
Intercropping Between Panax ginseng and Arisaema amurense Improves Ginseng Quality by Improving Soil Properties and Microbial Communities

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Field Experiment Design
2.3. Sample Collection and Preprocessing
2.4. Measurement Items and Methods
2.4.1. Determination of Soil Physical and Chemical Properties
2.4.2. Determination of Soil Enzyme Activity
2.4.3. High-Throughput Sequencing of Soil Microorganisms
2.4.4. Determination of Ginsenoside Content
2.5. Data Analysis
3. Results
3.1. Effect of Intercropping on Soil pH and Nutrient Content
3.2. Effect of Intercropping on Soil Enzyme Activities
3.3. Effect of Intercropping on Soil Microbial Communities
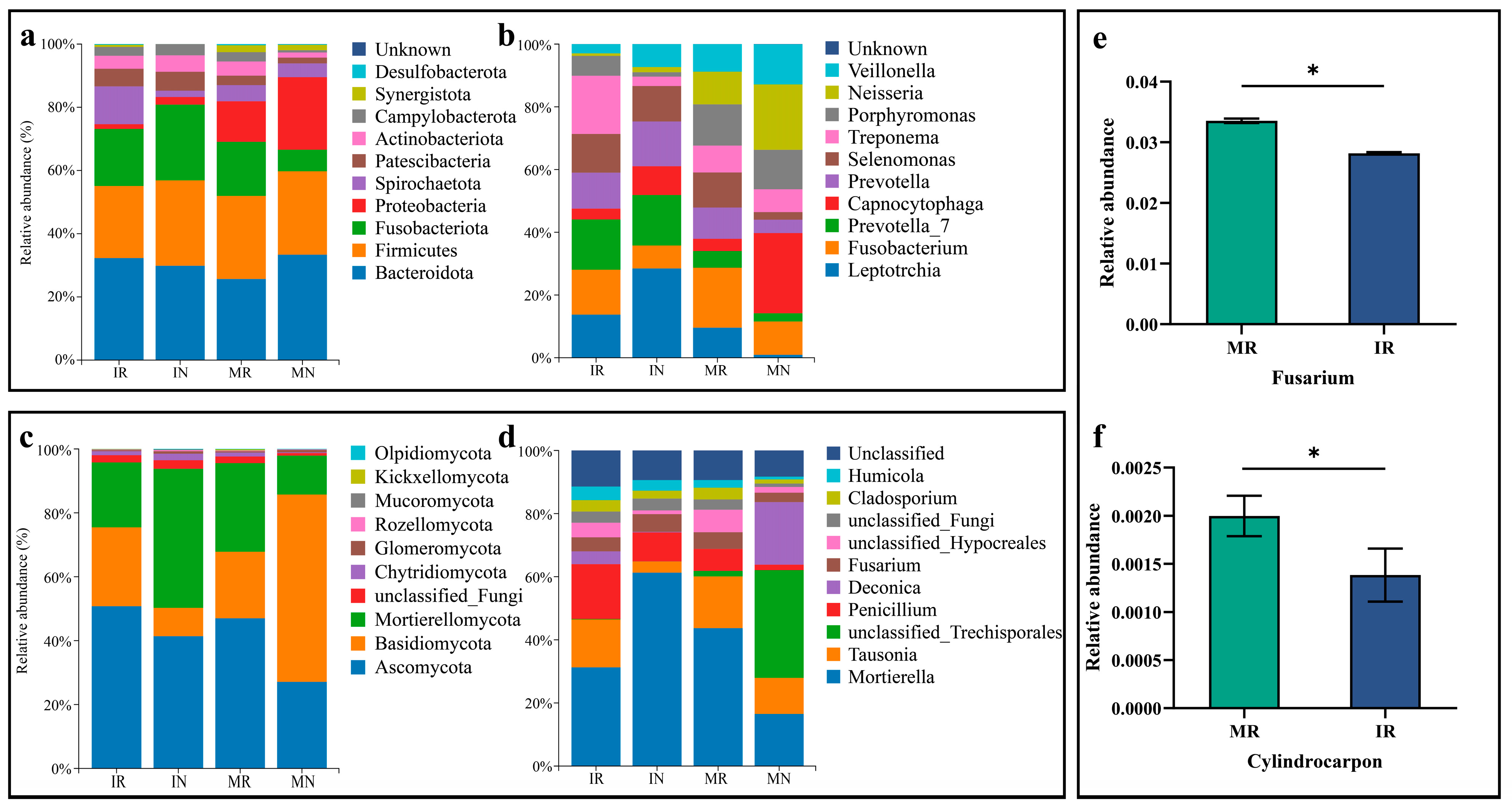
3.3.1. Changes in Relative the Abundance of Species
3.3.2. Changes in the Relative Abundance of Pathogenic Fungi of Ginseng
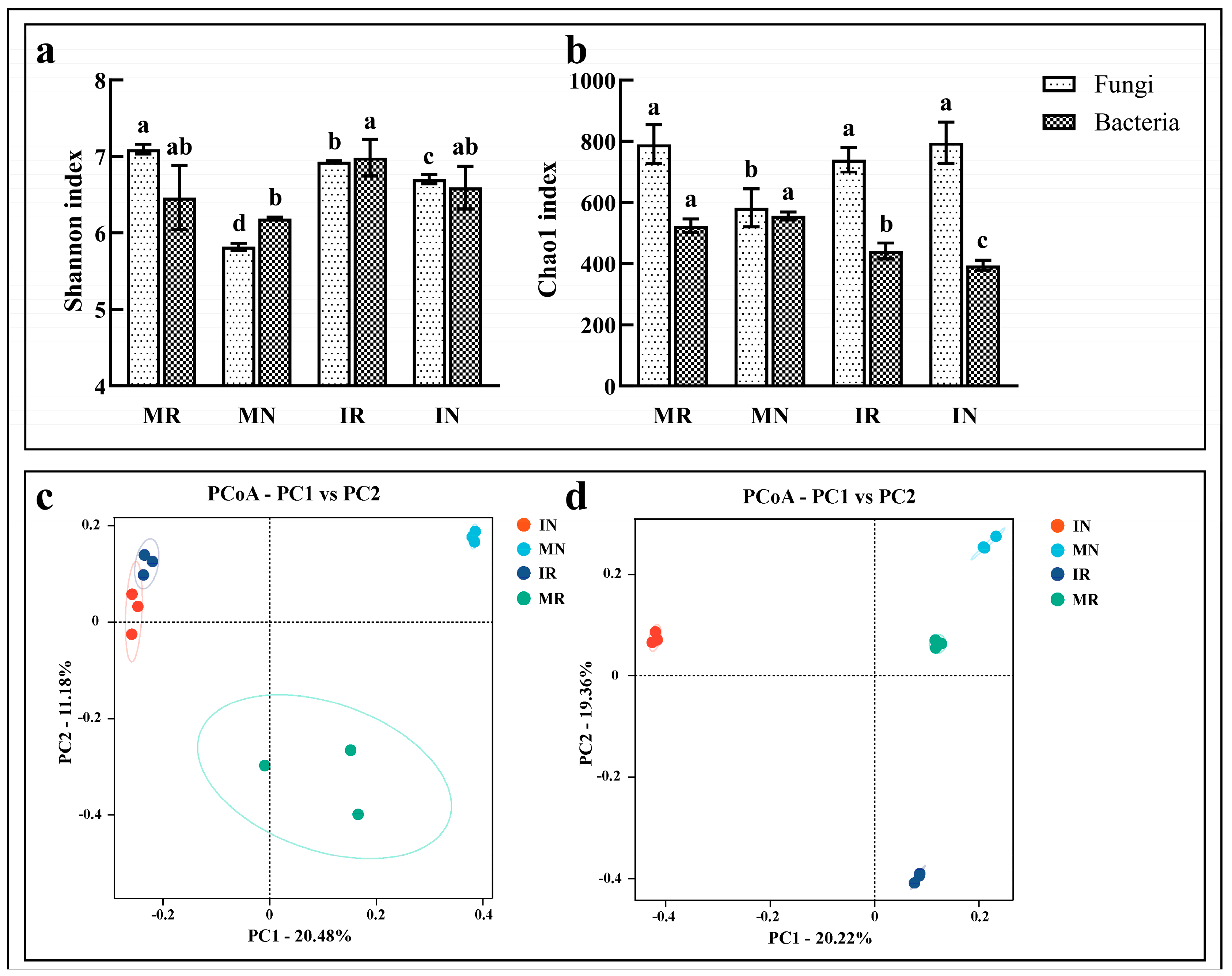
3.3.3. Change in α-Diversity Index
3.3.4. Changes in Microbial Community Structure
3.4. Effect of Intercropping on the Content of Ginsenosides
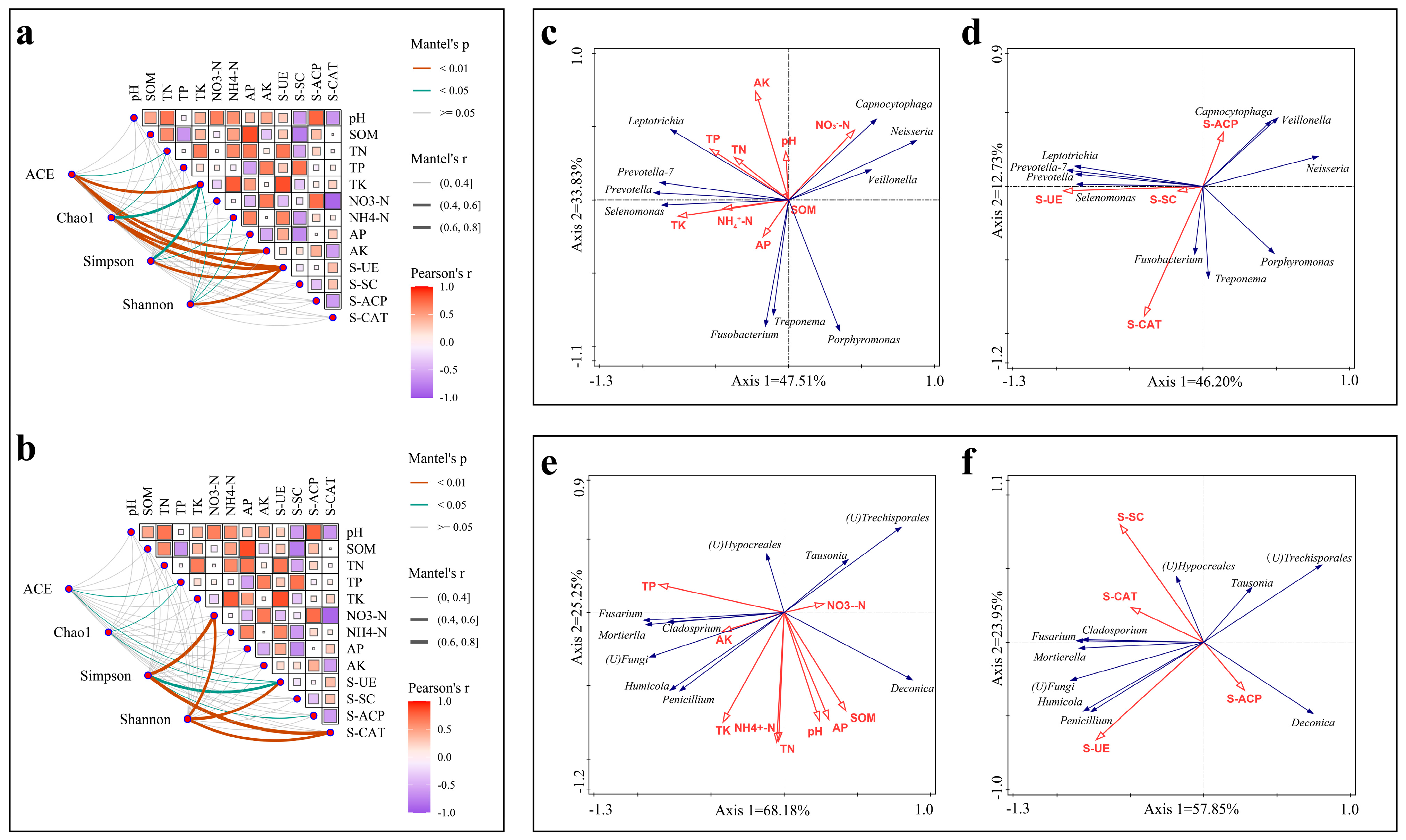
3.5. Correlation Between Soil pH, Nutrient and Soil Microbiology
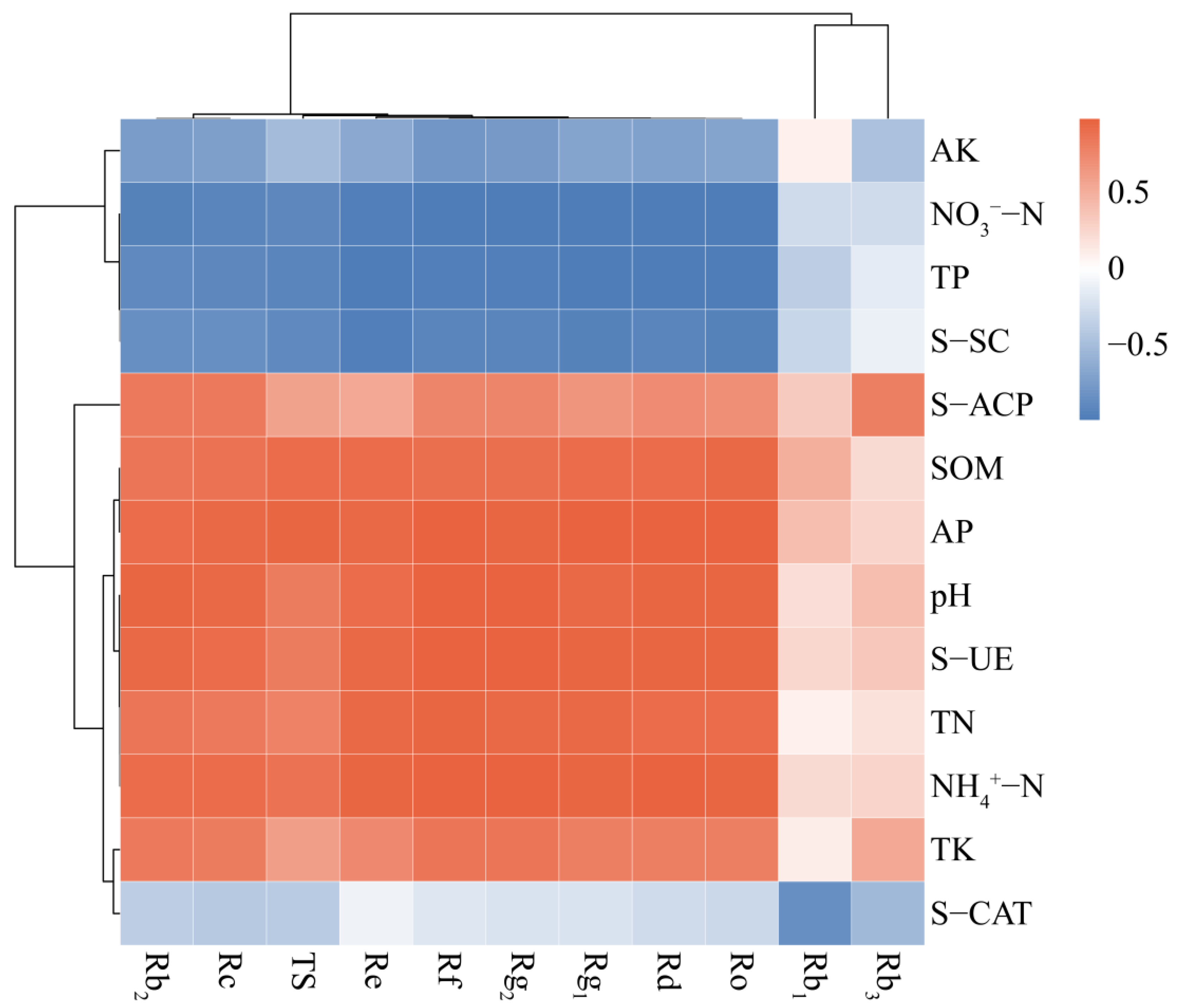
3.6. Correlation Between Soil Factors and Ginsenosides
3.6.1. Correlation of Soil Nutrients, Enzyme Activities and Ginsenosides
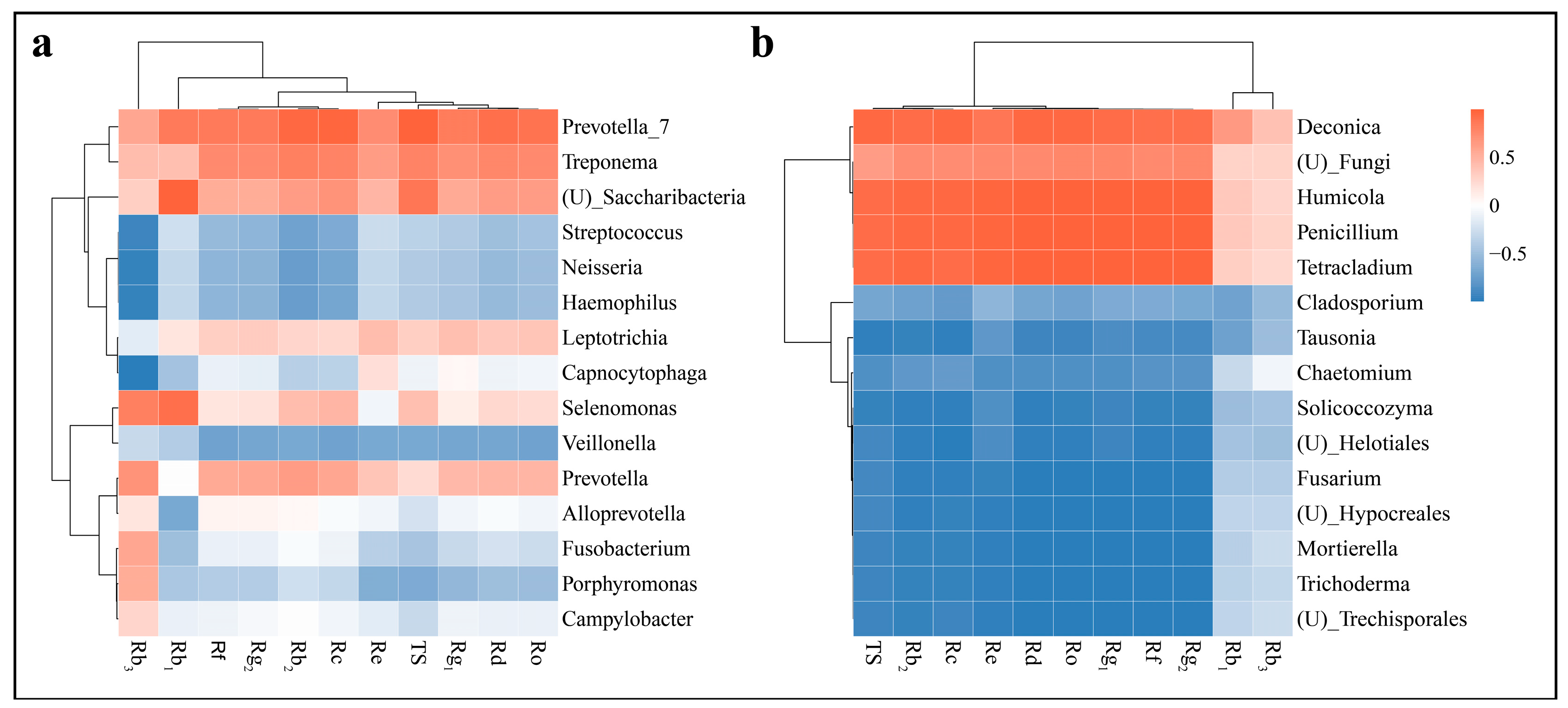
3.6.2. Correlation of Soil Microorganisms with Ginsenosides
4. Discussion
4.1. Effect of Intercropping on Soil Nutrients and Enzymes
4.2. Effect of Intercropping on Rhizosphere Microorganisms
4.3. Intercropping Promotes the Accumulation of Ginsenosides
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, B.L.; Sun, X.B.; Zhao, X.J. Panax Ginseng, 1st ed.; World Publishing Corporation: Beijing, China, 2023; pp. 3–12. [Google Scholar]
