
Interaction Effects of Cucumber Varieties and Pruning Methods Across Different Growth Stages

 ,
,
Abstract
1. Introduction
2. Materials and Methods
3. Results
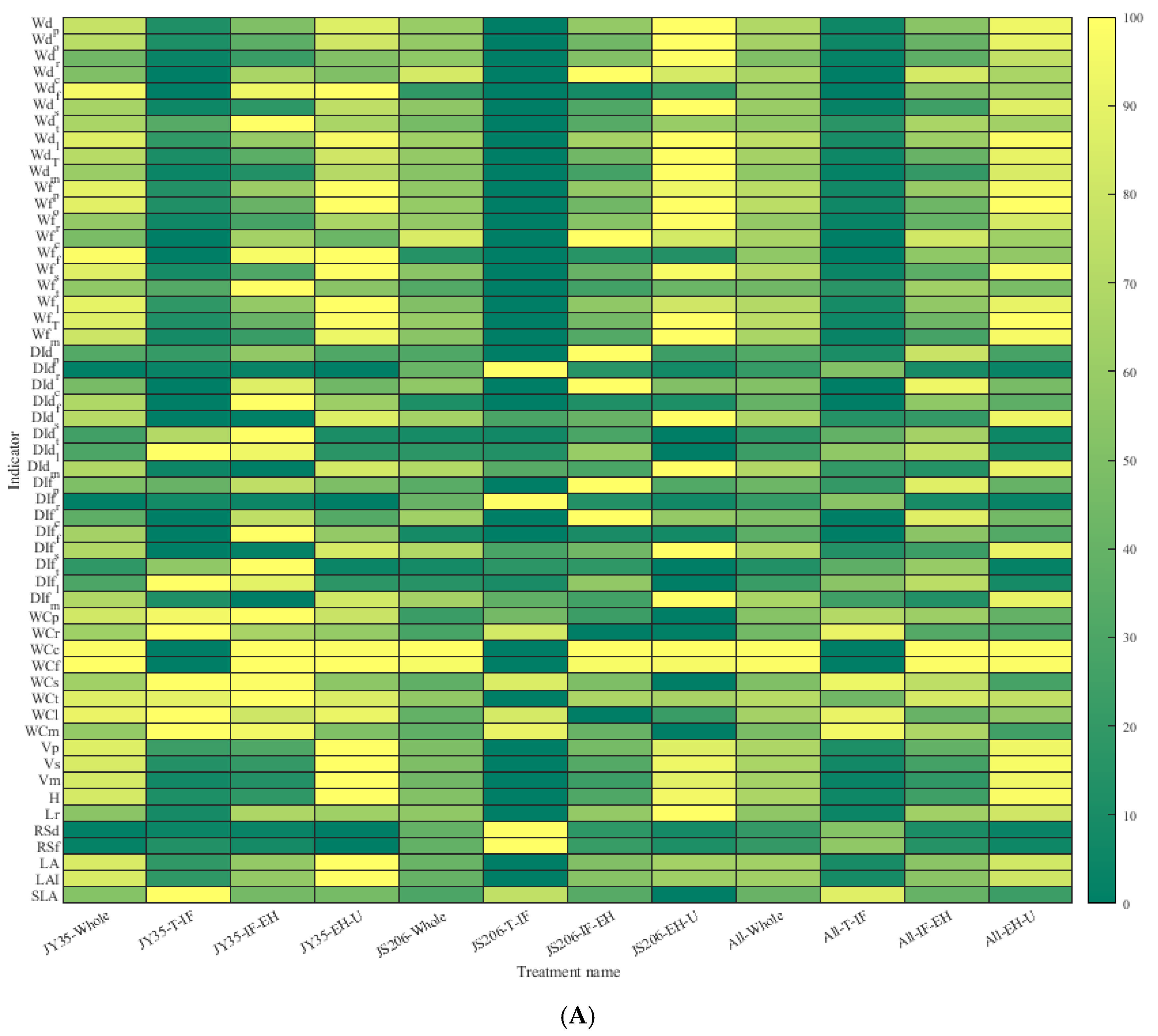
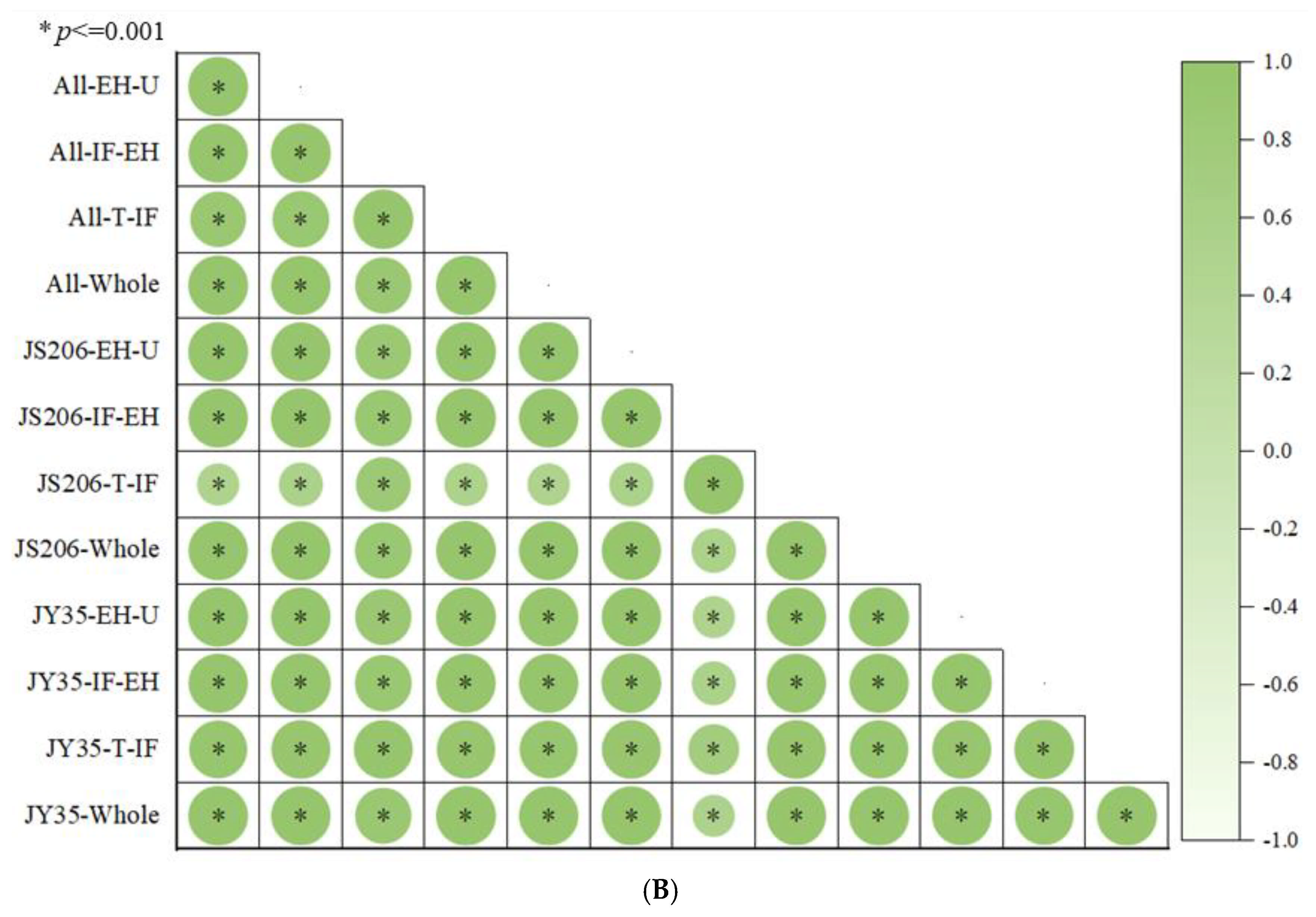
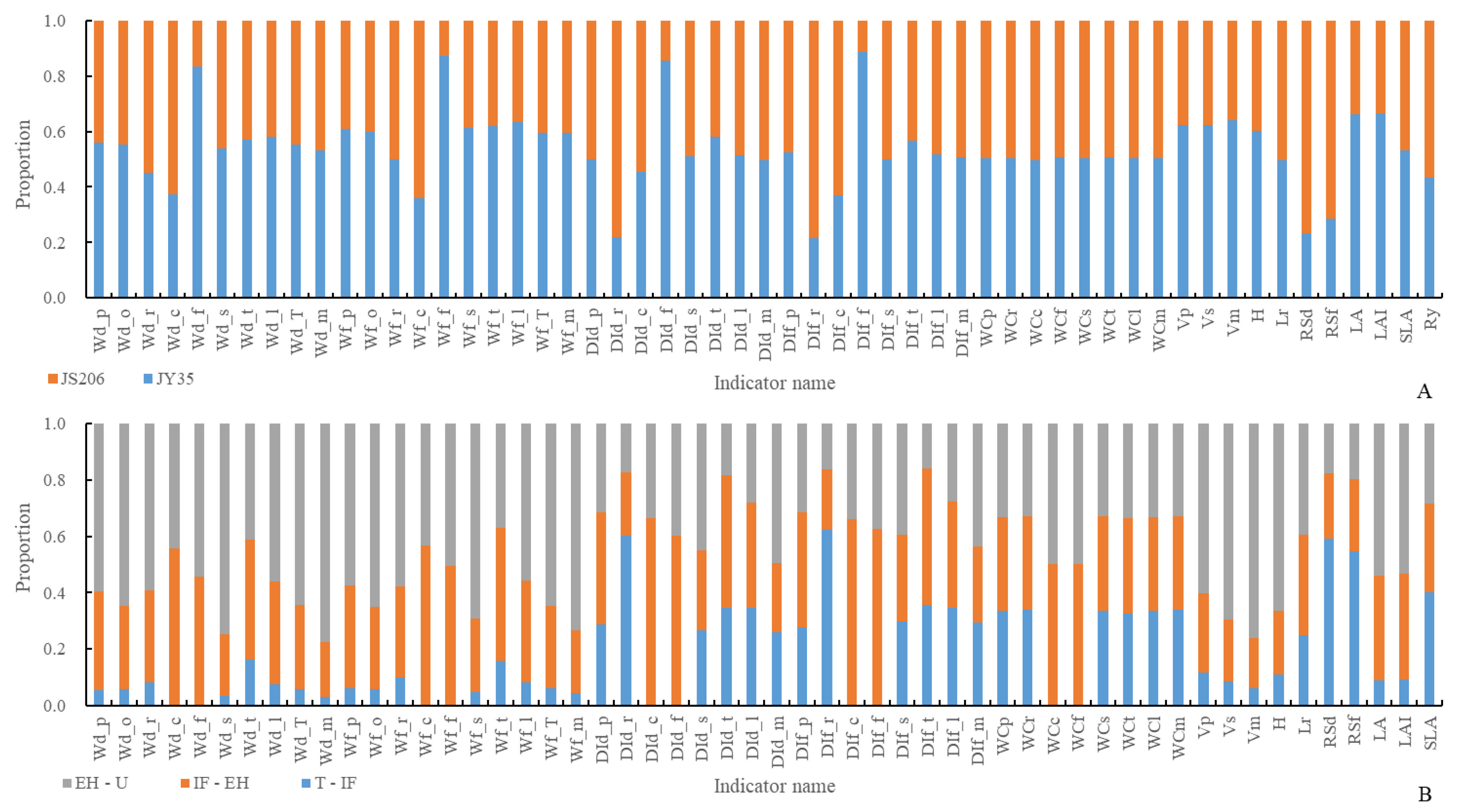
3.1. Analysis of Plant Architecture and Productivity Characteristics at Different Development Stages of Different Varieties
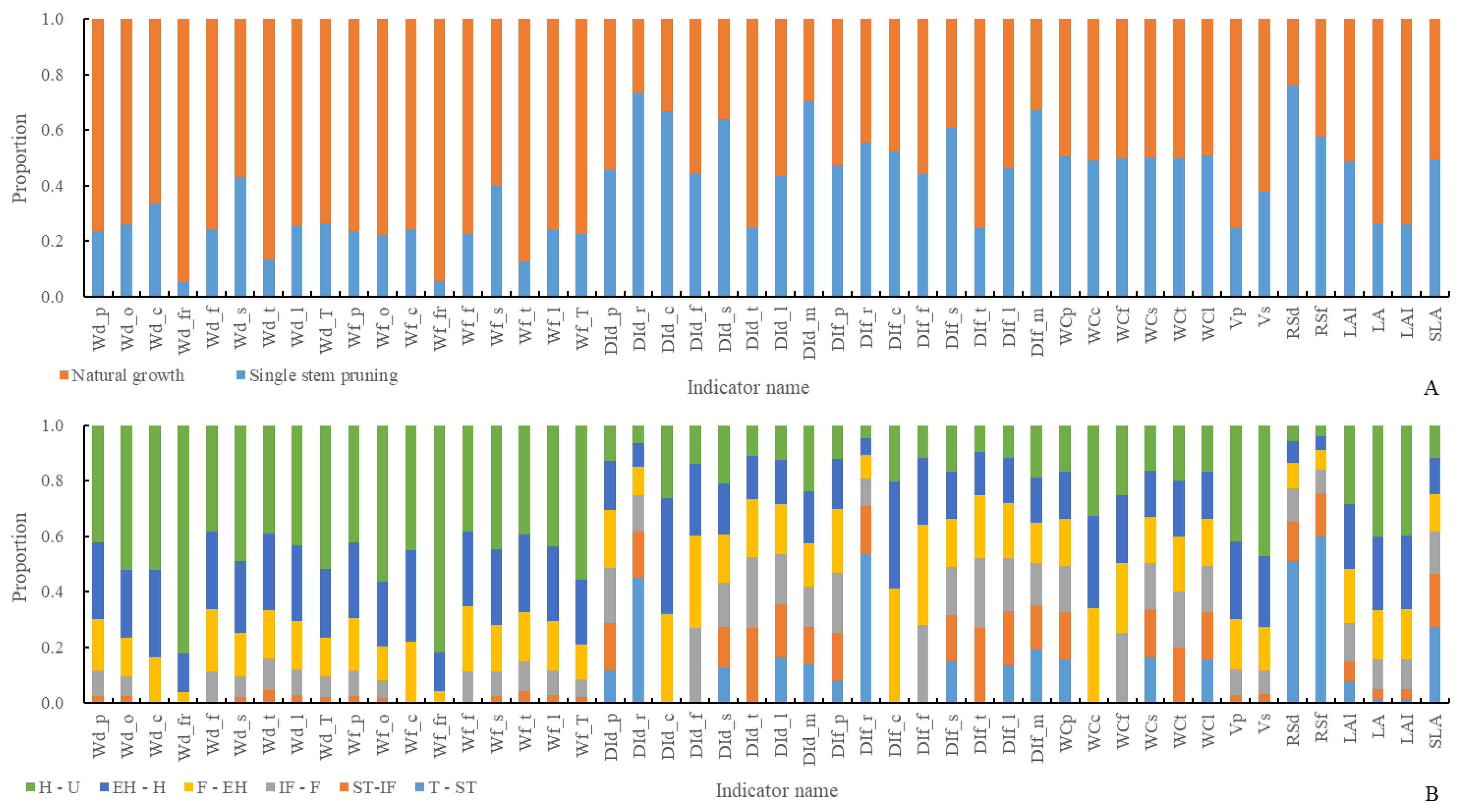
3.2. Analysis of Plant Architecture and Productivity Characteristics at Different Development Stages Under Different Pruning Methods
4. Discussion
4.1. The Impact Mechanism of Crop Varieties on Plant Plasticity
4.2. Dynamic Regulation Rules of Developmental Stages
4.3. The Impact Mechanism of Pruning Method on Plant Plasticity
5. Conclusions
- (1)
- Varieties have a significant impact on the plant architecture characteristics and productivity indicators of JY35 and JS206. The dominant plant architecture characteristics and productivity indicators of JY35 include dry and fresh weights of the tendril, main stem, total stem, leaves, petioles, flowers, overground parts, and overall plant, as well as dry and fresh weight distribution index of the tendril, total stem, leaves, petioles, flowers, overground parts, and overall plant, main stem fresh weight distribution index, water content of roots, tendrils, main stem, leaves, petioles, and flowers, volume of total stem, main stem, and petioles, plant height, total leaf area per plant, leaf area index, and specific leaf area. The remaining plant architecture characteristics and productivity indicators are dominated by the plant architecture of JS206;
- (2)
- Pruning methods have a significant impact on the plant architecture characteristics and productivity indicators of single-stem pruning (SP) and natural growth (NG). The dominant plant architecture characteristics and productivity indicators of the SP method include dry and fresh weight distribution index of roots, fruit carpopodiums, main stems, and total stems, water content of petioles, stems, and leaves, and root-to-shoot ratio. The remaining plant architecture characteristics and productivity indicators are dominated by the NG method.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cheng, C.; Feng, L.P.; Dong, C.Y.; Chen, X.G.; Yang, F.Y.; Wu, L.; Yang, J.; Zhao, C.S.; Yuan, G.Y.; Li, Z.F. Vegetable commodity organ quality formation simulation model (VQSM) in solar greenhouses. Agriculture 2024, 14, 1531. [Google Scholar] [CrossRef]
