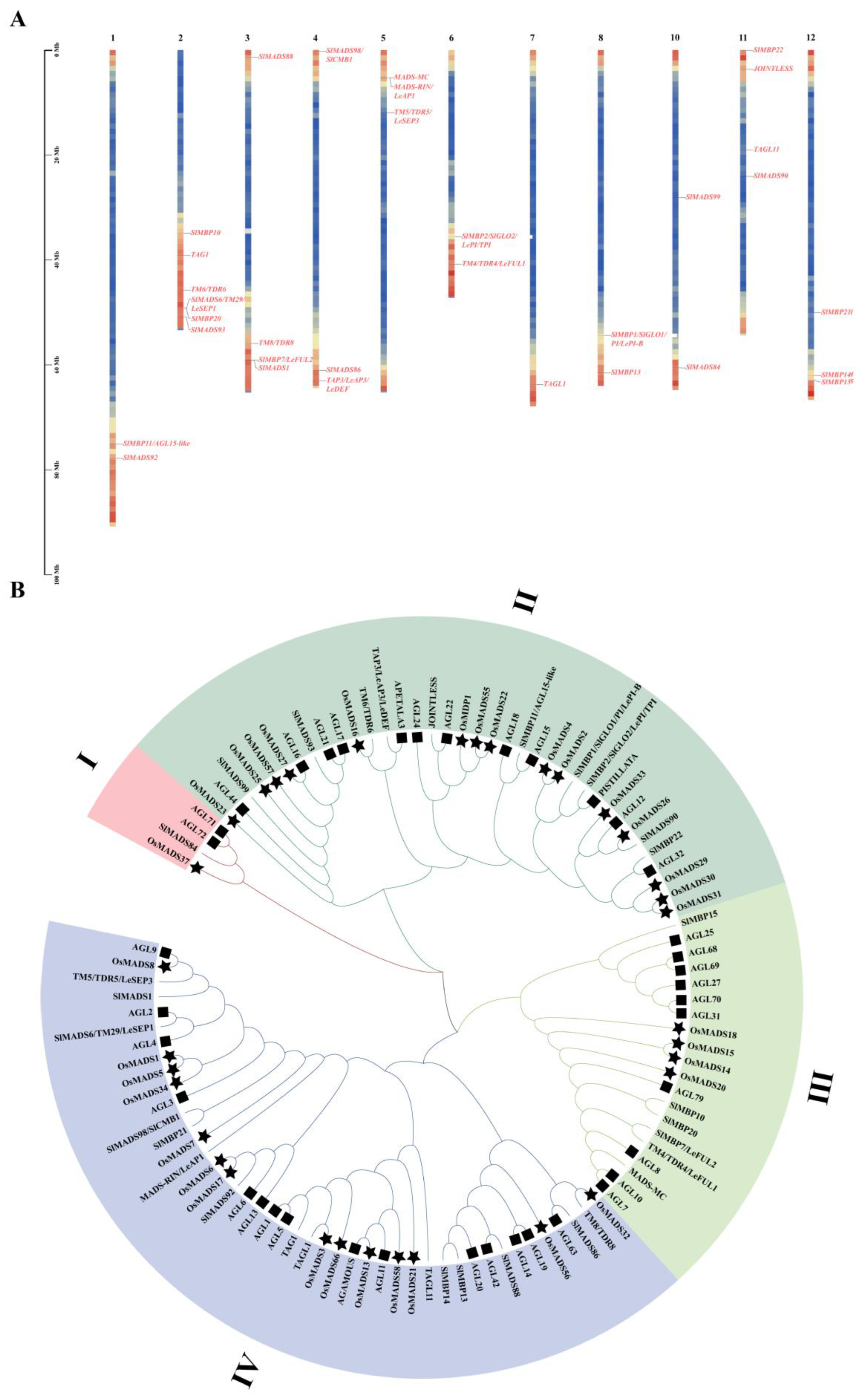
3.1. Genome-Wide Identification and Phylogenetic Analyses of MIKC-Type MADS-box Genes
3.2. Exon–Intron Structure, Conserved Motifs, and Cis-Acting Elements of MIKC-Type MADS-box Genes
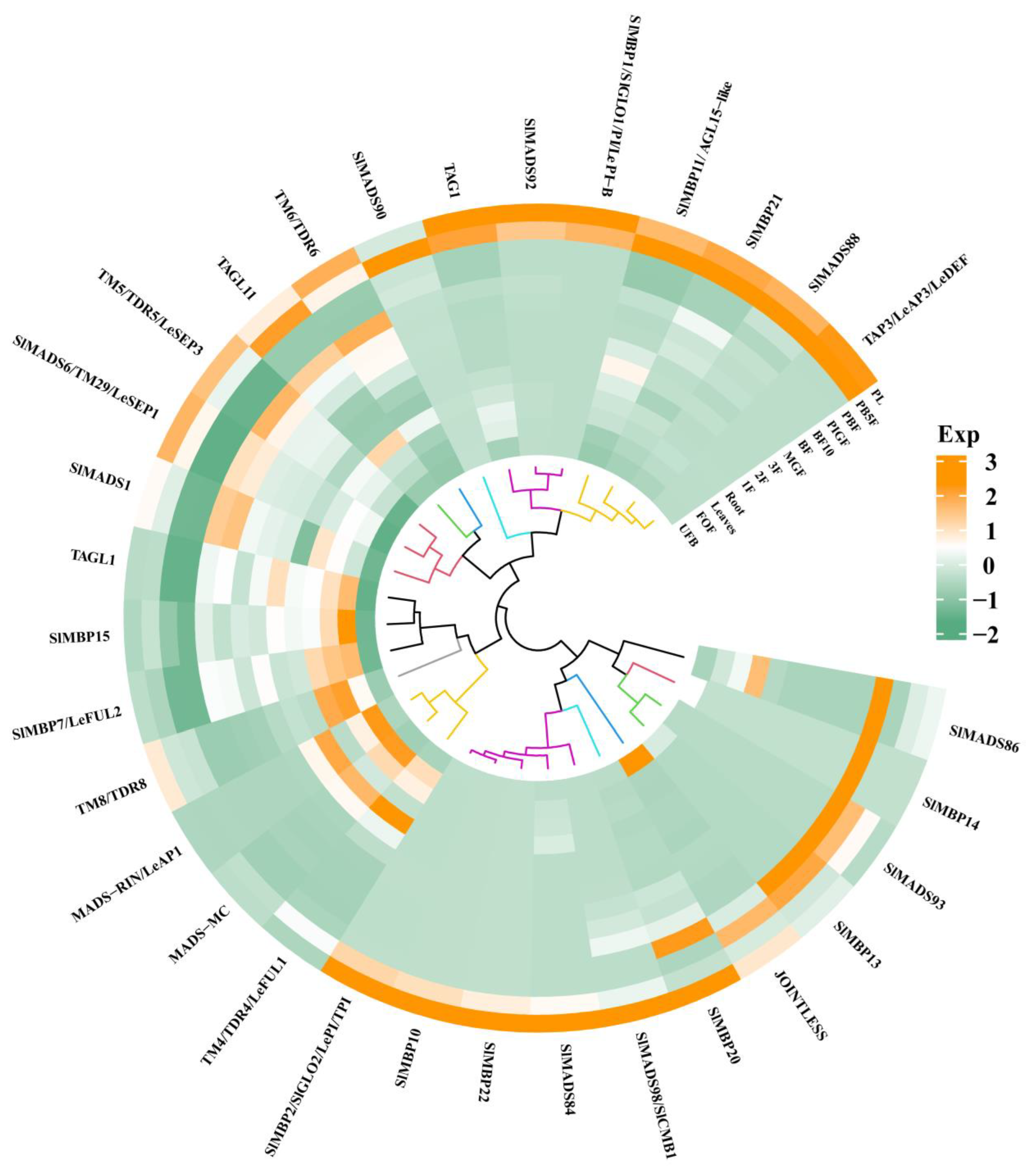
3.3. Expression Patterns of MIKC-Type MADS-box Genes During Growth and Development
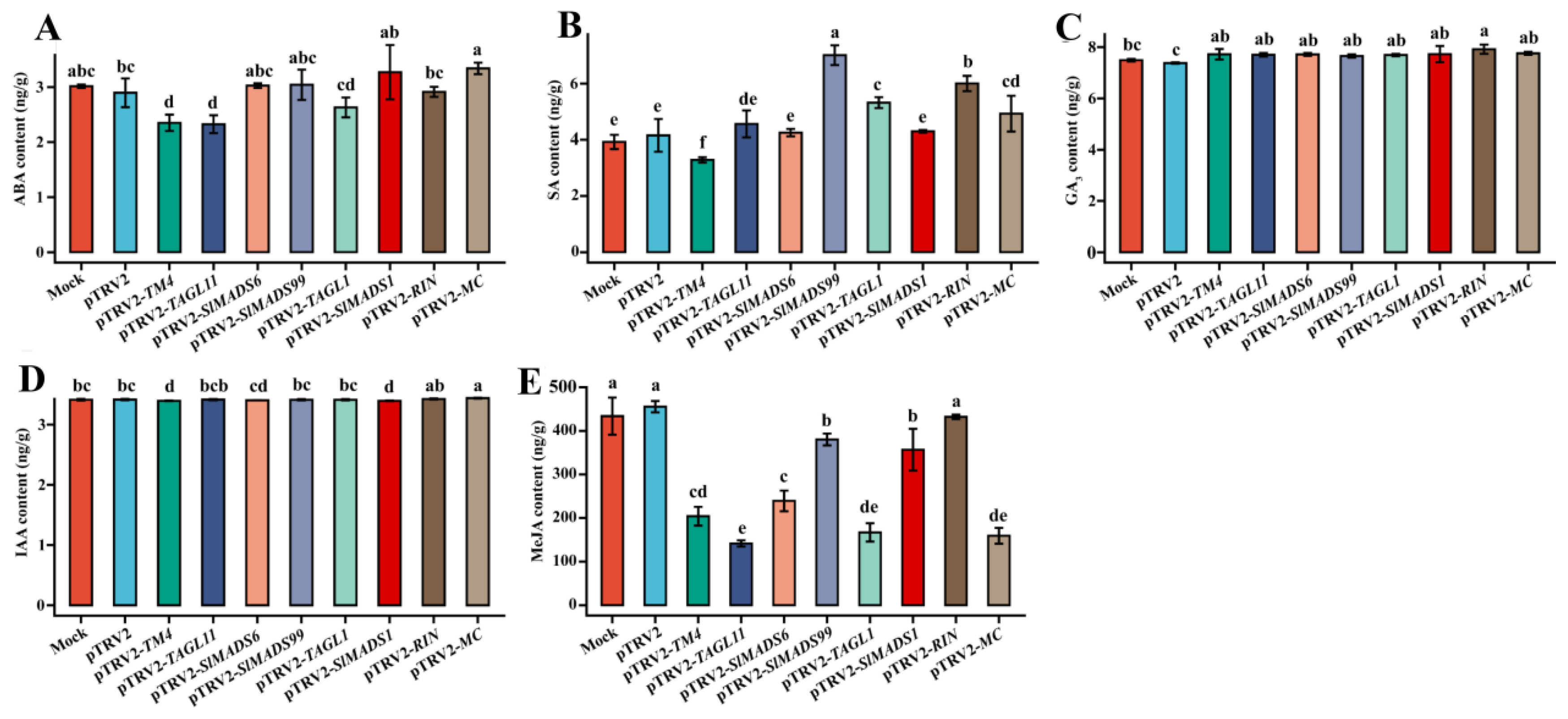
3.4. Silencing of MADS-box Genes Affects Tomato Fruit Ripening
3.5. Silencing of MADS-box Genes Regulated Endogenous Plant Hormone Contents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cara, B.; Giovannoni, J.J. Molecular biology of ethylene during tomato fruit development and maturation. Plant Sci. 2008, 175, 106–113. [Google Scholar] [CrossRef]
- Yokotani, N.; Nakano, R.; Imanishi, S.; Nagata, M.; Inaba, A.; Kubo, Y. Ripening-associated ethylene biosynthesis in tomato fruit is autocatalytically and developmentally regulated. J. Exp. Bot. 2009, 60, 3433–3442. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.; Yang, S.; Chai, L.; Wu, C.; Zhou, J.; Liu, Y.; Xue, Z. Abscisic acid and fruit ripening: Multifaceted analysis of the effect of abscisic acid on fleshy fruit ripening. Sci. Hortic. 2021, 281, 109999. [Google Scholar] [CrossRef]
- Shi, H.Y.; Cao, L.W.; Xu, Y.; Yang, X.; Liu, S.L.; Liang, Z.S.; Li, G.C.; Yang, Y.P.; Zhang, Y.X.; Chen, L. Transcriptional profiles underlying the effects of salicylic acid on fruit ripening and senescence in pear (Pyrus pyrifolia Nakai). J. Integr. Agric. 2021, 20, 2424–2437. [Google Scholar] [CrossRef]
- Wu, M.; Liu, K.; Li, H.; Li, Y.; Zhu, Y.; Su, D.; Zhang, Y.; Deng, H.; Wang, Y.; Liu, M. Gibberellins involved in fruit ripening and softening by mediating multiple hormonal signals in tomato. Hortic. Res. 2023, 11, uhad275. [Google Scholar] [CrossRef]
- Böttcher, C.; Keyzers, R.A.; Boss, P.K.; Davies, C. Sequestration of auxin by the indole-3-acetic acid-amido synthetase GH3-1 in grape berry (Vitis vinifera L.) and the proposed role of auxin conjugation during ripening. J. Exp. Bot. 2010, 61, 3615–3625. [Google Scholar] [CrossRef] [PubMed]
- Zuñiga, P.E.; Castañeda, Y.; Arrey-Salas, O.; Fuentes, L.; Aburto, F.; Figueroa, C.R. Methyl jasmonate applications from flowering to ripe fruit stages of strawberry (Fragaria × ananassa ‘Camarosa’) reinforce the fruit antioxidant response at post-harvest. Front. Plant Sci. 2020, 11, 538. [Google Scholar] [CrossRef]
- Tigchelaar, E.; Tomes, M.; Kerr, E.; Barman, R. A new fruit ripening mutant, non-ripening (nor). Rep. Tomato Genet. Coop. 1973, 23, 33. [Google Scholar]
- Wilkinson, J.Q.; Lanahan, M.B.; Yen, H.C.; Giovannoni, J.J.; Klee, H.J. An ethylene-inducible component of signal transduction encoded by never-ripe. Science 1995, 270, 1807–1809. [Google Scholar] [CrossRef]