- Zhang, J.X.; Zhou, D.P.; Yuan, X.Q.; Xu, Y.H.; Chen, C.B.; Zhao, L. Soil Microbiome and Metabolome Analysis Reveals Beneficial Effects of Ginseng–celandine Rotation on the Rhizosphere Soil of Ginseng-used Fields. Rhizosphere 2022, 23, 100559. [Google Scholar] [CrossRef]
- Xiao, C.P.; Yang, L.M.; Zhang, L.X.; Liu, C.J.; Han, M. Effects of Cultivation Ages and Modes on Microbial Diversity in the Rhizosphere Soil of Panax ginseng. J. Ginseng Res. 2016, 40, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Feng, S.; Lu, B.A.; Yang, L.N.; Wang, X.; Zhang, Y.J.; Gao, J. Fusarium oxysporum f. sp. ginseng, A New forma specialis Causing Fusarium Root Rot of Panax ginseng. Phytopathol. Mediterr. 2022, 61, 417–429. [Google Scholar] [CrossRef]
- Li, Z.B.; Sun, C.L.; Liu, X.R.; Zhou, R.J. Molecular Identification and Analysis on Differential Pathogenicity of Cylindrocarpon Species Associated with Ginseng Root Rust Rot in Northeastern China. Front. Plant Sci. 2022, 13, 894104. [Google Scholar] [CrossRef] [PubMed]
- Li, Q. Effect of Ginsenoside on Chemotaxis Aggravation of Soil Rust Rot in Continuous Cropping Soil and Its Mechanism. Ph.D. Thesis, Jilin Agricultural University, Changchun, China, 2020. [Google Scholar]
- Xu, Z.Y. Study on Effects of Ginseng Root Exudates to Chemotaxis of Its Pathogens and Allelopathy to Seeds and Seedlings of Panax. Master’s Thesis, Jilin Agricultural University, Changchun, China, 2021. [Google Scholar]
- Shen, Y.L.; Zhang, H.; Zhan, Y.; Zhang, T.; Li, Q.; Wang, E.P.; Chen, C.B. Transcriptomics and Metabolomics of the Molecular Mechanisms of Ginseng’s Response to the Continuous Cropping Obstacle. Physiol. Mol. Plant Pathol. 2024, 133, 102329. [Google Scholar] [CrossRef]
- Wang, X.Y.; Shen, B.Y.; Sun, L.F.; Song, G.Z.; Jia, H.Q.; Sun, W.S. Development and Advantages of Ecological Planting Patterns for Dictamnus dasycarpus. Hortic. Seed 2023, 43, 22–32. [Google Scholar]