- Li, A.L.; Hao, C.Y.; Wang, Z.Y. Wheat breeding history reveals synergistic selection of pleiotropic genomic sites for plant architecture and grain yield. Mol. Plant 2022, 15, 504–519. [Google Scholar] [CrossRef]
- Cheng, C.; Dong, C.Y.; Guan, X.L.; Chen, X.G.; Wu, L.; Zhu, Y.C.; Zhang, L.; Ding, F.H.; Feng, L.P.; Li, Z.F. CPSM: A dynamic simulation model for cucumber productivity in solar greenhouse based on the principle of effective accumulated temperature. Agronomy 2024, 14, 1242. [Google Scholar] [CrossRef]
- Zhan, A.; Liu, J.L.; Yue, S.C.; Chen, X.P.; Li, S.Q.; Bucksch, A. Architectural and anatomical responses of maize roots to agronomic practices in a semi-arid environment. J. Plant Nutr. Soil Sci. 2019, 182, 751–762. [Google Scholar] [CrossRef]
- Tang, L.Y.; Yin, D.; Chen, C.C.; Yu, D.Y.; Han, W. Optimal design of plant canopy based on light interception: A case study with loquat. Front. Plant Sci. 2019, 10, 364. [Google Scholar] [CrossRef]
- Li, P.F.; Ma, B.L.; Yan, W.K.; Cheng, Z.G.; Li, F.M.; Xiong, Y.C. Plant architecture, plasticity, and adaptation strategies of two oat genotypes under different competition intensities. J. Sci. Food Agric. 2016, 96, 1431–1439. [Google Scholar] [CrossRef]
- Cheng, C.; Wu, L.; Yu, W.D.; Yang, F.Y.; Feng, L.P. Dynamic evaluation of winter wheat’s freezing resistance under different low-temperature periods and durations. Sci. Rep. 2025, 15, 8488. [Google Scholar] [CrossRef]
- Cheng, C.; Feng, L.P.; Barcena, J.F.B.; Yu, W.D.; Li, G.; Li, Z.F.; Ye, C.H. A growth model based on standardized growing degree days for hydroponic fresh cut tulip in solar greenhouses. Eur. J. Hortic. Sci. 2022, 87, 1–13. [Google Scholar] [CrossRef]
- Li, J.X.; Liu, S.S.; Gu, Q.S. Transmission efficiency of cucumber green mottle mosaic virus via seeds, soil, pruning and irrigation water. J. Phytopathol. 2015, 164, 300–309. [Google Scholar] [CrossRef]
- Sole, R.A.; Raga, H.A.; Riwukaho, U.J.; Naisanu, J.; Ndun, A.A.; Bunyani, N.A.; Kisse, D.F. Effect of giving lamtoro leaf extract and pruning on cucumber plant production (Cucumis sativus L.). J. Biol. Trop. 2022, 22, 1370–1377. [Google Scholar] [CrossRef]
- Cheng, C.; Li, Z.F.; Dong, C.Y.; Gong, Z.H.; Feng, L.P. Simulation and validation of extinction coefficient at different positions of cucumber and celery in solar greenhouse. Trans. Chin. Soc. Agric. Eng. 2020, 36, 243–252. [Google Scholar]