- Vrebalov, J.; Ruezinsky, D.; Padmanabhan, V.; White, R.; Medrano, D.; Drake, R.; Schuch, W.; Giovannoni, J. A MADS-box gene necessary for fruit ripening at the tomato ripening-inhibitor (rin) locus. Science 2002, 296, 343–346. [Google Scholar] [CrossRef]
- Messenguy, F.; Dubois, E. Role of MADS box proteins and their cofactors in combinatorial control of gene expression and cell development. Gene 2003, 316, 1–21. [Google Scholar] [CrossRef]
- Alvarez-Buylla, E.R.; Pelaz, S.; Liljegren, S.J.; Gold, S.E.; Burgeff, C.; Ditta, G.S.; de Pouplana, L.R.; Martínez-Castilla, L.; Yanofsky, M.F. An ancestral MADS-box gene duplication occurred before the divergence of plants and animals. Proc. Natl. Acad. Sci. USA 2000, 97, 5328–5333. [Google Scholar] [CrossRef]
- Becker, A.; Theissen, G. The major clades of MADS-box genes and their role in the development and evolution of flowering plants. Mol. Phylogenetics Evol. 2003, 29, 464–489. [Google Scholar] [CrossRef]
- Yu, L.H.; Wu, J.; Zhang, Z.S.; Miao, Z.Q.; Zhao, P.X.; Wang, Z.; Xiang, C.B. Arabidopsis MADS-box transcription factor AGL21 acts as environmental surveillance of seed germination by regulating ABI5 expression. Mol. Plant 2017, 10, 834–845. [Google Scholar] [CrossRef]
- Sun, C.H.; Yu, J.Q.; Duan, X.; Wang, J.H.; Zhang, Q.Y.; Gu, K.D.; Hu, D.G.; Zheng, C.S. The MADS transcription factor CmANR1 positively modulates root system development by directly regulating CmPIN2 in chrysanthemum. Hortic. Res. 2018, 5, 52. [Google Scholar] [CrossRef] [PubMed]
- Saedler, H.; Becker, A.; Winter, K.U.; Kirchner, C.; Theissen, G. MADS-box genes are involved in floral development and evolution. Acta Biochim. Pol. 2001, 48, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Pnueli, L.; Hareven, D.; Rounsley, S.D.; Yanofsky, M.F.; Lifschitz, E. Isolation of the tomato AGAMOUS gene TAG1 and analysis of its homeotic role in transgenic plants. Plant Cell 1994, 6, 163–173. [Google Scholar] [PubMed]
- Pan, I.L.; McQuinn, R.; Giovannoni, J.J.; Irish, V.F. Functional diversification of AGAMOUS lineage genes in regulating tomato flower and fruit development. J. Exp. Bot. 2010, 61, 1795–1806. [Google Scholar] [CrossRef]
- Itkin, M.; Seybold, H.; Breitel, D.; Rogachev, I.; Meir, S.; Aharoni, A. TOMATO AGAMOUS-LIKE 1 is a component of the fruit ripening regulatory network. Plant J. 2009, 60, 1081–1095. [Google Scholar] [CrossRef]
- Vrebalov, J.; Pan, I.L.; Arroyo, A.J.M.; McQuinn, R.; Chung, M.; Poole, M.; Rose, J.; Seymour, G.; Grandillo, S.; Giovannoni, J.; et al. Fleshy fruit expansion and ripening are regulated by the Tomato SHATTERPROOF gene TAGL1. Plant Cell 2009, 21, 3041–3062. [Google Scholar] [CrossRef]