- Yang, L.M. Theory and Technology Frontiers of Ecological Planting of Chinese Medicinal Materials. J. Jilin Agric. Univ. 2020, 42, 355–363. [Google Scholar]
- Bian, X.B.; Yang, X.H.; Zhang, K.X.; Zhai, Y.R.; Li, Q.; Zhang, L.X.; Sun, X. Potential of Medicago sativa and Perilla frutescens for Overcoming the Soil Sickness Caused by Ginseng Cultivation. Front. Microbiol. 2023, 14, 1134331. [Google Scholar] [CrossRef]
- Bian, X.B.; Yang, X.H.; Li, Q.; Sun, X. Effects of Planting of Two Common Crops, Allium fistulosum and Brassica napus, on Soil Properties and Microbial Communities of Ginseng Cultivation in Northeast China. BMC Microbiol. 2022, 22, 182. [Google Scholar] [CrossRef]
- Gao, S.H.; Lv, C.J. Advances in the Mechanism of Intercropping to Alleviate Continuous Cropping Obstacles in Plants. Hortic. Seed 2023, 43, 98–100. [Google Scholar]
- Huang, L.Q.; Guo, L.P. Ecological Agriculture of Chinese Materia Medica, 2nd ed.; Shanghai Scientific & Technical Publishers: Shanghai, China, 2022; pp. 81–87. [Google Scholar]
- Hang, Y.; Luo, F.L.; Zhao, Z.; Tang, C.L.; Lin, J.; Li, Q.W.; Deng, C. Effect of Pinellia ternata with Different Intercropping Crops on Soil Microorganism, Nutrient and Enzyme Activity. J. Chin. Med. Mater. 2018, 41, 1522–1528. [Google Scholar]
- Zhang, X.H. Studies on Mechanism of Continuous Angelica sinensis Cropping Obstacle and Its Preparatory Bioremediation. Ph.D. Thesis, Gansu Agricultural University, Lanzhou, China, 2009. [Google Scholar]
- Lyu, B.C.; Sun, H.; Qian, J.Q.; Liang, H.; Zhu, J.P.; Zhang, Q.S.; Shao, C.; Zhang, Y.Y. Interaction between Root Exudates of Medicinal Plants and Rhizosphere Microorganisms and Its Application in Ecological Planting of Chinese Medicinal Materials. China J. Chin. Mater. Med. 2024, 49, 2128–2137. [Google Scholar]
