
Variations in the Mineral Composition of Houpoea Officinalis Flowers at Different Stages of Development


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Chemicals
2.3. Sample Digestion and Mineral Analysis
2.4. Statistical Analysis
3. Results and Discussion
3.1. Contents of Elements
3.1.1. Assessment of Macroelements
3.1.2. Assessment of Trace Elements
3.1.3. Assessment of Toxic Trace Elements
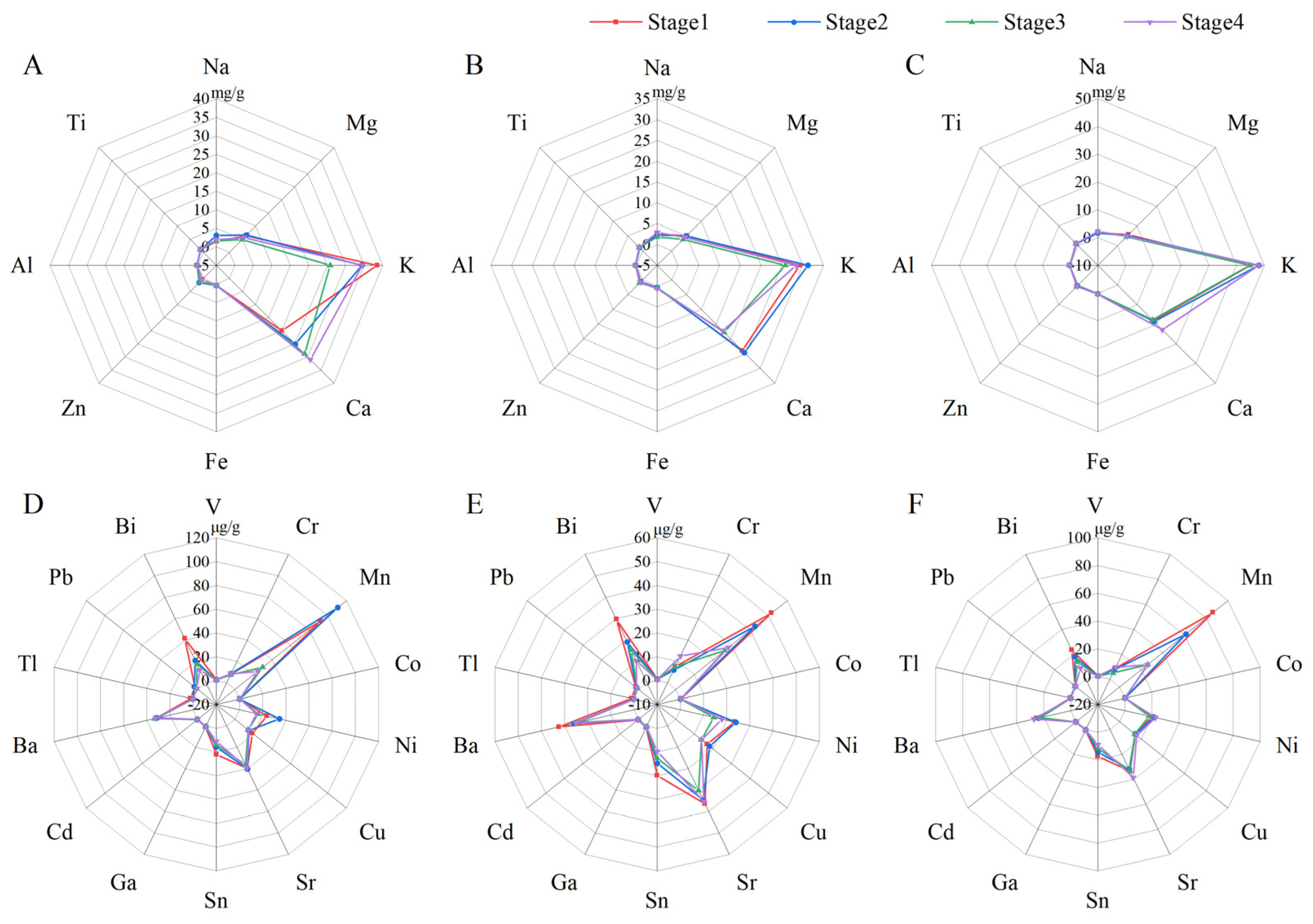
3.2. Radar Map Analysis
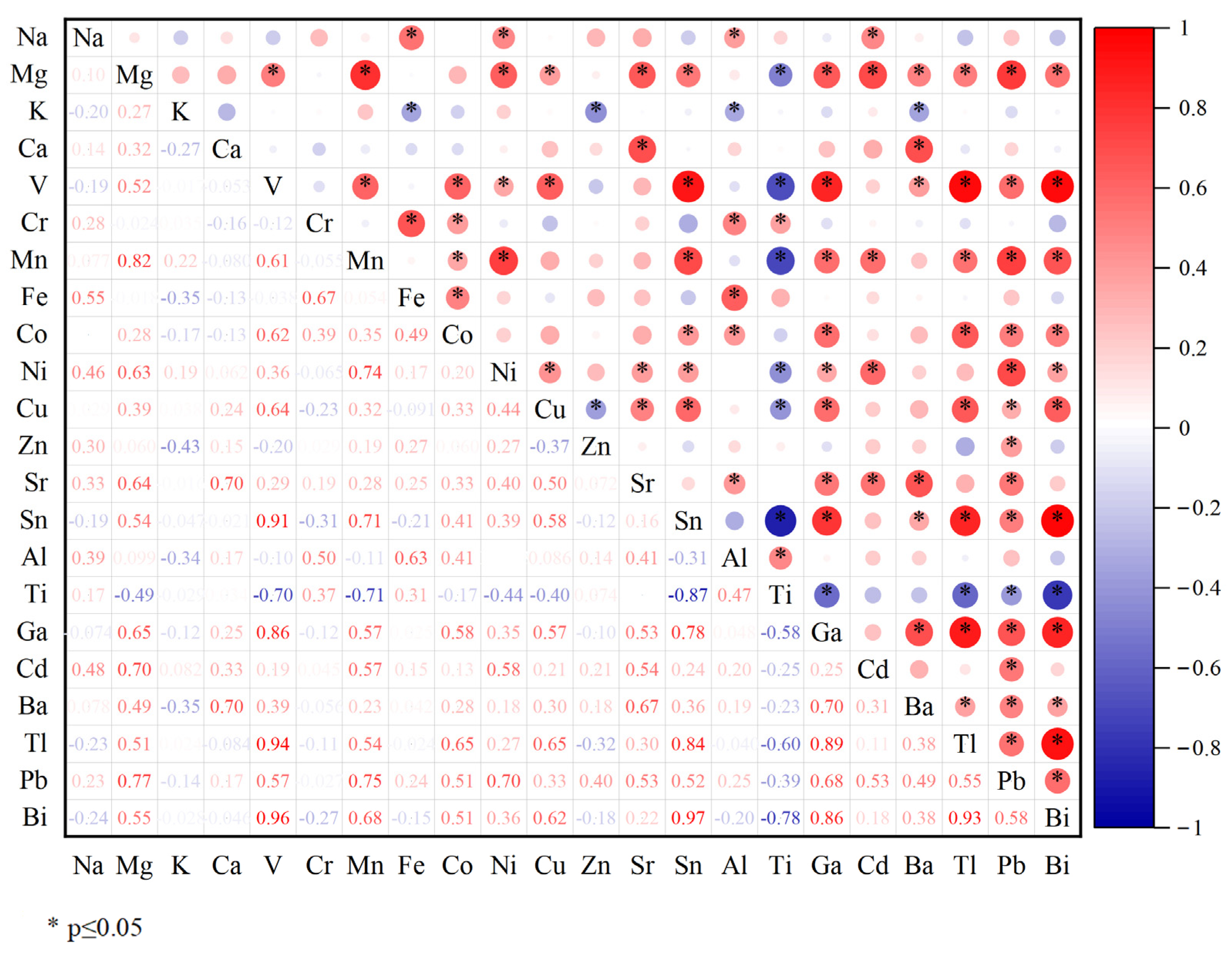
3.3. Pearson Correlation Analysis
3.4. Principal Component Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Önder, S.; Tonguç, M.; Erbaş, S.; Önder, D.; Mutlucan, M. Investigation of Phenological, Primary and Secondary Metabolites Changes During Flower Developmental of Rosa damascena. Plant Physiol. Biochem. 2022, 192, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Jhanji, S.; Kaur, G.; Kaur, R.; Dhatt, U.K. Physiological and Biochemical Changes During Flower Development and Senescence in Chrysanthemum and Gladiolus. Acta Physiol. Plant. 2023, 45, 14. [Google Scholar] [CrossRef]
- Thomson, B.; Wellmer, F. Molecular Regulation of Flower Development. Curr. Top. Dev. Biol. 2019, 131, 185–210. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.-Y.; Qian, W.-Z.; Yi, L.; Ye, Y.-L.; Gu, T.; Gao, S.; Cao, G.-X. Nutrient Composition and Antioxidant Activity of Cercis chinensis Flower in Response to Different Development Stages. Horticulturae 2023, 9, 961. [Google Scholar] [CrossRef]
- Yu, T.; Yang, Y.; Wang, H.; Qian, W.; Hu, Y.; Gao, S.; Liao, H. The Variations of C/N/P Stoichiometry, Endogenous Hormones, and Non-structural Carbohydrate Contents in Michelia maudiae ‘Rubicunda’ Flower at Five Development Stages. Horticulturae 2023, 9, 1198. [Google Scholar] [CrossRef]
- Kaur, H.; Kaur, H.; Kaur, H.; Srivastava, S. The Beneficial Roles of Trace and Ultratrace Elements in Plants. Plant Growth Regul. 2023, 100, 219–236. [Google Scholar] [CrossRef]
- Önder, D. Variation in Antioxidant Capacity, Antioxidant Activity and Mineral Composition during Flower Development of Oil-Bearing Rose (Rosa damascena Mill.). Sci. Rep. 2023, 13, 17255. [Google Scholar] [CrossRef]
- Jia, S.; Wang, Y.; Hu, J.; Ding, Z.; Liang, Q.; Zhang, Y.; Wang, H. Mineral and Metabolic Profiles in Tea Leaves and Flowers during Flower Development. Plant Physiol. Biochem. 2016, 106, 316–326. [Google Scholar] [CrossRef]
- Tian, J.; Gong, Y.; Li, J. Nutritional Attributes and Phenolic Composition of Flower and Bud of Sophora japonica L. and Robinia pseudoacacia L. Molecules 2022, 27, 8932. [Google Scholar] [CrossRef]
- Morrissey, J.; Guerinot, M.L. Trace Elements: Too Little or Too Much and How Plants Cope. F1000 Biol. Rep. 2009, 1, 14. [Google Scholar] [CrossRef]