- Tian, J.; Wang, C.; Xia, J.; Wu, L.S.; Xu, G.H.; Wu, W.H.; Li, D.; Qin, W.C.; Han, X.; Chen, Q.Y.; et al. Teosinte ligule allele narrows plant architecture and enhances high-density maize yields. Science 2019, 365, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Chen, L.M.; Herrera-Estrella, L.; Cao, D.; Tran, L.S.P. Altering plant architecture to improve performance and resistance. Trends Plant Sci. 2020, 25, 1154–1170. [Google Scholar] [CrossRef] [PubMed]
- Li, G.H.; Xue, L.H.; Gu, W.; Yang, C.D.; Wang, S.H.; Ling, Q.H.; Qin, X.; Ding, Y.F. Comparison of yield components and plant type characteristics of high-yield rice between Taoyuan, a ‘special eco-site’ and Nanjing, China. Field Crops Res. 2009, 112, 214–221. [Google Scholar] [CrossRef]
- Flint, J.; Eskin, E. Genome-wide association studies in mice. Nat. Rev. Genet. 2012, 13, 807–817. [Google Scholar] [CrossRef]
- Henke, M.; Kurth, W.; Buck-Sorlin, G.H. FSPM-P: Towards a general functional-structural plant model for robust and comprehensive model development. Front. Comput. Sci. 2016, 10, 1103–1117. [Google Scholar] [CrossRef]
- Abuqamar, S.; Moustafa, K.; Tran, L.S. Mechanisms and strategies of plant defense against Botrytis cinerea. Crit. Rev. Biotechnol. 2017, 37, 262–274. [Google Scholar] [CrossRef]
- Cheng, X.Y.; Xu, J.; Liu, H.; Zhu, Z.J.; Xu, Z.H. Effects of re-growth pruning of over-winter cultivated tomato plants on yield and nutrient uptake. Acta Agric. Zhejiangensis 2005, 17, 37–40. [Google Scholar]
- Chen, J.F.; Cui, L.H.; Tian, Q.J.; Liu, S.B. Effects of two different pruning methods on growth, development and internal quality of siraitia grosvenorii. Chin. J. Trop. Crops 2013, 34, 1435–1438. [Google Scholar]
- Wang, H.R.; Yan, S.H.; Gao, Y.M.; Li, J.S. Effects of different pruning patterns on fruit commodity, nutritional quality and yield of cherry tomato. J. Zhejiang Univ. 2021, 47, 347–353. [Google Scholar]
- Wang, H.R.; Li, J.S.; Yan, S.H.; Gao, Y.M. Effect of prunning patterns on canopy light interception characteristics and chlorophyll fluorescence parameters in cherry tomato. Acta Agric. Zhejiangensis 2021, 47, 347–353. [Google Scholar]
- Lin, H.X.; Yuan, Z.Q.; Zhang, Z.H.; Xiao, Y.P.; Wang, R.Q.; Lv, F.J. Effects of foliage removal on tuber yield and NPK accumulation and distribution in different cassava types and cultivars. J. Plant Nutr. Fertil. 2021, 27, 1829–1848. [Google Scholar]
