
Enhancement of Growth and Quality of Winter Watermelon Using LED Supplementary Lighting


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Cultivation Conditions
2.2. Experimental Design and Supplementary LED Lighting Treatment
2.3. Methods of Sampling and Measurement
2.4. Statistical Analysis
3. Results
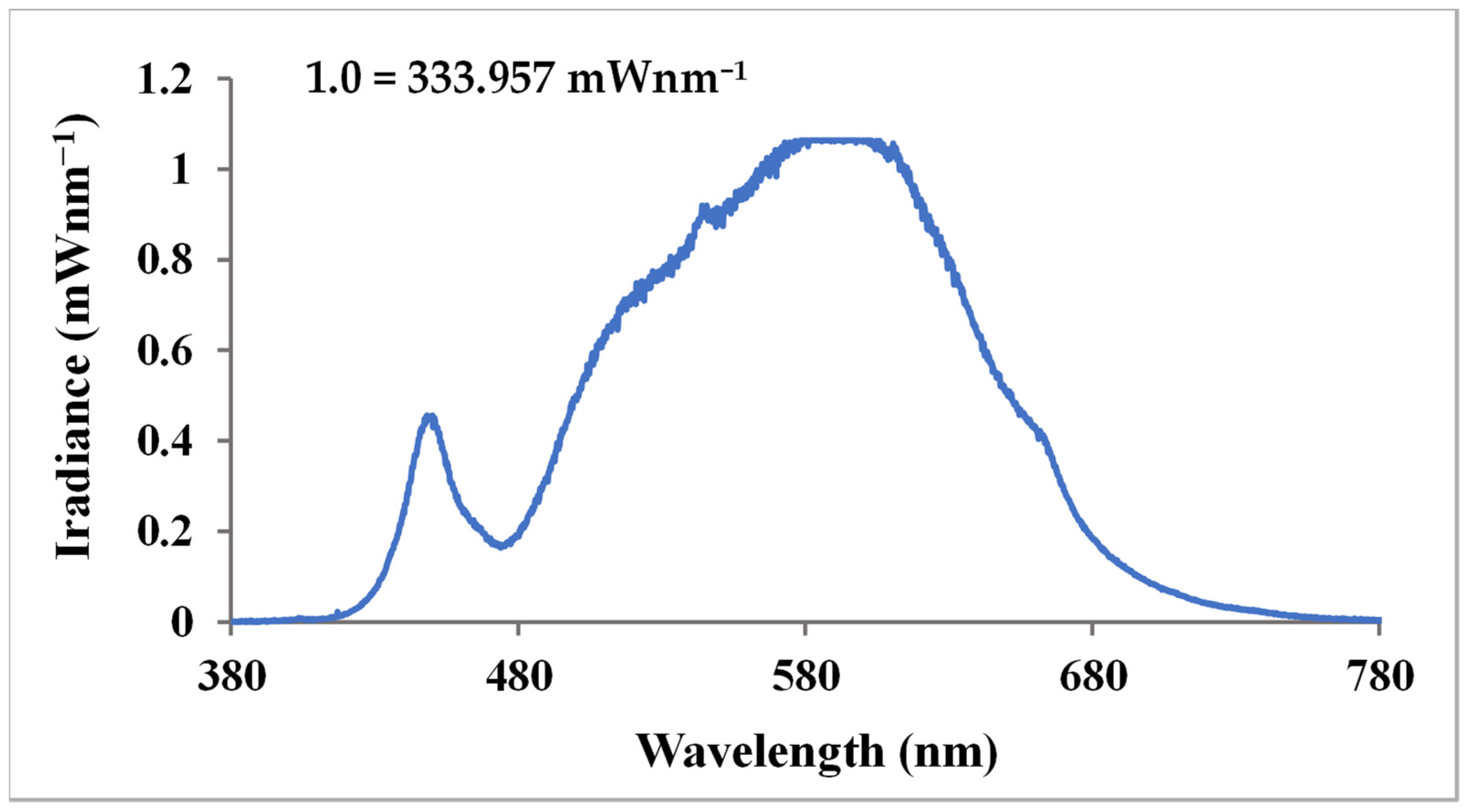
3.1. Determining Light Intensity and Environmental Conditions in Greenhouses
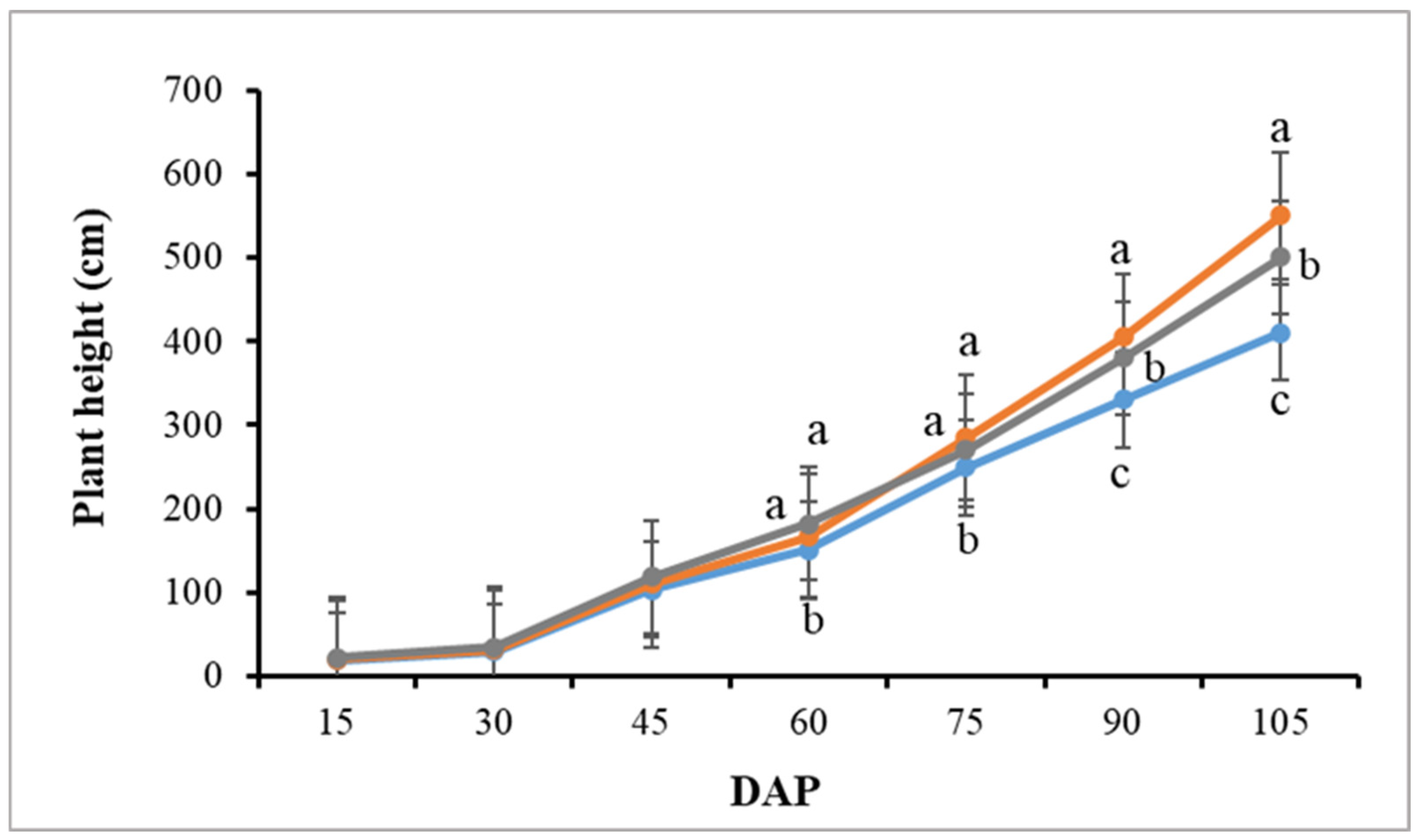
3.2. Plant Growth and Morphology
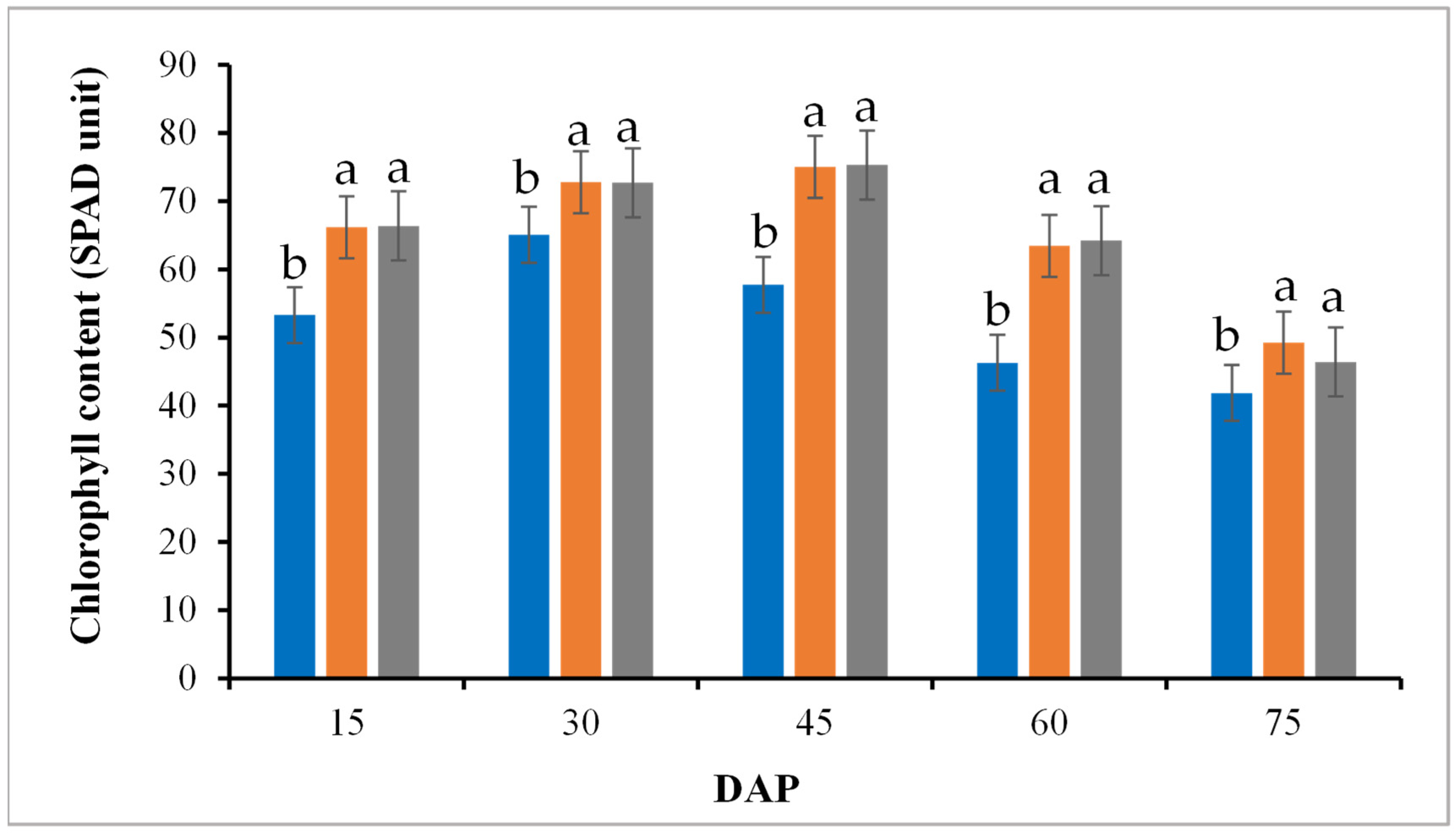
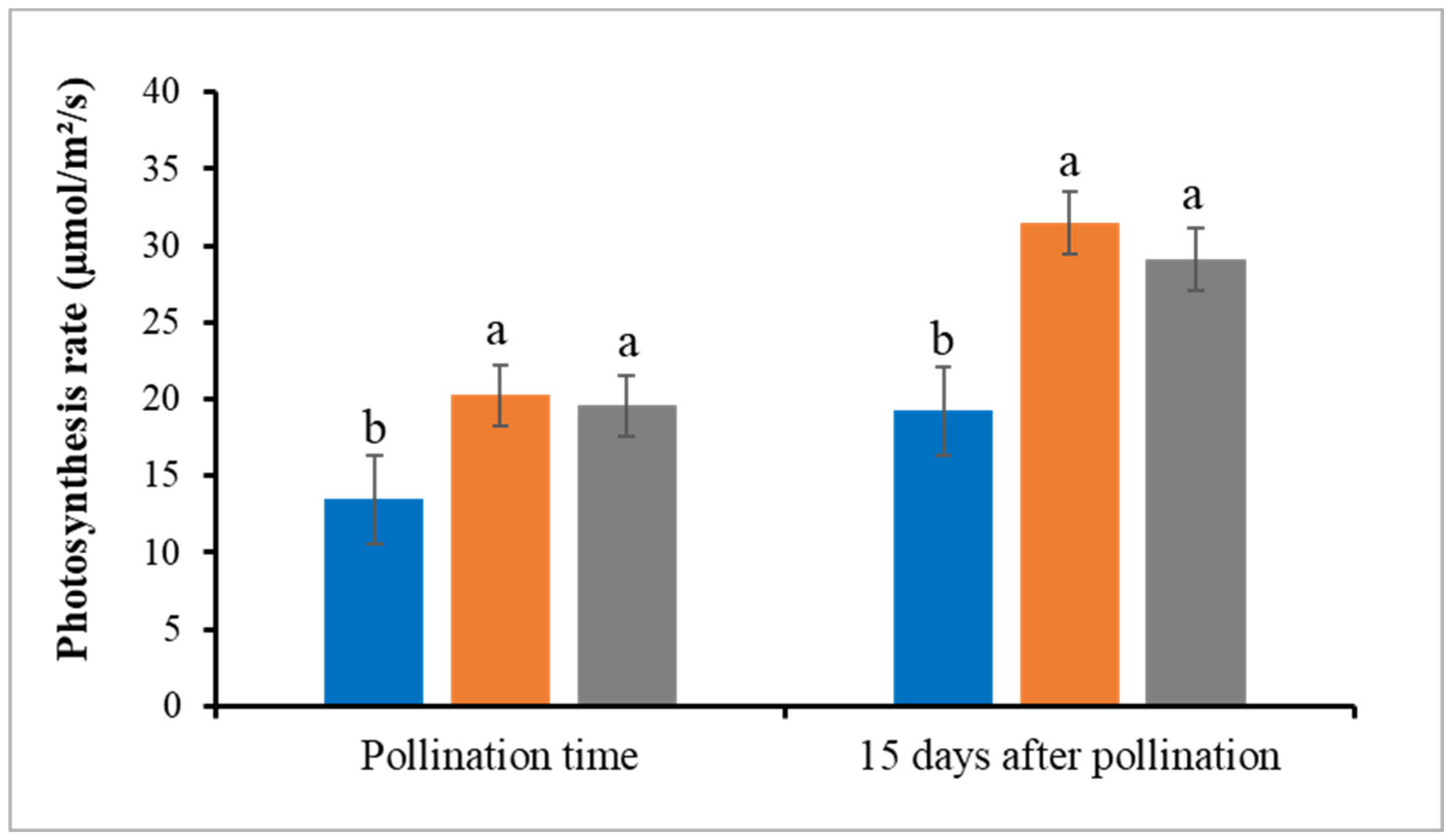
3.3. Leaf Chlorophyll Content and Photosynthesis
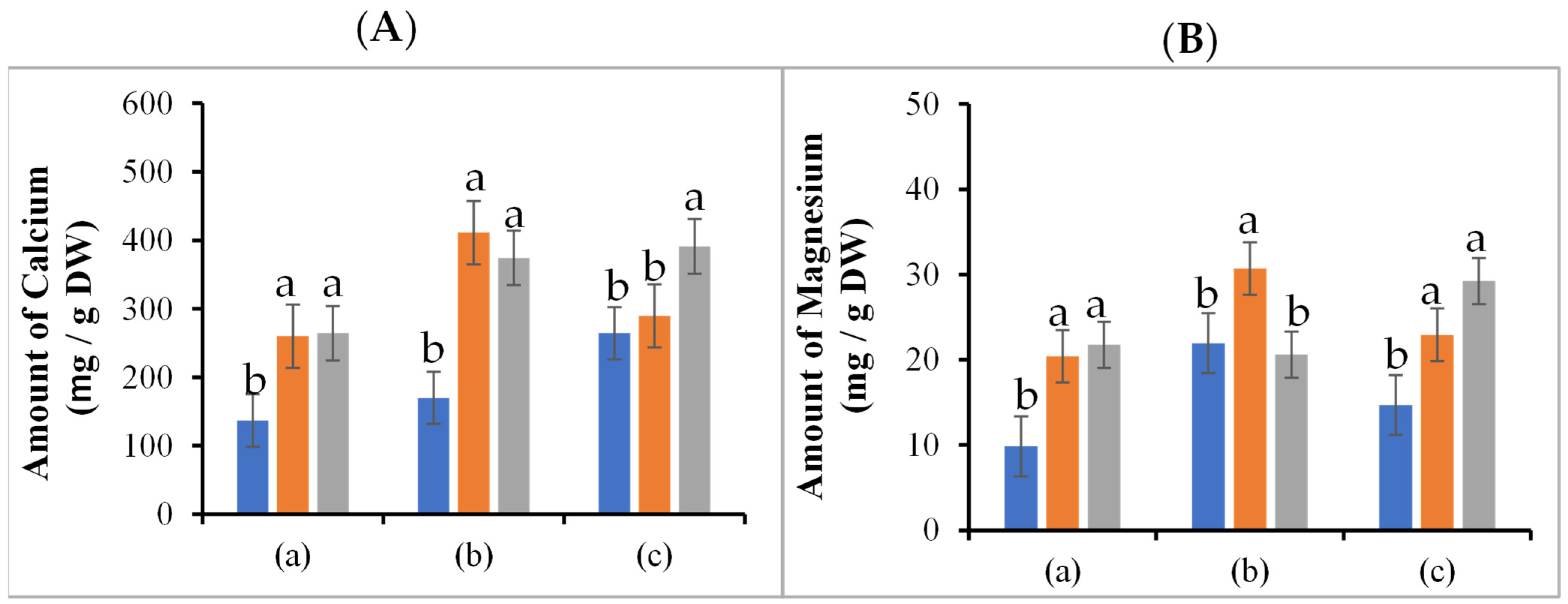
3.4. Ca2+ and Mg2+ Content in the Leaves of the Fruit Set Region
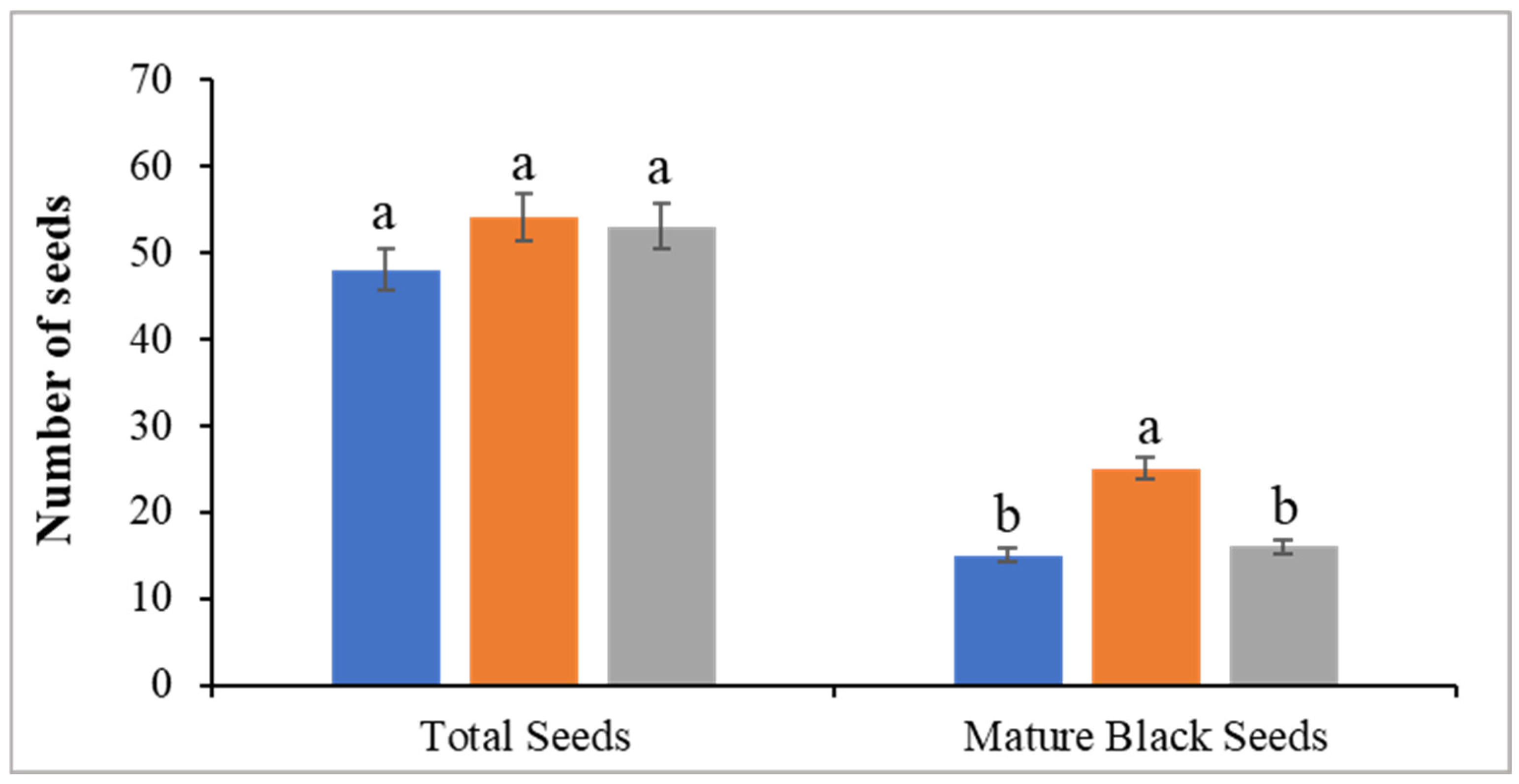
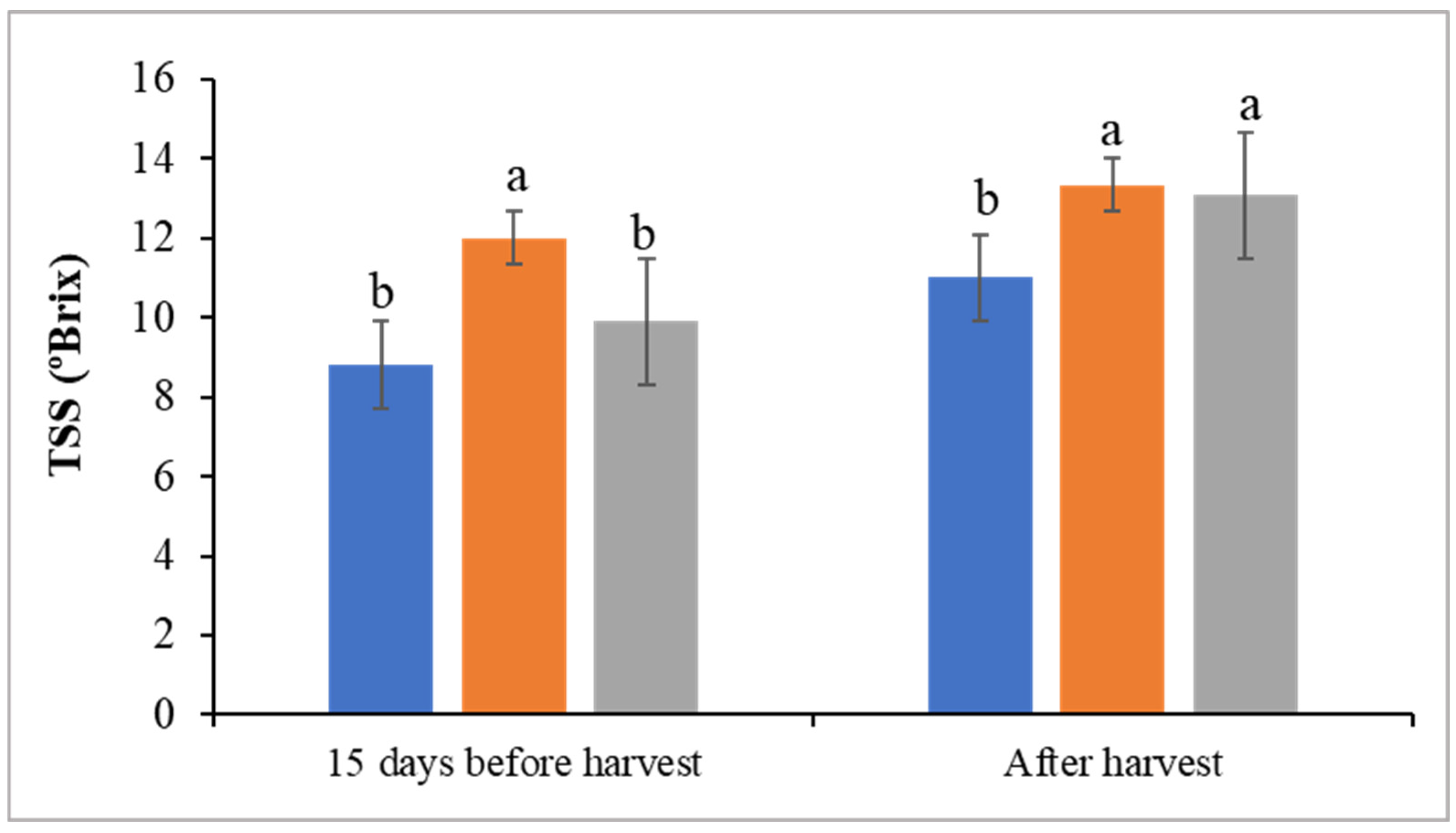
3.5. Fruit Yield and Quality
4. Discussion
4.1. LEDs Accelerate Early Growth and Have a Positive Effect Later on
4.2. In Winter, Strong Light Like That in Summer Is Not Necessarily Required
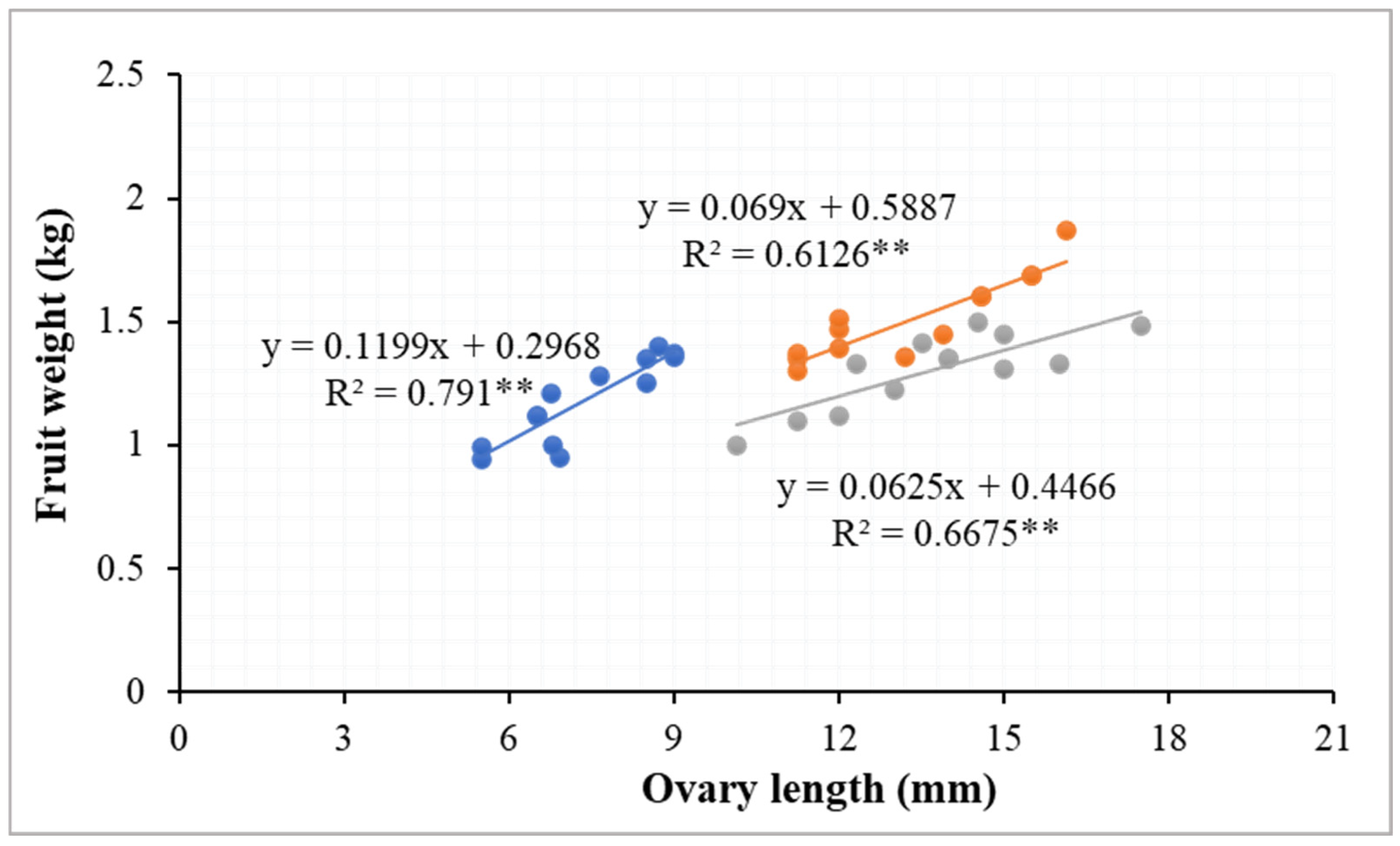
4.3. Fruit Characteristics of Winter-Grown Watermelons
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DAP | Days after pollination |
| Ca2+ | Calcium ion |
| Mg2+ | Magnesium ion |
References
- Reddy, U.K.; Abburi, L.; Abburi, V.L.; Saminathan, T.; Cantrell, R.; Vajja, V.G.; Reddy, R.; Tomason, Y.R.; Levi, A.; Wehner, T.C.; et al. A genome-wide scan of selective sweeps and association mapping of fruit traits using microsatellite markers in watermelon. J. Hered. 2015, 106, 166–176. [Google Scholar] [CrossRef]
- Hashizume, T. Breeding and utilization of watermelon. J. Jpn. Soc. Food Sci. 2019, 66, 314–318. [Google Scholar] [CrossRef]
- Levi, A.; Jarret, R.; Kousik, S.; Patrick Wechter, W.P.; Nimmakayala, P.; Reddy, U.K. Genetic resources of watermelon. In Genetics and Genomics of Cucurbitaceae, Plant Genetics and Genomics: Crops and Models; Grumet, R., Katzir, N., Garcia-Mas, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 87–110. [Google Scholar] [CrossRef]