- Rai, S.; Singh, P.K.; Mankotia, S.; Swain, J.; Satbhai, S.B. Iron Homeostasis in Plants and Its Crosstalk with Copper, Zinc, and Manganese. Plant Stress 2021, 1, 100008. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, C.; Hu, J.; Wu, K.; Gong, B.; Xu, Y. A Flowering Morphological Investigation, Fruit Fatty Acids, and Mineral Elements Dynamic Changes of Idesia polycarpa Maxim. Plants 2024, 13, 2663. [Google Scholar] [CrossRef]
- Niu, L.; Hou, Y.; Jiang, M.; Bai, G. The Rich Pharmacological Activities of Magnolia Officinalis and Secondary Effects Based on Significant Intestinal Contributions. J. Ethnopharmacol. 2021, 281, 114524. [Google Scholar] [CrossRef]
- Poivre, M.; Duez, P. Biological Activity and toxicity of the Chinese Herb Magnolia officinalis Rehder & E. Wilson (Houpo) and Its Constituents. J. Zhejiang Univ. Sci. B 2017, 18, 194–214. [Google Scholar] [CrossRef] [PubMed]
- Pires, E.D.O.; Di Gioia, F.; Rouphael, Y.; García-Caparrós, P.; Tzortzakis, N.; Ferreira, I.C.F.R.; Barros, L.; Petropoulos, S.A.; Caleja, C. Edible Flowers as an Emerging Horticultural Product: A Review on Sensorial Properties, Mineral and Aroma Profile. Trends Food Sci. Technol. 2023, 137, 31–54. [Google Scholar] [CrossRef]
- Yang, Y.; Qiu, Y.; Cheng, Y.; Yu, T.; Zhu, M.; Qian, W.; Gao, S.; Zhuang, G. Non-structural Carbohydrate Content and C:N:P Stoichiometry in Houpoea Officinalis Flowers in Response To Development Stages. Horticulturae 2024, 10, 784. [Google Scholar] [CrossRef]
- Zhao, S.; Xiong, F.; Wang, L.; Wang, B.; Chen, K.; Chen, C.; Zhou, G. Study on the Quality Characteristics and Geographical Origin Authentication of Wild Rheum Tanguticum in Three Authentic Regions. J. Food Compos. Anal. 2023, 123, 105463. [Google Scholar] [CrossRef]
- Zhang, X. Extraction from Different Parts of Citrus maxima Flowers Using Ultrasound as an Aid And Study of Their Composition and Function. Ultrason. Sonochem. 2023, 100, 106632. [Google Scholar] [CrossRef]
- Liu, Q. Characterization of the Phytochemical Content, Antioxidant Activity and Inhibition Capacity Against α-Glucosidase of Different Flower Parts of Seven Lotuses (Nelumbo). Sci. Hort. 2023, 316, 112007. [Google Scholar] [CrossRef]
- Zoroddu, M.A. The Essential Metals for Humans: A Brief Overview. J. Inorg. Biochem. 2019, 195, 120–129. [Google Scholar] [CrossRef]
- Jomova, K.; Makova, M.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Rhodes, C.J.; Valko, M. Essential Metals in Health and Disease. Chem. Biol. Interact. 2022, 367, 110173. [Google Scholar] [CrossRef]
- Al Alawi, A.M.; Majoni, S.W.; Falhammar, H. Magnesium and Human Health: Perspectives and Research Directions. Int. J. Endocrinol. 2018, 2018, 9041694. [Google Scholar] [CrossRef]