- Song, M.F.; Zha, G.H.; Chen, J.F.; Lou, Q.F. Research progress on molecular basis of plant architecture related traits in cucumber. Acta Hortic. Sin. 2022, 49, 2683–2702. [Google Scholar]
- Ezzat, A.O.; Tawfeek, A.M.; Mohammad, F.; Al-Lohedan, H.A. Modification of magnetite nanoparticles surface with multifunctional ionic liquids for coomassie brilliant blue R-250 dye removal from aqueous solutions. J. Mol. Liq. 2022, 358, 119195. [Google Scholar] [CrossRef]
- Li, Y.; Liang, G.; Lu, S.; Wang, H.; Zeng, F.W.; Nai, G.J.; Mao, J.; Chen, B.H. Overexpression of VaSS4 negatively regulates cold tolerance by disturbing ROS balance and decreasing soluble sugar content. Sci. Hortic. 2024, 337, 113590. [Google Scholar] [CrossRef]
- Li, X.Y.; Meng, L.; Shen, L.; Ji, H.F. Regulation of gut microbiota by vitamin C, vitamin E and β-carotene. Food Res. Int. 2023, 169, 112749. [Google Scholar] [CrossRef]
- Zhao, C.B.; Song, S.R.; Zhao, J.; Zhang, X.M.; Zhang, Y.; Zhang, S.T. Variation in nitrogen uptake and utilization efficiency of different cucumber varieties in Northern China. Sci. Agric. Sin. 2015, 48, 1569–1578. [Google Scholar]
- Wang, D.D.; Zhang, Q.Y.; Li, Y.; Qi, L.F.; Niu, R.S.; Shi, J.H. Evaluation of agronomic characters of different cucumber varieties. North. Hortic. 2022, 5, 35–40. [Google Scholar]
- Cheng, C.; Feng, L.P.; Xue, Q.Y.; Li, C.; Gong, Z.H.; Dong, C.Y.; Wu, L.; Wang, C.L.; Liu, S.M.; Li, Y.Z.; et al. Simulation model for cucumber growth and development in sunlight greenhouse. Chin. J. Appl. Ecol. 2019, 30, 3491–3500. [Google Scholar]
- Cheng, C.; Li, C.; Li, W.M.; Ye, C.Y.; Wang, Y.S.; Zhao, C.S.; Ding, F.H.; Jin, Z.F.; Feng, L.P.; Li, Z.F. Optimal path of the simulation model in horticultural crop development and harvest period. Trans. Chin. Soc. Agric. Eng. 2023, 39, 158–167. [Google Scholar]
- Cheng, C.; Dong, C.Y.; Li, Z.F.; Gong, Z.H.; Feng, L.P. Simulation model of external morphology and dry matter accumulation and distribution of celery in solar greenhouse. Trans. Chin. Soc. Agric. Eng. 2021, 37, 142–151. [Google Scholar]
- Li, Y.X.; Luo, W.H.; Ni, J.H.; Chen, Y.S.; Xu, G.B.; Jin, L.; Dai, J.F.; Chen, C.H. Simulation of leaf area, photosynthetic rate and dry matter production in greenhouse cucumber based on product of thermal effectiveness and photosynthetically active radiation. Trans. Chin. Soc. Agric. Eng. 2005, 21, 131–136. [Google Scholar]