- Zhou, Y.F.; Feng, R.; Tang, L. Research Progress on the Root Exudate-mediated Mechanism of Intercropping for Prevention and Control of Soil-borne Diseases. J. Yunnan Agric. Univ. Nat. Sci. 2023, 38, 353–360. [Google Scholar]
- Iwamoto, K.; Kawamoto, H.; Takeshita, F.; Matsumura, S.; Ayaki, I.; Moriyama, T.; Zaima, N. Mixing Ginkgo biloba Extract with Sesame Extract and Turmeric Oil Increases Bioavailability of Ginkgolide A in Mice Brain. J. Oleo Sci. 2019, 68, 923–930. [Google Scholar] [CrossRef]
- Zeng, J.R.; Liu, J.Z.; Lu, C.H.; Ou, X.H.; Luo, K.K.; Li, C.M.; He, M.L.; Zhang, H.Y.; Yan, H.J. Intercropping with Turmeric or Ginger Reduce the Continuous Cropping Obstacles That Affect Pogostemon cablin (Patchouli). Front. Microbiol. 2020, 11, 579719. [Google Scholar] [CrossRef]
- Hao, H.P.; Bai, H.T.; Xia, F.; Hao, Y.P.; Li, H.; Cui, H.X.; Xie, X.M.; Shi, L. Effects of Tea-Litsea cubeba Intercropping on Soil Microbial Community Structure in Tea Plantation. Sci. Agric. Sin. 2021, 54, 3959–3969. [Google Scholar]
- Zhou, X.G.; Zhang, J.Y.; Khashi u Rahman, M.; Gao, D.M.; Wei, Z.; Wu, F.Z.; Dini-Andreote, F. Interspecific Plant Interaction Via Root Exudates Structures the Disease Suppressiveness of Rhizosphere Microbiomes. Mol. Plant 2023, 16, 849–864. [Google Scholar] [CrossRef] [PubMed]
- Du, M.X.; Qiu, D.Y.; Ren, F.Y.; Wang, S.J.; Hu, F.D. Effects of Intercropping Crops and Stubbles on Growth, Yield and Quality of Codonopsis pilosula. Acta Bot. Boreali-Occident. Sin. 2021, 41, 1884–1892. [Google Scholar]
- Duan, W.Y.; Chen, X.L.; Ran, Z.F.; Fang, L.; Song, Z.J.; Guo, L.P.; Zhou, J. Intercropping History, Pattern, and Case Analysis of Traditional Chinese Materials. China J. Chin. Mater. Med. 2024, 58, 104. [Google Scholar]
- Chinese Pharmacopoeia Commission. Herbs and tablets. In Chinese Pharmacopeia, 1st ed.; China Medical Science and Technology Press: Beijing, China, 2020; pp. 58–59. [Google Scholar]