- Giménez, E.; Pineda, B.; Capel, J.; Antón, M.T.; Atarés, A.; Pérez-Martín, F.; García-Sogo, B.; Angosto, T.; Moreno, V.; Lozano, R. Functional analysis of the Arlequin mutant corroborates the essential role of the Arlequin/TAGL1 gene during reproductive development of tomato. PLoS ONE 2010, 5, e14427. [Google Scholar] [CrossRef] [PubMed]
- Busi, M.V.; Bustamante, C.; D’angelo, C.; Hidalgo-Cuevas, M.; Boggio, S.B.; Valle, E.M.; Zabaleta, E. MADS-box genes expressed during tomato seed and fruit development. Plant Mol. Biol. 2003, 52, 801–815. [Google Scholar]
- Seymour, G.B.; Manning, K.; Eriksson, E.M.; Popovich, A.H.; King, G.J. Genetic identification and genomic organization of factors affecting fruit texture. J. Exp. Bot. 2002, 53, 2065–2071. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, M.; Shima, Y.; Higuchi, N.; Nakano, T.; Koyama, Y.; Kasumi, T.; Ito, Y. Direct targets of the tomato-ripening regulator RIN identified by transcriptome and chromatin immunoprecipitation analyses. Planta 2012, 235, 1107–1122. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef]
- Schilling, S.; Kennedy, A.; Pan, S.; Jermiin, L.S.; Melzer, R. Genome-wide analysis of MIKC-type MADS-box genes in wheat: Pervasive duplications, functional conservation and putative neofunctionalization. New Phytol. 2020, 225, 511–529. [Google Scholar] [CrossRef]
- Kaufmann, K.; Melzer, R.; Theissen, G. MIKC-type MADS-domain proteins: Structural modularity, protein interactions and network evolution in land plants. Gene 2005, 347, 183–198. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Wang, M.; Meyerowitz, E.M. DNA-binding properties of Arabidopsis MADS domain homeotic proteins APETALA1, APETALA3, PISTILLATA and AGAMOUS. Nucleic Acids Res. 1996, 24, 3134–3141. [Google Scholar] [CrossRef]
- Immink, R.G.; Gadella, T.W., Jr.; Ferrario, S.; Busscher, M.; Angenent, G.C. Analysis of MADS box protein-protein interactions in living plant cells. Proc. Natl. Acad. Sci. USA 2002, 99, 2416–2421. [Google Scholar] [CrossRef]
- Yang, Y.; Jack, T. Defining subdomains of the K domain important for protein-protein interactions of plant MADS proteins. Plant Mol. Biol. 2004, 55, 45–59. [Google Scholar] [CrossRef]
- Melzer, R.; Verelst, W.; Theissen, G. The class E floral homeotic protein SEPALLATA3 is sufficient to loop DNA in ‘floral quartet’-like complexes in vitro. Nucleic Acids Res. 2009, 37, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Theißen, G.; Melzer, R.; Rümpler, F. MADS-domain transcription factors and the floral quartet model of flower development: Linking plant development and evolution. Development 2016, 143, 3259–3271. [Google Scholar] [CrossRef]
- Honma, T.; Goto, K. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs. Nature 2001, 409, 525–529. [Google Scholar] [CrossRef]
- Gramzow, L.; Theissen, G. A hitchhiker’s guide to the MADS world of plants. Genome Biol. 2010, 11, 214. [Google Scholar] [CrossRef]