- Tan, W.; Yang, Z.Q.; Li, J. Simulation of nutrient quality of pakchoi based on temperature-light function. Chin. J. Agrometeorol. 2016, 37, 59–67. [Google Scholar]
- Zhao, G.; Siebert, S.; Enders, A.; Rezaei, E.E.; Yan, C.Q.; Ewert, F. Demand for multi-scale weather data for regional crop modeling. Agric. For. Meteorol. 2015, 200, 156–171. [Google Scholar] [CrossRef]
- Zhu, Y.; Tang, L.; Liu, L.L.; Liu, B.; Zhang, X.H.; Qiu, X.L.; Tian, Y.C.; Cao, W.X. Research progress on the crop growth model (CropGrow). Sci. Agric. Sin. 2020, 53, 3235–3256. [Google Scholar]
- Yang, Q.; Shi, L.S.; Han, J.Y.; Zha, Y.Y.; Yu, J.; Wu, W.X.; Huang, K. Regulating the time of the crop model clock: A data assimilation framework for regions with high phenological heterogeneity. Field Crops Res. 2023, 293, 108847. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Developmental Period | Morphological Characteristics |
|---|---|
| Transplanting date (T) | The third true leaf unfolds, and the fourth true leaf appears, known as ‘Three Leaves and One Heart’ |
| Stretch Tendril period (ST) | The fourth true leaf unfolds, and the fifth true leaf appears, known as ‘four leaves and one heart’. The internode length increases, and tendrils appear at the leaf axils, changing from upright to prostrate. At the end of the seedling stage, the plant transitions from slow growth to rapid growth |
| Initial Flowering period (IF) | The first female flower blooms (in cultivation management, only the flowers above the sixth leaf are left, and the flowers below the sixth leaf are removed) |
| Fruiting period (F) | When the root fruit sits (when the fruit turns from yellow green to dark green, it means the root fruit is sitting), the plant flowers wither and grow young fruit that are 2–3 cm long |
| Early Harvest period (EH) | The root fruit is ripe, and the harvesting standard is that the fruit length is about 30 cm. The process of fruit elongation and thickening is basically completed, and it is crispy, tender, and fragrant |
| Harvest period (H) | Usually, on the 30th to 60th day of the fruiting period, plants will rapidly and continuously bloom and bear fruit |
| Uprooting period (U) | Related to seasonal production capacity, crop rotation, utilization of light and heat resources, and human factor |
| Indicator Name | Abbreviation | Unit | Observation or Calculation Method |