- Wang, M. Comparative Analysis of Effective Ingredients and Antioxidant Activity of Arisaema amurense Maxim. Master’s Thesis, Yanbian University, Yanji, China, 2021. [Google Scholar]
- Wang, L.W.; Xu, B.G.; Wang, J.Y.; Su, Z.Z.; Lin, F.C.; Zhang, C.L.; Kubicek, C.P. Bioactive Metabolites from Phoma Species, an Endophytic Fungus from the Chinese Medicinal Plant Arisaema erubescens. Appl. Microbiol. Biotechnol. 2012, 93, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Gong, M.F.; Wu, S.L.; Fu, Q.C.; Yan, J.J.; Xu, G.Y. Analysis of Bacteriostatic Activity of Ethanol Extract from Plants in Araceae on Plant Pathogenic Fungi. Genom. Appl. Biol. 2016, 35, 152–157. [Google Scholar]
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2015; pp. 70–114. [Google Scholar]
- Li, Y.J.; Xu, G.H.; Yu, Y. Freeze-thaw Aged Polyethylene and Polypropylene Microplastics Alter Enzyme Activity and Microbial Community Composition in Soil. J. Hazard. Mater. 2024, 470, 134249. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; O’Connor, T.K.; Arnold, H.K.; Hubbell, S.P.; Wright, S.J.; Green, J.L. Relationships between Phyllosphere Bacterial Communities and Plant Functional Traits in a Neotropical Forest. Proc. Natl. Acad. Sci. USA 2014, 111, 13715–13720. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Chinese Pharmacopoeia Commission. Vegetable oils and extracts. In Chinese Pharmacopeia, 1st ed.; China Medical Science and Technology Press: Beijing, China, 2020; pp. 409–410. [Google Scholar]
- Zhang, H.; Yang, H.; Wang, Y.P.; Gao, Y.G.; Zhang, L.X. The Response of Ginseng Grown on Farmland to Foliar-applied Iron, Zinc, Manganese and Copper. Ind. Crops Prod. 2013, 45, 388–394. [Google Scholar] [CrossRef]