- Pohl, H.R.; Wheeler, J.S.; Murray, H.E. Sodium and Potassium in Health and Disease. In Interrelations Between Essential Metal Ions and Human Diseases; Metal Ions in Life Sciences; Springer: Berlin/Heidelberg, Germany, 2013; pp. 29–47. [Google Scholar] [CrossRef]
- Norikoshi, R.; Imanishi, H.; Ichimura, K. Changes in Cell Number, Osmotic Potential and Concentrations of Carbohydrates and Inorganic Ions in Tweedia caerulea During Flower Opening. J. Jpn. Soc. Hortic. Sci. 2013, 82, 51–56. [Google Scholar] [CrossRef]
- Weaver, C.M. Potassium and Health. Adv. Nutr. 2013, 4, 368S–377S. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Chen, X.; Su, H.; Eneji, A.E.; Guo, Y.; Dong, X. Changes of Secondary Metabolites and Trace Elements in Gentiana Macrophylla Flowers: A Potential Medicinal Plant Part. Chin. Herb. Med. 2014, 6, 145–151. [Google Scholar] [CrossRef]
- Sayeed, R.; Thakur, M.; Gani, A. Celosia cristata Linn. Flowers as a New Source of Nutraceuticals- a Study On Nutritional Composition, Chemical Characterization and In-vitro Antioxidant Capacity. Heliyon 2020, 6, e05792. [Google Scholar] [CrossRef]
- Katarzyna, J.; Karolina, J.; Patrycja, K.; Mateusz, B.; Izabela, G. Mineral Composition and Antioxidant Potential in the Common Poppy (Papaver rhoeas L.) Petal Infusions. Biol. Trace Elem. Res. 2021, 199, 371–381. [Google Scholar] [CrossRef]
- Mlcek, J.; Plaskova, A.; Jurikova, T.; Sochor, J.; Baron, M.; Ercisli, S. Chemical, Nutritional and Sensory Characteristics of Six Ornamental Edible Flowers Species. Foods 2021, 10, 2053. [Google Scholar] [CrossRef]
- Singh, M.; Nara, U.; Rani, N.; Pathak, D.; Sangha, M.K.; Kaur, K. Mineral Content Variation in Leaves, Stalks, and Seeds of Celery (Apium graveolens L.) Genotypes. Biol. Trace Elem. Res. 2023, 201, 2665–2673. [Google Scholar] [CrossRef]
- Abbaspour, N.; Hurrell, R.; Kelishadi, R. Review on Iron and Its Importance for Human Health. J. Res. Med.Sci. 2014, 19, 164–174. [Google Scholar]
- Xun, E. Translocation of Heavy Metals from Soils into Floral Organs and Rewards of Cucurbita pepo: Implications for Plant Reproductive Fitness. Ecotoxicol. Environ. Saf. 2017, 145, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Genchi, G.; Lauria, G.; Catalano, A.; Carocci, A.; Sinicropi, M.S. The Double Face of Metals: The Intriguing Case of Chromium. Appl. Sci. 2021, 11, 638. [Google Scholar] [CrossRef]
- Chen, P. Manganese Metabolism in Humans. Front. Biosci. 2018, 23, 1655–1679. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ma, Z. Research Progress on the Effects of Nickel on Hormone Secretion in The Endocrine Axis and on Target Organs. Ecotoxicol. Environ. Saf. 2021, 213, 112034. [Google Scholar] [CrossRef]