- Chamchum, W.; Glahan, S.; Kramchote, S.; Maniwara, P.; Suwor, P. Growth and yield of watermelon (Citrullus lanatus) in plastic house in response to white led supplementary lighting. Agrivita J. Agr. Sci. 2023, 45, 271–277. [Google Scholar] [CrossRef]
- Stamford, J.D.; Stevens, J.; Mullineaux, P.M.; Lawson, T. LED lighting: A grower’s guide to light spectra. Hortscience 2023, 58, 180–196. [Google Scholar] [CrossRef]
- Folta, K.M.; Carvalho, S.D. Photoreceptors and control of horticultural plant traits. Hortic. Sci. 2015, 50, 1274–1280. [Google Scholar] [CrossRef]
- Fukuda, N.; Fujita, M.; Ohta, Y.; Sase, S.; Nishimura, S.; Ezura, H. Directional blue light irradiation triggers epidermal cell elongation of abaxial side resulting in inhibition of leaf epinasty in geranium under red light condition. Sci. Hortic. 2008, 115, 176–182. [Google Scholar] [CrossRef]
- Olarewaju, O.O.; Fajinmi, O.O.; Arthur, G.D.; Coopoosamy, R.M.; Naidoo, K. Effect of climate change on the production of Cucurbitaceae species in North African countries. J. Agric. Food Res. 2023, 14, 100742. [Google Scholar] [CrossRef]
- Varaldo, A.; Alchera, F.; Giuggioli, N.R.; Giacalone, G. LED UVB postharvest treatments modify nutraceutical quality and physical properties of ‘Cargo’ blueberries. J. Agric. Food Res. 2024, 15, 101040. [Google Scholar] [CrossRef]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.O. Spectral effects of artificial light on plant physiology and secondary metabolism: A review. Hortic. Sci. 2015, 50, 1128–1135. [Google Scholar] [CrossRef]
- Soufi, H.R.; Roosta, H.R.; Fatehi, F.; Ghorbanpour, M. Spectral composition of LED light differentially affects biomass, photosynthesis, nutrient profile, and foliar nitrate accumulation of lettuce grown under various replacement methods of nutrient solution. Food Sci. Nutr. 2023, 11, 8143–8162. [Google Scholar] [CrossRef]
- Bantis, F.; Koukounaras, A.; Siomos, A.S.; Radoglou, K.; Dangitsis, C. Optimal LED wavelength composition for the production of high-quality watermelon and interspecific squash seedlings used for grafting. Agronomy 2019, 9, 870. [Google Scholar] [CrossRef]