- Arora, R.; Agarwal, P.; Ray, S.; Singh, A.K.; Singh, V.P.; Tyagi, A.K.; Kapoor, S. MADS-box gene family in rice: Genome-wide identification, organization and expression profiling during reproductive development and stress. BMC Genom. 2007, 8, 242. [Google Scholar] [CrossRef]
- Jia, J.; Zhao, P.; Cheng, L.; Yuan, G.; Yang, W.; Liu, S.; Chen, S.; Qi, D.; Liu, G.; Li, X. MADS-box family genes in sheepgrass and their involvement in abiotic stress responses. BMC Plant Biol. 2018, 18, 42. [Google Scholar] [CrossRef]
- Wei, M.; Wang, Y.; Pan, R.; Li, W. Genome-wide identification and characterization of MADS-box family genes related to floral organ development and stress resistance in Hevea brasiliensis Müll. Arg. Forests 2018, 9, 304. [Google Scholar] [CrossRef]
- Dong, T.; Hu, Z.; Deng, L.; Wang, Y.; Zhu, M.; Zhang, J.; Chen, G. A tomato MADS-box transcription factor, SlMADS1, acts as a negative regulator of fruit ripening. Plant Physiol. 2013, 163, 1026–1036. [Google Scholar] [CrossRef]
- Lu, W.; Chen, J.; Ren, X.; Yuan, J.; Han, X.; Mao, L.; Ying, T.; Luo, Z. One novel strawberry MADS-box transcription factor FaMADS1a acts as a negative regulator in fruit ripening. Sci. Hortic. 2018, 227, 124–131. [Google Scholar] [CrossRef]
- Elitzur, T.; Yakir, E.; Quansah, L.; Zhangjun, F.; Vrebalov, J.; Khayat, E.; Giovannoni, J.J.; Friedman, H. Banana MaMADS transcription factors are necessary for fruit ripening and molecular tools to promote shelf-life and food security. Plant Physiol. 2016, 171, 380–391. [Google Scholar] [PubMed]
- Qi, X.L.; Liu, C.L.; Song, L.L.; Li, M. PaMADS7, a MADS-box transcription factor, regulates sweet cherry fruit ripening and softening. Plant Sci. 2020, 301, 110634. [Google Scholar] [CrossRef]
- Yin, W.; Yu, X.; Chen, G.; Tang, B.; Wang, Y.; Liao, C.; Zhang, Y.; Hu, Z. Suppression of SlMBP15 inhibits plant vegetative growth and delays fruit ripening in tomato. Front. Plant Sci. 2018, 9, 938. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.R.; Cui, J.; He, Q.; Liu, Y.Z.; Lu, X.F.; Qi, J.; Xiong, J.; Yu, W.J.; Li, C.X. Genome-wide identification of HIPP and mechanism of SlHIPP4/7/9/21/26/32 mediated phytohormones response to Cd, osmotic, and salt stresses in tomato. Plant Physiol. 2024, 217, 109220. [Google Scholar] [CrossRef]
- Bi, Y.; Fu, H.; Jiang, Z.; Jiang, Y.; You, L.; Li, C.; Tu, X.; Ahmad, S.; Liu, Z.; Chen, S.; et al. Integrating Genome and Transcriptome-Wide Data to Explore the Expression Dynamics of ABCDE-like MADS-Box Genes in Phoebe bournei Floral Organs. Forests 2025, 16, 313. [Google Scholar] [CrossRef]
- Cui, J.; Xu, J.R.; Qi, J.; Lu, X.F.; Liu, Y.Z.; Xiong, J.; Yu, W.J.; Li, C.X. Genome-wide identification of SlIQMs and the regulatory effect of calcium on tomato seedlings under drought stress and phytohormone treatment. Plant Cell 2025, 44, 70. [Google Scholar]