|---|---|---|---|
| Dry weight of petiole | g/plant | Oven drying and electronic balance weighing | |
| Dry weight of overground | g/plant | ||
| Dry weight of root | g/plant | Oven drying and electronic balance weighing | |
| Dry weight of carpopodium | g/plant | Oven drying and electronic balance weighing | |
| Dry weight of flower | g/plant | Oven drying and electronic balance weighing | |
| Dry weight of stem | g/plant | ||
| Dry weight of tendrils | g/plant | Oven drying and electronic balance weighing | |
| Dry weight of leaf | g/plant | Oven drying and electronic balance weighing | |
| Dry weight of total plant | g/plant | ||
| Dry weight of main stem | g/plant | Oven drying and electronic balance weighing | |
| Dry weight of fruit | g/plant | Oven drying and electronic balance weighing | |
| Fresh weight of petiole | g/plant | Electronic balance weighing | |
| Fresh weight of overground | g/plant | ||
| Fresh weight of root | g/plant | Electronic balance weighing | |
| Fresh weight of carpopodium | g/plant | Electronic balance weighing | |
| Fresh weight of flower | g/plant | Electronic balance weighing | |
| Fresh weight of stem | g/plant | ||
| Fresh weight of tendrils | g/plant | Electronic balance weighing | |
| Fresh weight of leaf | g/plant | Electronic balance weighing | |
| Fresh weight of total plant | g/plant | ||
| Fresh weight of main stem | g/plant | Electronic balance weighing | |
| Fresh weight of fruit | g/plant | Electronic balance weighing | |
| Dry weight distribution index of petiole | % | ||
| Dry weight distribution index of root | % | ||
| Dry weight distribution index of carpopodium | % | ||
| Dry weight distribution index of flower | % | ||
| Dry weight distribution index of stem | % | ||
| Dry weight distribution index of tendrils | % | ||
| Dry weight distribution index of leaf | % | ||
| Dry weight distribution index of main stem | % | ||
| Dry weight distribution index of fruit | % | ||
| Fresh weight distribution index of petiole | % | ||
| Fresh weight distribution index of root | % | ||
| Fresh weight distribution index of carpopodium | % | ||
| Fresh weight distribution index of flower | % | ||
| Fresh weight distribution index of stem | % | ||
| Fresh weight distribution index of tendrils | % | ||
| Fresh weight distribution index of leaf | % | ||
| Fresh weight distribution index of main stem | % | ||
| Fresh weight distribution index of fruit | % | ||