- Xiao, L.; Shibuya, T.; Kato, K.; Nishiyama, M.; Kanayama, Y. Effects of light quality on plant development and fruit metabolism and their regulation by plant growth regulators in tomato. Sci. Hortic. 2022, 300, 111076. [Google Scholar] [CrossRef]
- Tang, N.; Wang, J.; Zhang, B.; Chen, H.; Qiu, M. Chromatic effects of supplemental light on the fruit quality of strawberries. Horticulturae 2023, 9, 1333. [Google Scholar] [CrossRef]
- Hao, X.; Little, C.; Zheng, J.M.; Cao, R. Far-red LEDs improve fruit production in greenhouse tomato grown under high-pressure sodium lighting. Acta Hortic. 2016, 1134, 95–102. [Google Scholar] [CrossRef]
- Park, Y.H.; Seo, B.S.; Seo, J.B.; Shin, G.H.; Yun, B.G. Effects of supplemental light on quality, yield, and growth rate of tomatoes. Acta Hortic. 2016, 1129, 105–110. [Google Scholar] [CrossRef]
- Choi, H.G.; Moon, B.Y.; Kang, N.J. Effects of LED light on the production of strawberry during cultivation in a plastic greenhouse and in a growth chamber. Sci. Hortic. 2015, 189, 22–31. [Google Scholar] [CrossRef]
- Pérez-Romero, L.F.; Stirling, P.J.; Hancock, R.D. Light-emitting diodes improve yield, quality and inhibitory effects on digestive enzymes of strawberry. Sci. Hortic. 2024, 332, 113192. [Google Scholar] [CrossRef]
- Cui, X.H.; Guo, X.O.; Sun, T.Y.; Qi, H.Y. Effects of LED supplementary lighting on seedling growth and fruit quality of oriental melon. Plant Physiol. J. 2017, 53, 657–667. [Google Scholar] [CrossRef]
- Demers, D.A.; Gosselin, A.; Wien, H.C. Effects of supplemental light duration on greenhouse sweet pepper plants and fruit yields. J. Am. Soc. Hortic. Sci. 1998, 123, 202–207. [Google Scholar] [CrossRef]
- Zhou, C.; Li, Z.; Liu, W.; Bian, Z.; Lu, W.; Zhou, B.; Wang, S.; Li, Q.; Yang, Q. High-proportion blue light irradiation at the end-of-production stage promotes the biosynthesis and recycling of ascorbate in lettuce. Int. J. Mol. Sci. 2023, 24, 16524. [Google Scholar] [CrossRef]
- Nadalini, S.; Zucchi, P.; Andreotti, C. Effects of blue and red LED lights on soilless cultivated strawberry growth performances and fruit quality. Eur. J. Hortic. Sci. 2017, 82, 12–20. [Google Scholar] [CrossRef]