- Zhou, E.; Zhang, Y.; Wang, H.; Jia, Z.; Wang, X.; Wen, J.; Shen, J.; Fu, T.; Yi, B. Identification and characterization of the MIKC-Type MADS-Box gene family in Brassica napus and its role in floral transition. Int. J. Mol. Sci. 2022, 23, 4289. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Hu, Z.; Guo, X.; Tian, S.; Chen, G. Genome-Wide analysis of the MADS-Box transcription factor family in Solanum lycopersicum. Int. J. Mol. Sci. 2019, 20, 2961. [Google Scholar] [CrossRef] [PubMed]
- Parenicová, L.; de Folter, S.; Kieffer, M.; Horner, D.S.; Favalli, C.; Busscher, J.; Cook, H.E.; Ingram, R.M.; Kater, M.M.; Davies, B.; et al. Molecular and phylogenetic analyses of the complete MADS-box transcription factor family in Arabidopsis: New openings to the MADS world. Plant Cell 2003, 15, 1538–1551. [Google Scholar] [CrossRef]
- Ray, S.; Agarwal, P.; Arora, R.; Kapoor, S.; Tyagi, A.K. Expression analysis of calcium-dependent protein kinase gene family during reproductive development and abiotic stress conditions in rice (Oryza sativa L. Ssp. Indica). Mol. Genet. Genom. 2007, 278, 493–505. [Google Scholar] [CrossRef]
- Qu, Y.; Kong, W.; Wang, Q.; Fu, X. Genome-wide identification MIKC-Type MADS-Box gene family and their roles during development of floral buds in wheel wingnut (Cyclocarya paliurus). Int. J. Mol. Sci. 2021, 22, 10128. [Google Scholar] [CrossRef]
- Chen, S.P.; Sun, W.H.; Xiong, Y.F.; Jiang, Y.T.; Liu, X.D.; Liao, X.Y.; Zhang, D.Y.; Jiang, S.Z.; Li, Y.; Liu, B.; et al. The Phoebe genome sheds light on the evolution of magnoliids. Hortic. Res. 2020, 7, 146. [Google Scholar] [CrossRef]
- Chen, T.T.; Liu, H.; Li, Y.P.; Yao, X.H.; Qin, W.; Yan, X.; Wang, X.Y.; Peng, B.W.; Zhang, Y.J.; Shao, J.; et al. AaSEPALLATA1 integrates jasmonate and light-regulated glandular secretory trichome initiation in Artemisia annua. Plant Physiol. 2023, 192, 1483–1497. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Yuan, C.; Sun, Q.; Yan, C.; Zhao, X.; Wang, J.; Wang, Q.; Shan, S.; Li, C. MIKC-type MADS-box transcription factor gene family in peanut: Genome-wide characterization and expression analysis under abiotic stress. Front. Plant Sci. 2022, 13, 980933. [Google Scholar] [CrossRef] [PubMed]
- Martel, C.; Vrebalov, J.; Tafelmeyer, P.; Giovannoni, J.J. The tomato MADS-box transcription factor RIPENING INHIBITOR interacts with promoters involved in numerous ripening processes in a COLORLESS NONRIPENING-dependent manner. Plant Physiol. 2011, 157, 1568–1579. [Google Scholar] [CrossRef]