- Chen, L. Copper Homeostasis and Cuproptosis in Health and Disease. Signal Transduct. Target. Ther. 2022, 7, 378. [Google Scholar] [CrossRef]
- Pilmane, M. Strontium and Strontium Ranelate: Historical Review of Some of Their Functions. Mater. Sci. Eng. C 2017, 78, 1222–1230. [Google Scholar] [CrossRef]
- Yusof, E.N.M.; Ravoof, T.B.S.A.; Page, A.J. Cytotoxicity of Tin(Iv)-based Compounds: A Review. Polyhedron 2021, 198, 115069. [Google Scholar] [CrossRef]
- Ścibior, A.; Pietrzyk, Ł.; Plewa, Z.; Skiba, A. Vanadium: Risks and Possible Benefits in The Light of a Comprehensive Overview of Its Pharmacotoxicological Mechanisms and Multi-Applications with a Summary of Further Research Trends. J. Trace Elem. Med. Biol. 2020, 61, 126508. [Google Scholar] [CrossRef]
- Leyssens, L.; Vinck, B.; Van Der Straeten, C.; Wuyts, F.; Maes, L. Cobalt Toxicity in Humans-a Review of the Potential Sources and Systemic Health Effects. Toxicology 2017, 387, 43–56. [Google Scholar] [CrossRef]
- Li, S. Development and Applications of Aluminum Alloys for Aerospace Industry. J. Mater. Res. Technol. 2023, 27, 944–983. [Google Scholar] [CrossRef]
- Najafizadeh, M.; Yazdi, S.; Bozorg, M.; Ghasempour-Mouziraji, M.; Hosseinzadeh, M.; Zarrabian, M.; Cavaliere, P. Classification and Applications of Titanium and Its Alloys: A Review. J. Alloys Compd. Commun. 2024, 3, 100019. [Google Scholar] [CrossRef]
- Zhang, J.; Kuang, X.; Tu, R.; Zhang, S. A Review on Synthesis and Applications of Gallium Oxide Materials. Adv. Colloid Interface Sci. 2024, 328, 103175. [Google Scholar] [CrossRef]
- Gonçalves, D.A.; De Souza, I.D.; Rosa, A.C.G.; Melo, E.S.P.; Goncalves, A.-M.B.; De Oliveira, L.C.S.; Do Nascimento, V.A. Multi-wavelength Calibration: Determination of Trace Toxic Elements in Medicine Plants by ICP OES. Microchem. J. 2019, 146, 381–386. [Google Scholar] [CrossRef]
- Chen, Y.-G. Impacts of Heavy Metals and Medicinal Crops on Ecological Systems, Environmental Pollution, Cultivation, and Production Processes in China. Ecotoxicol. Environ. Saf. 2021, 219, 112336. [Google Scholar] [CrossRef] [PubMed]
- Li, X. Effect of Germination on Mineral Content Changes in Brown Rice (Oryza sativa L.). Biol. Trace Elem. Res. 2025, 203, 535–543. [Google Scholar] [CrossRef]
- Xu, L.; Sun, L.; Chen, Y.; Nie, T.; Zhu, H.; Yin, Z. Magnolia biondii Pamp.: A Comprehensive Review of The Pharmacognosy, Phytochemistry, Pharmacology, and Applications. Ind. Crops Prod. 2024, 222, 119648. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element Name | Abbreviation | Element Name | Abbreviation |
|---|---|---|---|
| Sodium | Na | Zinc | Zn |
| Magnesium | Mg | Aluminum | Al |
| Potassium | K | Chromium | Cr |
| Calcium | Ca | Gallium | Ga |