- Noh, J.; Kim, J.M.; Sheikh, S.; Lee, S.G.; Lim, J.H.; Seong, M.H.; Jung, G.T. Effect of heat treatment around the fruit set region on growth and yield of watermelon [Citrullus lanatus (Thunb.) Matsum. and Nakai]. Physiol. Mol. Biol. Plants 2013, 19, 509–514. [Google Scholar] [CrossRef]
- Lu, W.; Edelson, J.V.; Duthie, J.A.; Roberts, B.W. A comparison of yield between high- and low-intensity management for three watermelon cultivars. HortScience 2003, 38, 351–356. [Google Scholar] [CrossRef]
- Hidaka, K.; Okamoto, A.; Araki, T.; Miyoshi, Y.; Dan, K.; Imamura, H.; Kitano, M.; Sameshima, K.; Okimura, M. Effect of photoperiod of supplemental lighting with light-emitting diodes on growth and yield of strawberry. Environ. Control Biol. 2014, 52, 63–71. [Google Scholar] [CrossRef]
- Hidaka, K.; Dan, K.; Imamura, H.; Takayama, T.; Sameshima, K.; Okimura, M. Variety comparison of effect of supplemental lighting with LED on growth and yield in forcing culture of strawberry. Environ. Control Biol. 2015, 53, 135–143. [Google Scholar] [CrossRef]
- McNellis, T.W.; Deng, X.W. Light control of seedling morphogenetic pattern. Plant Cell 1995, 7, 1749–1761. [Google Scholar] [CrossRef]
- Gómez, C.; Mitchell, C.A. Physiological and productivity responses of high-wire tomato as affected by supplemental light source and distribution within the canopy. J. Am. Soc. Hortic. Sci. 2016, 141, 196–208. [Google Scholar] [CrossRef]
- Tewolde, F.T.; Shiina, K.; Maruo, T.; Takagaki, M.; Kozai, T.; Yamori, W. Supplemental LED inter-lighting compensates for a shortage of light for plant growth and yield under the lack of sunshine. PLoS ONE 2018, 13, e0206592. [Google Scholar] [CrossRef]
- Paponov, M.; Kechasov, D.; Lacek, J.; Verheul, M.J.; Paponov, I.A. Supplemental light-emitting diode inter-lighting increases tomato fruit growth through enhanced photosynthetic light use efficiency and modulated root activity. Front. Plant Sci. 2019, 10, 1656. [Google Scholar] [CrossRef]