| Water content of petiole | % | ||
| Water content of root | % | ||
| Water content of carpopodium | % | ||
| Water content of flower | % | ||
| Water content of stem | % | ||
| Water content of tendrils | % | ||
| Water content of leaf | % | ||
| Water content of main stem | % | ||
| Water content of fruit | % | ||
| Length of petiole | cm | Using a ruler for measurement | |
| Length of stem | cm | Using a ruler for measurement | |
| Length of main stem | cm | Using a ruler for measurement | |
| Diameter of petiole | mm | Using a vernier caliper for measurement | |
| Diameter of stem | mm | Using a vernier caliper for measurement | |
| Diameter of main stem | mm | Using a vernier caliper for measurement | |
| Volume of petiole | cm3 | ||
| Volume of stem | cm3 | ||
| Volume of main stem | cm3 | ||
| Plant height | cm | Distance between the top of root and the top of the plant | |
| Maximum root length | cm | Distance from the bottom of root to the bottom of the stem | |
| Length of carpopodium | cm | Using a ruler for measurement | |
| Length of leaf | cm | Using a ruler for measurement | |
| Length of petiole | cm | Using a ruler for measurement | |
| Main stem internode stem length | cm | Using a ruler for measurement | |
| Width of leaf | cm | Using a ruler for measurement | |
| Diameter of petiole | mm | Using a vernier caliper for measurement | |
| Diameter of internode stem length | cm | Using a vernier caliper for measurement | |
| Dry root crown ratio | g/g | ||
| Fresh root crown ratio | g/g | ||
| Total leaf area per plant | cm2 | Using the coordinate paper method | |
| Leaf area per leaf | cm2 | Using the coordinate paper method | |
| Leaf area index | m2/m2 | , which d was the planting density | |
| Specific Leaf Area | cm2/g | ||
| Growth rate of individual plant yield | g/d | , which D was the number of days in transplanting date, and was the number of days in early harvest period related to the variety | |
| Number of fruit | — | Observation count | |
| Number of stem nodes | — | Observation count | |
| Chroma L of fruit | — | Each point was measured 3 times using the 3nh NR110 precision colorimeter | |
| Chroma a of fruit | — | Each point was measured 3 times using the 3nh NR110 precision colorimeter | |
| Chroma b of fruit | — | Each point was measured 3 times using the 3nh NR110 precision colorimeter | |