- Wu, L.K.; Wu, H.M.; Zhu, Q.; Chen, J.; Wang, J.Y.; Wu, Y.H.; Lin, H.; Lin, W.X. Effects of Different Amendments on Contents of Phenolic Acids and Specific Microbes in Rhizosphere of Pseudostellaria heterophylla. Chin. J. Appl. Ecol. 2016, 27, 3623–3630. [Google Scholar]
- Xing, H.Q. The Effects of Intercropping Legume Green Manure in Tea Plantation on Soil Properties and Tea Quality. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2020. [Google Scholar]
- Zhang, J.G.; Fan, S.H.; Qin, J.; Dai, J.C.; Zhao, F.J.; Gao, L.Q.; Lian, X.H.; Shang, W.J.; Xu, X.M.; Hu, X.P. Changes in the Microbiome in the Soil of an American Ginseng Continuous Plantation. Front. Plant Sci. 2020, 11, 572199. [Google Scholar] [CrossRef] [PubMed]
- Yu, X. Study on the Variation Characteristics of Crop Yield, Carbon and Nitrogen Accumulation and Soil Nutrients under Sweet Corn || Soybean Intercropping System in Upland Red Soil. Master’s Thesis, Jiangxi Agricultural University, Nanchang, China, 2021. [Google Scholar]
- Chen, X.; Tian, Y.; Guo, X.F.; Chen, G.K.; He, H.Z.; Li, H.S. The Effect of Monoculture Peanut and Cassava/Peanut Intercropping on Physical and Chemical Properties in Peanut Rhizosphere Soil under the Biochar Application and Straw Mulching. IOP Conf. Ser. Earth Environ. Sci. 2017, 59, 012021. [Google Scholar] [CrossRef]
- Cong, W.F.; Hoffland, E.; Li, L.; Six, J.; Sun, J.X.; Bao, X.G.; Zhang, F.S.; Van Der Werf, W. Intercropping Enhances Soil Carbon and Nitrogen. Glob. Chang. Biol. 2015, 21, 1715–1726. [Google Scholar] [CrossRef]
- Cesco, S.; Mimmo, T.; Tonon, G.; Tomasi, N.; Pinton, R.; Terzano, R.; Neumann, G.; Weisskopf, L.; Renella, G.; Landi, L.; et al. Plant-borne Flavonoids Released into the Rhizosphere: Impact on Soil Bio-activities Related to Plant Nutrition. A Review. Biol. Fertil. Soils 2012, 48, 123–149. [Google Scholar] [CrossRef]