- Anderson, C.M.; Mattoon, E.M.; Zhang, N.; Becker, E.; McHargue, W.; Yang, J.; Patel, D.; Dautermann, O.; McAdam, S.A.M.; Tarin, T.; et al. High light and temperature reduce photosynthetic efficiency through different mechanisms in the C4 model Setaria viridis. Commun. Biol. 2021, 4, 1092. [Google Scholar] [CrossRef]
- Wimalasekera, R. Effect of light intensity on photosynthesis. In Photosynthesis, Productivity and Environmental Stress; Ahmad, P., Ahanger, M.S., Alyemeni, M.N., Alam, P., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 65–73. [Google Scholar] [CrossRef]
- Hanyu, H.; Shoji, K. Acceleration of growth in spinach by short-ter exposure to red, blue light at the beginning and the end of the daily dark period. Acta Hortic. 2002, 580, 145–150. [Google Scholar] [CrossRef]
- Kalathil, N.; Thirunavookarasu, N.; Lakshmipathy, K.; Chidanand, D.V.; Radhakrishnan, M.; Baskaran, N. Application of light-based, non-thermal techniques to determine physico-chemical characteristics, pungency and aflatoxin levels of dried red chilli pods (Capsicum annuum). J. Agric. Food Res. 2023, 13, 100648. [Google Scholar] [CrossRef]
- Hori, H. Gravel culture of vegetable and ornamental crops. Agric. Hortic. 1966, 210. (In Japanese) [Google Scholar]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Lee, J.H.; Nam, S.Y. Vegetative propagation of six Pachyphytum species as influenced by different LED light qualities. Hortic. Sci. Technol. 2023, 41, 237–249. [Google Scholar] [CrossRef]
- Ahmed, N.; Zhang, B.; Bozdar, B.; Chachar, S.; Rai, M.; Li, J.; Li, Y.; Hayat, F.; Chachar, Z.; Tu, P. The power of magnesium: Unlocking the potential for increased yield, quality, and stress tolerance of horticultural crops. Front. Plant Sci. 2023, 14, 1285512. [Google Scholar] [CrossRef]
- Wei, H.; Wang, M.; Jeong, B.R. Effect of supplementary lighting duration on growth and activity of antioxidant enzymes in grafted watermelon seedlings. Agronomy 2020, 10, 337. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, B.; Teng, Z.; Zhang, M.; Yu, L.; Luo, Y.; Chen, P.; Sun, J. Improved metabolomic approach for evaluation of phytochemicals in mustard, kale, and broccoli microgreens under different controlled environment agriculture conditions. J. Agric. Food Res. 2023, 14, 100719. [Google Scholar] [CrossRef]