| Titanium | Ti | Strontium | Sr |
| Vanadium | V | Cadmium | Cd |
| Manganese | Mn | Tin | Sn |
| Iron | Fe | Barium | Ba |
| Cobalt | Co | Thallium | Tl |
| Nickel | Ni | Lead | Pb |
| Copper | Cu | Bismuth | Bi |
| Element | Background Correction Mode | λ (nm) | Lamp Current (mA) | Spectral Bandwidth | Flame Type | Gas Flow Rate (L/min) | Combustion Head Height |
|---|---|---|---|---|---|---|---|
| Na | NON-BGC | 589.0 | 3 | 0.2 nm | Air-C2H2 | 1.9 | 6 mm |
| Mg | BGC-D2 | 285.2 | 8 | 0.7 nm | Air-C2H2 | 2.0 | 7 mm |
| K | NON-BGC | 589.0 | 3 | 0.2 nm | Air-C2H2 | 1.9 | 6 mm |
| Ca | BGC-D2 | 422.7 | 4 | 0.7 nm | Air-C2H2 | 2.4 | 8 mm |
| Elements | Linear Equation | Correlation Coefficient (R2) | Linear Range (mg/L) | RSD (%) | LOD (ppm) | LOQ (ppm) |
|---|---|---|---|---|---|---|
| Na | y = 0.0248x + 0.0064 | R2 = 0.9932 | 0–0.5 | 0.034 | 0.001 | 0.004 |
| Mg | y = 0.7778x − 0.001 | R2 = 0.9997 | 0–0.2 | 0.024 | 0.0007 | 0.002 |
| K | y = 0.1995x + 0.0109 | R2 = 0.995 | 0–4 | 0.034 | 0.001 | 0.004 |
| Ca | y = 0.0647x + 0.0069 | R2 = 0.994 | 0–5 | 0.667 | 0.02 | 0.05 |
| Elements | Linear Equation | Correlation Coefficient (R2) | Linear Range (mg/L) | RSD (%) | LOD (ppm) | LOQ (ppm) |
|---|---|---|---|---|---|---|
| Ti | y = 2695.5x − 5938.5 | R2 = 0.9982 | 0–150 | 4.5% | 0.0328 | 0.1094 |
| V | y = 5329.2x − 1252.5 | R2 = 0.9997 | 0–150 | 2.8% | 0.0006 | 0.0019 |
| Mn | y = 4387.2x + 826.95 | R2 = 0.9997 | 0–150 | 2.2% | 0.0032 | 0.0107 |
| Fe | y = 137.14x − 388.5 | R2 = 0.9943 | 0–150 | 5.2% | 0.4357 | 1.4522 |
| Co | y = 9094x + 7055.6 | R2 = 0.9995 | 0–150 | 2.7% | 0.0007 | 0.0023 |
| Ni | y = 2437.9x + 1643.1 | R2 = 0.9993 | 0–150 | 0.9% | 0.0397 | 0.1322 |
| Cu | y = 6390x + 7327.4 | R2 = 0.9993 | 0–150 | 0.3% | 0.5929 | 1.9763 |
| Zn | y = 1327.4x + 2975.3 | R2 = 0.9992 | 0–150 | 2.2% | 0.1018 | 0.3394 |
| Al | y = 525.98x + 521.96 | R2 = 0.9996 | 0–150 | 0.6% | 0.0135 | 0.0450 |
| Cr | y = 6133.7x − 3264.9 | R2 = 0.9994 | 0–150 | 5.0% | 0.0077 | 0.0257 |
| Ga | y = 3813.4x − 8523.2 | R2 = 0.9987 | 0–150 | 6.8% | 0.0049 | 0.0165 |
| Sr | y = 6576.3x + 5639.7 | R2 = 0.9997 | 0–150 | 2.6% | 0.0029 | 0.0097 |
| Cd | y = 1830.2x + 1477.3 | R2 = 0.9996 | 0–150 | 2.4% | 0.0025 | 0.0084 |
| Sn | y = 3212.5x – 14,307 | R2 = 0.9948 | 0–150 | 3.3% | 0.0051 | 0.0170 |
| Ba | y = 12,831x + 31,426 | R2 = 0.9978 | 0–150 | 3.5% | 0.0038 | 0.0125 |
| Tl | y = 18,451x + 81,280 | R2 = 0.9938 | 0–150 | 3.5% | 0.0007 | 0.0024 |
| Pb | y = 14,793x + 61,577 | R2 = 0.9954 | 0–150 | 1.3% | 0.0013 | 0.0042 |
| Bi | y = 20,581x – 74,109 | R2 = 0.9905 | 0–150 | 1.5% | 0.0020 | 0.0065 |