- Bemer, M.; Karlova, R.; Ballester, A.R.; Tikunov, Y.M.; Bovy, A.G.; Wolters-Arts, M.; de Barros Rossetto, P.; Angenent, G.C.; de Maagd, R.A. The tomato FRUITFULL homologs TDR4/FUL1 and MBP7/FUL2 regulate ethylene-independent aspects of fruit ripening. The Plant Cell 2012, 24, 4437–4451. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.D.; Yuan, X.; Chen, S.; Fu, D.Q.; Jiang, C.Z. Metabolomic and transcriptomic analyses reveal that a MADS-Box transcription factor TDR4 regulates tomato fruit quality. Front. Plant Sci. 2019, 10, 792. [Google Scholar] [CrossRef]
- Gimenez, E.; Castañeda, L.; Pineda, B.; Pan, I.L.; Moreno, V.; Angosto, T.; Lozano, R. TOMATO AGAMOUS1 and ARLEQUIN/TOMATO AGAMOUS-LIKE1 MADS-box genes have redundant and divergent functions required for tomato reproductive development. Plant Mol. Biol. 2016, 91, 513–531. [Google Scholar] [CrossRef]
- Nakano, T.; Kimbara, J.; Fujisawa, M.; Kitagawa, M.; Ihashi, N.; Maeda, H.; Kasumi, T.; Ito, Y. MACROCALYX and JOINTLESS interact in the transcriptional regulation of tomato fruit abscission zone development. Plant Physiol. 2012, 58, 439–450. [Google Scholar] [CrossRef]
- Zheng, S.; He, J.J.; Lin, Z.S.; Zhu, Y.Y.; Sun, J.Y.; Li, L.G. Two MADS-box genes regulate vascular cambium activity and secondary growth by modulating auxin homeostasis in Populus. Plant Commun. 2020, 2, 100134. [Google Scholar] [CrossRef]
- Li, C.H.; Lei, C.Y.; Wang, K.T.; Tan, M.L.; Xu, F.; Wang, J.S.; Zheng, Y.H. MADS2 regulates priming defence in postharvest peach through combined salicylic acid and abscisic acid signaling. J. Exp. Bot. 2022, 73, 3787–3806. [Google Scholar] [CrossRef]
- Kou, X.; Zhao, Y.; Wu, C.; Jiang, B.; Zhang, Z.; Rathbun, J.R.; He, Y.; Xue, Z. SNAC4 and SNAC9 transcription factors show contrasting effects on tomato carotenoids biosynthesis and softening. Postharvest Biol. Technol. 2018, 144, 9–19. [Google Scholar] [CrossRef]
- Negi, S.; Bhakta, S.; Ganapathi, T.R.; Tak, H. MpSNAC67 transcription factor of banana regulates stress induced senescence through salicylic acid dependent pathway. Environ. Exp. Bot. 2023, 205, 105104. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Cis-Element | Sequence of Cis-Element | Functions of Cis-Element |
|---|---|---|---|
| TM4/TDR4/LeFUL1 | TGA element | AACGAC | Auxin-responsive element |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| TATC box | TATCCCA | Cis-acting element involved in gibberellin responsiveness | |
| TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness | |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| GARE motif | TCTGTTG | Gibberellin-responsive element | |
| AuxRE | TGTCTCAATAAG | Part of an auxin-responsive element | |
| SlMBP22 | TATC box | TATCCCA | Cis-acting element involved in gibberellin responsiveness |
| TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness | |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| ABRE | CACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| P-box | CCTTTTG | Gibberellin-responsive element | |