- Dong, C.; Fu, Y.; Liu, G.; Liu, H. Growth, photosynthetic characteristics, antioxidant capacity and biomass yield and quality of wheat (Triticum aestivum L.) exposed to LED light sources with different spectra combinations. J. Agron. Crop Sci. 2014, 200, 219–230. [Google Scholar] [CrossRef]
- Sobczak, A.; Kowalczyk, K.; Gajc-Wolska, J.; Kowalczyk, W.; Niedzińska, M. Growth, yield and quality of sweet pepper fruits fertilized with polyphosphates in hydroponic cultivation with LED lighting. Agronomy 2020, 10, 1560. [Google Scholar] [CrossRef]
- Piovene, C.; Orsini, F.; Bosi, S.; Sanoubar, R.; Bregola, V.; Dinelli, G.; Gianquinto, G. Optimal red:blue ratio in led lighting for nutraceutical indoor horticulture. Sci. Hortic. 2015, 193, 202–208. [Google Scholar] [CrossRef]
- Choi, H.G.; Moon, B.Y.; Kang, N.J. Correlation between strawberry (Fragaria ananassa Duch.) productivity and photosynthesis-related parameters under various growth conditions. Front. Plant Sci. 2016, 7, 1607. [Google Scholar] [CrossRef]
- Nawaz, M.; Sun, J.; Shabbir, S.; Khattak, W.A.; Ren, G.; Nie, X.; Bo, Y.; Javed, Q.; Du, D.; Sonne, C. A review of plants’ strategies to resist biotic and abiotic environmental stressors. Sci. Total Environ. 2023, 900, 165832. [Google Scholar] [CrossRef]
- O’Carrigan, A.; Hinde, E.; Lu, N.; Xu, X.-Q.; Duan, H.; Huang, G.; Mak, M.; Bellotti, W.; Chen, Z.-H. Effects of light irradiance on stomatal regulation and growth of tomato. Environ. Exp. Bot. 2014, 98, 65–73. [Google Scholar] [CrossRef]
- Kramchote, S.; Glahan, S. Effects of LED supplementary lighting and NPK fertilization on fruit quality of melon (Cucumis melo L.) grown in plastic house. J. Hortic. Res. 2020, 28, 111–122. [Google Scholar] [CrossRef]
- Klatt, B.K.; de La Vega, B.; Smith, H.G. Altered winter conditions impair plant development and yield in oilseed rape. J. Agric. Food Res. 2021, 5, 100160. [Google Scholar] [CrossRef]