- Jing, C.L.; Xu, Z.C.; Zou, P.; Tang, Q.; Li, Y.Q.; You, X.W.; Zhang, C.S. Coastal Halophytes Alter Properties and Microbial Community Structure of the Saline Soils in the Yellow River Delta, China. Appl. Soil Ecol. 2019, 134, 1–7. [Google Scholar] [CrossRef]
- Li, Q.S.; Wu, L.K.; Chen, J.; A Khan, M.; Luo, X.M.; Lin, W.X. Biochemical and Microbial Properties of Rhizospheres under Maize/Peanut Intercropping. J. Integr. Agric. 2016, 15, 101–110. [Google Scholar] [CrossRef]
- Wu, X.; Hu, Y.C.; Yang, F.J.; Su, C.J.; Wang, X.Z.; Wu, F.Z. Effects of Intercropping Tomato with Potato Onion on Nitrogen Absorption and Rhizosphere Microbial Diversity of Tomato. J. Plant Nutr. Fert. 2022, 28, 1478–1493. [Google Scholar]
- Wang, G.W.; Jin, Z.X.; Xinxin Wang, X.X.; George, T.S.; Feng, G.; Zhang, L. Simulated Root Exudates Stimulate the Abundance of Saccharimonadales to Improve the Alkaline Phosphatase Activity in Maize Rhizosphere. Appl. Soil Ecol. 2022, 170, 104274. [Google Scholar] [CrossRef]
- Yang, F.; Liao, D.P.; Wu, X.L.; Gao, R.C.; Fan, Y.F.; Ali Raza, M.; Wang, X.C.; Yong, T.W.; Liu, W.G.; Liu, J.; et al. Effect of Aboveground and Belowground Interactions on the Intercrop Yields in Maize-soybean Relay Intercropping Systems. Field Crops Res. 2017, 203, 16–23. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The Rhizosphere Microbiome and Plant Health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Chialva, M.; Lanfranco, L.; Bonfante, P. The Plant Microbiota: Composition, Functions, and Engineering. Curr. Opin. Biotechnol. 2022, 73, 135–142. [Google Scholar] [CrossRef]
- Zhao, G.X.; Yang, H.T.; Shao, X.B.; Cui, Z.H.; Liu, H.G.; Zhang, J. Phosphate-solubilizing Properties and Optimization of Cultivation Conditions of Penicillium rubens: A Highly Efficient Phosphate Solubilizer. Biotechnol. Bull. 2023, 39, 71–83. [Google Scholar]