| SlMADS1 | TATC box | TATCCCA | Cis-acting element involved in gibberellin responsiveness |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| ABRE | GACACGTGGC | Cis-acting element involved in abscisic acid responsiveness | |
| ABRE | CACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| ABRE | CGCACGTGTC | Cis-acting element involved in abscisic acid responsiveness | |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| SlMADS6/TM29/LeSEP1 | TATC box | TATCCCA | Cis-acting element involved in gibberellin responsiveness |
| TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness | |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| AuxRR core | GGTCCAT | Cis-acting regulatory element involved in auxin responsiveness | |
| GARE motif | TCTGTTG | Gibberellin-responsive element | |
| SlMADS84 | TATC box | TATCCCA | Cis-acting element involved in gibberellin responsiveness |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| ABRE | CACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| GARE motif | TCTGTTG | Gibberellin-responsive element | |
| TAP3/LeAP3/LeDEF | TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| GARE motif | TCTGTTG | Gibberellin-responsive element | |
| SlMADS99 | GARE motif | TCTGTTG | Gibberellin-responsive element |
| MADS-RIN/LeAP1 | TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| SlMBP20 | ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGA box | TGACGTAA | Part of an auxin-responsive element | |
| TM8/TDR8 | TGA element | AACGAC | Auxin-responsive element |
| TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness | |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGA box | TGACGTAA | Part of an auxin-responsive element | |
| SlMBP21 | ABRE | GCAACGTGTC | Cis-acting element involved in abscisic acid responsiveness |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| ABRE | TACGGTC | Cis-acting element involved in abscisic acid responsiveness | |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| O2 site | GTTGACGTGA | Cis-acting regulatory element involved in zein metabolism regulation | |
| P-box | CCTTTTG | Gibberellin-responsive element | |
| TGA box | TGACGTAA | Part of an auxin-responsive element | |
| TAGL1 | TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness |
| ABRE | CACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| P-box | CCTTTTG | Gibberellin-responsive element | |
| SlMADS92 | SARE | TTCGACCATCTT | Cis-acting element involved in salicylic acid responsiveness |
| TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness | |
| P-box | CCTTTTG | Gibberellin-responsive element | |