- Ntagkas, N.; de Vos RC, H.; Woltering, E.J.; Nicole CC, S.; Labrie, C.; Marcelis, L.F.M. Modulation of the tomato fruit metabolome by led light. Metabolites 2020, 10, 266. [Google Scholar] [CrossRef]
- Appolloni, E.; Paucek, I.; Pennisi, G.; Stringari, G.; Gabarrell Durany, X.; Orsini, F.; Gianquinto, G. Supplemental LED lighting improves fruit growth and yield of tomato grown under the sub-optimal lighting condition of a building integrated rooftop greenhouse (i-RTG). Horticulturae 2022, 8, 771. [Google Scholar] [CrossRef]
- Sweetman, C.; Deluc, L.G.; Cramer, G.R.; Ford, C.M.; Soole, K.L. Regulation of malate metabolism in grape berry and other developing fruits. Phytochemistry 2009, 70, 1329–1344. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inside the Greenhouse | Outside the Greenhouse | |||||||
|---|---|---|---|---|---|---|---|---|
| Month | T (°C) | RH (%) | DLI (mol m−2d−1) | T (°C) | RH (%) | DLI (mol m−2d−1) | ||
| Control | LED900 | LED1500 | ||||||
| January | 19.5 | 52 | 6.3 | 19.3 | 27.9 | 7.1 | 54 | 10.2 |
| February | 20.5 | 65 | 6.5 | 19.5 | 28.1 | 8.0 | 63 | 11.1 |
| March | 22.0 | 60 | 9.5 | 22.5 | 31.1 | 9.6 | 59 | 15.3 |
| April | 22.5 | 69 | 10.5 | 23.5 | 32.1 | 17.1 | 72 | 15.3 |
| May | 25.5 | 69 | 12.0 | 25.0 | 33.6 | 20.0 | 72 | 18.2 |
| Supplementary LED Lighting | Plant Height (cm) | Shoot Dry Weight (g) | Root Dry Weight (g) | Root Length (cm) | No. of Nodes | No. of Leaves | Stem Diam Eter (mm) | Leaf Area (cm2) |
|---|---|---|---|---|---|---|---|---|
| Control | 20.0 b | 26.8 b | 3.9 c | 11.0 b | 5.0 b | 8.1 b | 4.1 b | 131.2 b |
| LED600 | 22.6 b | 31.2 ab | 5.2 b | 13.6 a | 5.7 ab | 9.8 ab | 4.2 b | 135.1 b |
| LED900 | 29.5 a | 35.6 a | 6.1 a | 13.6 a | 7.0 a | 11.6 a | 5.3 a | 157.3 a |
| LED1200 | 26.0 ab | 32.1 a | 5.6 b | 12.1 b | 6.3 a | 11.1 a | 5.0 a | 143 ab |
| SE | 2.1 | 1.2 | 0.5 | 1.0 | 0.5 | 0.7 | 0.1 | 9.5 |
| Supplementary LED Lighting | No. of Leaves | No. of Nodes | Stem Diameter (mm) | Leaf Length (cm) | Leaf Area (cm2) | Fresh Weight of Plant (g) | Dry Weight of Plant (g) |
|---|---|---|---|---|---|---|---|