| Chroma L of leaf | — | Each point was measured 3 times using the 3nh NR110 precision colorimeter | |
| Chroma a of leaf | — | Each point was measured 3 times using the 3nh NR110 precision colorimeter | |
| Chroma b of leaf | — | Each point was measured 3 times using the 3nh NR110 precision colorimeter | |
| Hue of fruit | — | ||
| Color value of fruit | — | ||
| Darker value of fruit | — | ||
| Hue of leaf | — | ||
| Color value of leaf | — | ||
| Darker value of leaf | — | ||
| Firmness of fruit | N | GY-4 hardness tester tested at 2–3 cm from the bottom, middle, and top of fruit | |
| Length of fruit | cm | Using a ruler for measurement | |
| Radian of fruit | cm | Using a ruler for measurement | |
| Diameter of fruit | mm | Using a vernier caliper for measurement | |
| Volume of fruit | cm3 | , assuming the fruit is considered two connected cylinders | |
| Shape index of fruit | — | ||
| Fruit edibility rate | % | ||
| Chlorophyll content | SPAD | Using a handheld portable chlorophyll meter | |
| Soluble solid content | Brix% | Using PAL-1 digital refractometer [3] | |
| Soluble protein content | mg/g | Coomassie Brilliant Blue method [24] | |
| Soluble sugar content | mg/g | Anthrone Colorimetric method [25] | |
| Vitamin C | mg/kg | 2,4-dinitrophenylhydrazine method [26] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, C.; Dong, C.; Wu, L.; Wu, Y.; Wang, J.; Gong, Z.; Feng, L.; Li, Z.; Yang, F.; Zheng, S. Interaction Effects of Cucumber Varieties and Pruning Methods Across Different Growth Stages. Horticulturae 2025, 11, 464. https://doi.org/10.3390/horticulturae11050464
Cheng C, Dong C, Wu L, Wu Y, Wang J, Gong Z, Feng L, Li Z, Yang F, Zheng S. Interaction Effects of Cucumber Varieties and Pruning Methods Across Different Growth Stages. Horticulturae. 2025; 11(5):464. https://doi.org/10.3390/horticulturae11050464
Chicago/Turabian StyleCheng, Chen, Chaoyang Dong, Lu Wu, You Wu, Jiali Wang, Zhihong Gong, Liping Feng, Zhenfa Li, Feiyun Yang, and Shenghong Zheng. 2025. "Interaction Effects of Cucumber Varieties and Pruning Methods Across Different Growth Stages" Horticulturae 11, no. 5: 464. https://doi.org/10.3390/horticulturae11050464
APA StyleCheng, C., Dong, C., Wu, L., Wu, Y., Wang, J., Gong, Z., Feng, L., Li, Z., Yang, F., & Zheng, S. (2025). Interaction Effects of Cucumber Varieties and Pruning Methods Across Different Growth Stages. Horticulturae, 11(5), 464. https://doi.org/10.3390/horticulturae11050464