| SlMBP2/SlGLO2/LePI/TPI | TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| O2 site | GATGATGTGG | Cis-acting regulatory element involved in zein metabolism regulation | |
| P-box | CCTTTTG | Gibberellin-responsive element | |
| TAG1 | TGA element | AACGAC | Auxin-responsive element |
| TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness | |
| AuxRR core | GGTCCAT | Cis-acting regulatory element involved in auxin responsiveness | |
| P-box | CCTTTTG | Gibberellin-responsive element | |
| TAGL11 | TGA element | AACGAC | Cis-responsive element |
| TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness | |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| O2 site | GATGACATGG | Cis-acting regulatory element involved in zein metabolism regulation | |
| P-box | CCTTTTG | Gibberellin-responsive element | |
| SlMADS90 | TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| O2 site | GATGATGTGG | Cis-acting regulatory element involved in zein metabolism regulation | |
| SlMBP1/SlGLO1/PI/LePI-B | ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness |
| ABRE | CGTACGTGCA | Cis-acting element involved in abscisic acid responsiveness | |
| O2 site | GATGA(C/T)(A/G)TG(A/G) | Cis-acting regulatory element involved in zein metabolism regulation | |
| MADS-MC | TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| O2 site | GTTGACGTGA | Cis-acting regulatory element involved in zein metabolism regulation | |
| P-box | CCTTTTG | Gibberellin-responsive element | |
| SlMADS88 | TCA element | CCATCTTTTT | Cis-acting element involved in salicylic acid responsiveness |
| ABRE | ACGTG | Cis-acting element involved in abscisic acid responsiveness | |
| CGTCA motif | CGTCA | Cis-acting regulatory element involved in MeJA responsiveness | |
| TGACG motif | TGACG | Cis-acting regulatory element involved in MeJA responsiveness | |
| P-box | CCTTTTG | Gibberellin-responsive element |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Lu, Y.; Xu, J.; Cui, J.; Liu, Y.; Yu, W. MIKC-Type MADS-box Genes Regulate Phytohormone-Dependent Fruit Ripening in Tomatoes. Horticulturae 2025, 11, 487. https://doi.org/10.3390/horticulturae11050487
Li C, Lu Y, Xu J, Cui J, Liu Y, Yu W. MIKC-Type MADS-box Genes Regulate Phytohormone-Dependent Fruit Ripening in Tomatoes. Horticulturae. 2025; 11(5):487. https://doi.org/10.3390/horticulturae11050487
Chicago/Turabian StyleLi, Changxia, Yushi Lu, Junrong Xu, Jing Cui, Yunzhi Liu, and Wenjin Yu. 2025. "MIKC-Type MADS-box Genes Regulate Phytohormone-Dependent Fruit Ripening in Tomatoes" Horticulturae 11, no. 5: 487. https://doi.org/10.3390/horticulturae11050487
APA StyleLi, C., Lu, Y., Xu, J., Cui, J., Liu, Y., & Yu, W. (2025). MIKC-Type MADS-box Genes Regulate Phytohormone-Dependent Fruit Ripening in Tomatoes. Horticulturae, 11(5), 487. https://doi.org/10.3390/horticulturae11050487