| Stage 1 | Stage 2 | Stage 3 | Stage 4 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pistil | Stamens | Petals | Pistil | Stamens | Petals | Pistil | Stamens | Petals | Pistil | Stamens | Petals | |
| Na(mg/g) | 1.45 ± 0.23 e | 2.58 ± 0.35 abc | 1.82 ± 0.07 de | 3.08 ± 0.28 a | 2.31 ± 0.19 bcd | 1.67 ± 0.05 de | 1.63 ± 0.14 de | 1.81 ± 0.16 de | 2.24 ± 0.10 cd | 1.93 ± 0.13 cde | 2.90 ± 0.25 ab | 2.20 ± 0.23 cd |
| Mg(mg/g) | 6.34 ± 0.05 a | 5.07 ± 0.12 c | 5.70 ± 0.04 b | 6.62 ± 0.04 a | 4.96 ± 0.14 cd | 5.16 ± 0.06 c | 4.84 ± 0.12 cde | 3.86 ± 0.06 f | 4.64 ± 0.14 de | 5.64 ± 0.12 b | 4.55 ± 0.18 e | 4.87 ± 0.16 cde |
| K(mg/g) | 38.57 ± 0.34 c | 29.25 ± 0.44 ef | 44.93 ± 0.34 b | 34.39 ± 0.21 d | 31.26 ± 0.28 e | 48.06 ± 0.40 a | 25.80 ± 0.57 g | 25.78 ± 0.21 g | 44.73 ± 0.37 b | 34.52 ± 0.89 d | 28.43 ± 1.94 f | 48.06 ± 0.53 a |
| Ca(mg/g) | 20.00 ± 0.60 cde | 23.96 ± 2.5 bcd | 18.23 ± 0.10 de | 25.17 ± 2.90 bc | 24.64 ± 1.60 bc | 18.61 ± 1.48 de | 28.77 ± 3.01 ab | 17.77 ± 1.09 e | 17.82 ± 1.53 e | 31.00 ± 0.29 a | 17.27 ± 1.34 e | 22.88 ± 2.77 bcde |
| V(μg/g) | 0.81 ± 0.01 a | 0.64 ± 0.05 b | 0.51 ± 0.03 c | 0.46 ± 0.03 cd | 0.42 ± 0.01 de | 0.39 ± 0.01 def | 0.42 ± 0.04 de | 0.34 ± 0.01 ef | 0.37 ± 0.03 ef | 0.33 ± 0.01 f | 0.35 ± 0.01 ef | 0.33 ± 0.02 f |
| Cr(μg/g) | 8.15 ± 0.39 bc | 7.46 ± 0.77 bc | 8.77 ± 0.59 b | 8.48 ± 0.8 bc | 6.05 ± 0.49 c | 7.67 ± 0.94 bc | 7.36 ± 0.22 bc | 7.62 ± 1.15 bc | 5.13 ± 2.6 bc | 8.35 ± 0.95 bc | 12.49 ± 0.59 a | 9.74 ± 0.37 b |
| Mn(μg/g) | 92.02 ± 1.58 b | 51.56 ± 0.37 e | 85.99 ± 1.29 c | 110.64 ± 1.22 a | 42.59 ± 1.97 f | 61.28 ± 0.61 d | 29.92 ± 0.35 g | 26.36 ± 1.52 gh | 25.94 ± 1.00 gh | 24.54 ± 1.88 h | 28.27 ± 1.07 gh | 25.66 ± 1.95 gh |
| Fe(μg/g) | 419.58 ± 31.65 bcd | 400.94 ± 8.70 cde | 387.57 ± 40.961 cde | 485.12 ± 18.02 b | 360.01 ± 26.52 de | 333.11 ± 14.28 e | 370.51 ± 15.92 cde | 443.68 ± 24.67 bc | 324.27 ± 10.21 e | 372.19 ± 16.86 cde | 622.31 ± 25.98 a | 441.12 ± 32.09 bc |
| Co(μg/g) | 0.31 ± 0.01 a | 0.22 ± 0.02 bc | 0.19 ± 0.01 c | 0.21 ± 0.03 bc | 0.16 ± 0.01 c | 0.16 ± 0.01 c | 0.21 ± 0.01 bc | 0.18 ± 0.02 c | 0.18 ± 0.02 c | 0.17 ± 0.01 c | 0.26 ± 0.03 ab | 0.18 ± 0.01 c |
| Ni(μg/g) | 23.70 ± 0.63 b | 23.26 ± 0.92 b | 19.87 ± 0.77 cd | 34.37 ± 1.30 a | 23.95 ± 1.41 b | 21.62 ± 0.51 bc | 16.03 ± 0.74 ef | 14.43 ± 0.28 f | 17.99 ± 1.18 de | 14.48 ± 0.23 f | 17.90 ± 0.40 de | 22.77 ± 1.02 b |
| Cu(μg/g) | 18.82 ± 0.93 a | 16.93 ± 0.70 bc | 14.46 ± 0.41 d | 15.04 ± 0.87 d | 18.24 ± 0.79 ab | 14.51 ± 0.26 d | 14.08 ± 0.73 d | 13.78 ± 0.43 d | 14.14 ± 0.28 d | 14.48 ± 0.42 d | 13.88 ± 0.65 d | 15.80 ± 0.32 cd |
| Zn(μg/g) | 445.17 ± 33.72 g | 568.88 ± 50.12 efg | 691.47 ± 61.87 def | 1553.16 ± 92.64 a | 615.97 ± 70.47 defg | 469.02 ± 8.27 g | 1368.03 ± 87.30 b | 986.18 ± 73.29 c | 750.23 ± 61.56 de | 502.42 ± 48.95 fg | 788.36 ± 44.13 d | 612.40 ± 28.67 defg |