| 56 DAP | |||||||
| Control | 87 c | 75 c | 5.2 b | 21.3 b | 187.7 c | ||
| LED900 | 140 a | 103 a | 6.8 a | 24.8 a | 236.3 a | ||
| LED1500 | 110 b | 93 b | 6.1 ab | 24.6 a | 226.2 b | ||
| SE | 8.0 | 4.2 | 0.0 | 1.0 | 12.3 | ||
| 86 DAP | |||||||
| Control | 180 c | 168 c | 8.3 b | 16.6 b | 128.6 c | 754.2 c | 80.3 b |
| LED900 | 260 a | 248 a | 10.4 a | 21.6 a | 190.4 a | 1039.8 a | 116.9 a |
| LED1500 | 228 b | 216 b | 9.9 b | 18.1 b | 151.4 b | 866.4 b | 113.5 a |
| SE | 13.1 | 11.5 | 0.9 | 1.2 | 10.5 | 29.0 | 11.0 |
| Supplementary LED Lighting | Fruit Length (cm) | Fruit Weight (kg) | Juice pH | Acid Content (%) | Thickness of Peel (mm) |
|---|---|---|---|---|---|
| 15 days before harvest | |||||
| Control | 11.01 b | 0.73 c | 5.34 a | 0.55 b | 4.56 a |
| LED 900 | 13.95 a | 1.30 a | 5.50 a | 0.53 a | 4.45 a |
| LED 1500 | 12.31 ab | 0.94 b | 5.48 a | 0.54 ab | 4.84 a |
| SE | 1.11 | 0.10 | 1.01 | 0.10 | 0.00 |
| Harvest stage | |||||
| Control | 13.12 b | 1.14 c | 5.76 a | 0.47 b | 3.66 a |
| LED 900 | 14.25 a | 1.50 a | 5.80 a | 0.40 a | 3.55 a |
| LED 1500 | 13.97 a | 1.30 b | 5.82 a | 0.40 a | 3.95 a |
| SE | 1.25 | 0.12 | 0.56 | 0.00 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.M.; Shibasaki, Y.; Goto, F. Enhancement of Growth and Quality of Winter Watermelon Using LED Supplementary Lighting. Horticulturae 2025, 11, 262. https://doi.org/10.3390/horticulturae11030262
Hossain MM, Shibasaki Y, Goto F. Enhancement of Growth and Quality of Winter Watermelon Using LED Supplementary Lighting. Horticulturae. 2025; 11(3):262. https://doi.org/10.3390/horticulturae11030262
Chicago/Turabian StyleHossain, Md Mukhtar, Yuki Shibasaki, and Fumiyuki Goto. 2025. "Enhancement of Growth and Quality of Winter Watermelon Using LED Supplementary Lighting" Horticulturae 11, no. 3: 262. https://doi.org/10.3390/horticulturae11030262
APA StyleHossain, M. M., Shibasaki, Y., & Goto, F. (2025). Enhancement of Growth and Quality of Winter Watermelon Using LED Supplementary Lighting. Horticulturae, 11(3), 262. https://doi.org/10.3390/horticulturae11030262