| Sr(μg/g) | 39.59 ± 0.44 ab | 36.35 ± 2.30 abcd | 33.33 ± 0.03 cde | 40.47 ± 3.01 a | 34.51 ± 1.11 bcde | 31.75 ± 0.70 de | 36.47 ± 1.73 abcd | 30.19 ± 1.75 e | 32.69 ± 1.72 de | 39.34 ± 1.44 ab | 35.34 ± 1.36 abcde | 38.51 ± 1.64 abc |
| Sn(μg/g) | 22.07 ± 1.25 a | 19.94 ± 0.68 b | 17.54 ± 0.30 c | 15.71 ± 0.44 d | 14.78 ± 0.19 de | 14.35 ± 0.36 de | 13.80 ± 0.17 ef | 12.65 ± 0.04 fg | 11.71 ± 0.12 gh | 10.94 ± 0.14 hi | 9.78 ± 0.32 ij | 8.96 ± 0.13 j |
| Al(μg/g) | 228.09 ± 7.66 bc | 179.21 ± 4.20 fg | 188.50 ± 5.75 efg | 226.63 ± 15.91 bc | 224.06 ± 11.94 bcd | 165.91 ± 6.06 g | 207.74 ± 8.51 cde | 200.93 ± 4.22 def | 200.30 ± 5.00 def | 235.29 ± 4.41 b | 271.63 ± 7.83 a | 214.53 ± 1.36 bcd |
| Ti(μg/g) | 996.73 ± 18.13 ef | 966.52 ± 6.48 f | 969.82 ± 14.05 f | 1030.84 ± 10.47 de | 1030.86 ± 28.60 de | 1051.60 ± 15.05 cd | 1089.03 ± 11.64 bc | 1133.76 ± 1.25 ab | 1137.49 ± 8.83 a | 1151.35 ± 19.44 a | 1178.16 ± 20.04 a | 1176.35 ± 13.65 a |
| Ga(μg/g) | 0.94 ± 0.06 a | 0.74 ± 0.06 b | 0.55 ± 0.02 cd | 0.61 ± 0.03 c | 0.47 ± 0.005 de | 0.41 ± 0.02 d | 0.56 ± 0.03 cd | 0.41 ± 0.002 d | 0.37 ± 0.02 d | 0.54 ± 0.04 cd | 0.42 ± 0.02 d | 0.46 ± 0.02 de |
| Cd(μg/g) | 0.28 ± 0.02 bc | 0.31 ± 0.05 bc | 0.36 ± 0.04 b | 0.55 ± 0.04 a | 0.27 ± 0.05 bc | 0.23 ± 0.02 bcd | 0.18 ± 0.03 cd | 0.10 ± 0.04 d | 0.25 ± 0.05 bcd | 0.37 ± 0.09 b | 0.29 ± 0.04 bc | 0.19 ± 0.02 cd |
| Ba(μg/g) | 31.72 ± 1.90 a | 32.44 ± 1.00 a | 25.44 ± 0.72 bc | 31.05 ± 1.26 a | 25.88 ± 0.45 bc | 23.93 ± 0.91 bc | 33.31 ± 1.81 a | 23.83 ± 0.98 bc | 22.65 ± 1.20 c | 32.43 ± 1.87 a | 25.05 ± 0.73 bc | 27.19 ± 1.37 b |
| Tl(μg/g) | 2.36 ± 0.07 a | 1.12 ± 0.13 b | 0.52 ± 0.04 c | 0.27 ± 0.02 d | 0.18 ± 0.01 de | 0.12 ± 0.01 e | 0.10 ± 0.01 e | 0.07 ± 0.00 e | 0.06 ± 0.00 e | 0.05 ± 0.00 e | 0.04 ± 0.00 e | 0.04 ± 0.00 e |
| Pb(μg/g) | 3.14 ± 0.27 b | 1.61 ± 0.04 c | 0.97 ± 0.09 ef | 3.75 ± 0.08 a | 1.13 ± 0.07 def | 0.86 ± 0.07 f | 1.41 ± 0.13 cd | 0.87 ± 0.04 f | 1.01 ± 0.05 ef | 1.23 ± 0.07 de | 1.17 ± 0.02 def | 0.87 ± 0.02 f |
| Bi(μg/g) | 41.26 ± 0.55 a | 29.66 ± 2.37 b | 23.56 ± 0.48 c | 20.82 ± 0.43 d | 19.12 ± 0.20 de | 18.07 ± 0.61 e | 17.23 ± 0.32 ef | 15.43 ± 0.49 fg | 13.92 ± 0.35 gh | 11.94 ± 0.18 hi | 10.29 ± 0.55 ij | 8.93 ± 0.41 j |
| Component | Eigenvalue | Percentage of Variance (%) | Cumulative (%) | Component | Eigenvalue | Percentage of Variance (%) | Cumulative (%) |
|---|---|---|---|---|---|---|---|
| 1 | 8.379 | 38.088 | 38.088 | 12 | 0.163 | 0.739 | 97.933 |
| 2 | 4.035 | 18.339 | 56.427 | 13 | 0.126 | 0.574 | 98.508 |
| 3 | 2.361 | 10.732 | 67.160 | 14 | 0.115 | 0.521 | 99.029 |
| 4 | 2.134 | 9.700 | 76.860 | 15 | 0.056 | 0.254 | 99.283 |
| 5 | 1.608 | 7.310 | 84.170 | 16 | 0.051 | 0.230 | 99.513 |
| 6 | 0.973 | 4.423 | 88.593 | 17 | 0.040 | 0.184 | 99.697 |
| 7 | 0.606 | 2.754 | 91.347 | 18 | 0.030 | 0.138 | 99.835 |
| 8 | 0.471 | 2.141 | 93.488 | 19 | 0.016 | 0.071 | 99.906 |
| 9 | 0.347 | 1.576 | 95.064 | 20 | 0.012 | 0.054 | 99.961 |
| 10 | 0.283 | 1.287 | 96.350 | 21 | 0.005 | 0.022 | 99.982 |
| 11 | 0.186 | 0.843 | 97.194 | 22 | 0.004 | 0.018 | 100.000 |
| PC1 | PC2 | PC3 | PC4 | PC5 | |
|---|---|---|---|---|---|
| Na | 0.007 | 0.35 | −0.113 | 0.171 | −0.009 |
| Mg | 0.276 | 0.105 | −0.223 | 0.083 | 0.176 |
| K | −0.002 | −0.165 | −0.172 | 0.291 | 0.557 |
| Ca | 0.067 | 0.19 | −0.297 | −0.499 | 0.052 |
| V | 0.307 | −0.13 | 0.196 | −0.01 | −0.012 |
| Cr | −0.05 | 0.272 | 0.323 | 0.186 | 0.22 |
| Mn | 0.277 | 0.005 | −0.138 | 0.33 | −0.055 |
| Fe | 0.004 | 0.365 | 0.323 | 0.185 | −0.062 |
| Co | 0.193 | 0.129 | 0.433 | 0.061 | −0.016 |
| Ni | 0.212 | 0.138 | −0.245 | 0.314 | 0.013 |
| Cu | 0.229 | −0.047 | 0.039 | −0.158 | 0.277 |
| Zn | −0.006 | 0.247 | −0.162 | 0.106 | −0.561 |
| Sr | 0.187 | 0.293 | −0.105 | −0.243 | 0.286 |
| Sn | 0.301 | −0.194 | 0.041 | 0.016 | −0.134 |
| Al | 0.001 | 0.378 | 0.247 | −0.08 | 0.115 |
| Ti | −0.248 | 0.225 | 0.113 | −0.105 | 0.186 |
| Ga | 0.313 | −0.011 | 0.116 | −0.173 | −0.016 |
| Cd | 0.167 | 0.233 | −0.303 | 0.149 | 0.114 |
| Ba | 0.197 | 0.163 | −0.073 | −0.413 | −0.1 |
| Tl | 0.296 | −0.133 | 0.252 | −0.045 | 0.07 |
| Pb | 0.274 | 0.182 | −0.055 | 0.112 | −0.166 |
| Bi | 0.311 | −0.177 | 0.122 | −0.01 | −0.076 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zhu, M.-Y.; Zhao, S.-M.; Fan, Y.-T.; Huang, J.-W.; Yu, T.; Zhuang, G.-Q.; Gao, S. Variations in the Mineral Composition of Houpoea Officinalis Flowers at Different Stages of Development. Horticulturae 2025, 11, 387. https://doi.org/10.3390/horticulturae11040387
Yang Y, Zhu M-Y, Zhao S-M, Fan Y-T, Huang J-W, Yu T, Zhuang G-Q, Gao S. Variations in the Mineral Composition of Houpoea Officinalis Flowers at Different Stages of Development. Horticulturae. 2025; 11(4):387. https://doi.org/10.3390/horticulturae11040387
Chicago/Turabian StyleYang, Yao, Mao-Yuan Zhu, Shi-Mei Zhao, Yi-Tong Fan, Jing-Wen Huang, Ting Yu, Guo-Qing Zhuang, and Shun Gao. 2025. "Variations in the Mineral Composition of Houpoea Officinalis Flowers at Different Stages of Development" Horticulturae 11, no. 4: 387. https://doi.org/10.3390/horticulturae11040387
APA StyleYang, Y., Zhu, M.-Y., Zhao, S.-M., Fan, Y.-T., Huang, J.-W., Yu, T., Zhuang, G.-Q., & Gao, S. (2025). Variations in the Mineral Composition of Houpoea Officinalis Flowers at Different Stages of Development. Horticulturae, 11(4), 387. https://doi.org/10.3390/horticulturae11040387