- Li, H.R.; Lin, W.; Wang, T.Y.; Lei, J.; Chen, P. Research Progress on Allelopathic Effects of Ginseng. Mol. Plant Breed. 2024, 22, 1254002. [Google Scholar]
- Qu, Y.T.; Zhang, Q.Q.; Yu, Y.F.; Sayikal, D.; Cai, L.L.; Zhang, S.J.; Li, Y.F.; Li, Y.C. Research Advances on Mechanisms and Preventions of the Medicinal Plants’ Continuous Cropping Obstacles from the Perspective of Rhizosphere Microecology. J. Zhejiang Univ. Agric. Life Sci. 2022, 48, 403–414. [Google Scholar]
- Ma, H.Y.; Surigaoge, S.; Xu, Y.; Li, Y.C.; Christie, P.; Zhang, W.P.; Li, L. Responses of soil microbial community diversity and co-occurrence networks to interspecific interactions in soybean/maize and peanut/maize intercropping systems. Appl. Soil Ecol. 2024, 203, 105613. [Google Scholar] [CrossRef]
- Trochine, A.; Bellora, N.; Nizovoy, P.; Duran, R.; Greif, G.; de García, V.; Batthyany, C.; Robello, C.; Libkind, D. Genomic and Proteomic Analysis of Tausonia pullulans Reveals a Key Role for a GH15 Glucoamylase in Starch Hydrolysis. Appl. Microbiol. Biotechnol. 2022, 106, 4655–4667. [Google Scholar] [CrossRef] [PubMed]
- Ogundeji, A.O.; Li, Y.; Liu, X.J.; Meng, L.B.; Sang, P.; Mu, Y.; Wu, H.L.; Ma, Z.N.; Hou, J.; Li, S.M. Eggplant by Grafting Enhanced with Biochar Recruits Specific Microbes for Disease Suppression of Verticillium Wilt. Appl. Soil Ecol. 2021, 163, 103912. [Google Scholar] [CrossRef]
- Paulus, J.K.; Kourelis, J.; Ramasubramanian, S.; Homma, F.; Godson, A.; Hörger, A.C.; Hong, T.N.; Krahn, D.; Carballo, L.O.; Wang, S.S.; et al. Extracellular Proteolytic Cascade in Tomato Activates Immune Protease Rcr3. Proc. Natl. Acad. Sci. USA 2020, 117, 17409–17417. [Google Scholar] [CrossRef]
- Wu, H.M.; Lin, M.H.; Rensing, C.; Qin, X.J.; Zhang, S.K.; Chen, J.; Wu, L.K.; Zhao, Y.L.; Lin, S.; Lin, W.X. Plant-mediated Rhizospheric Interactions in Intraspecific Intercropping Alleviate the Replanting Disease of Radix pseudostellariae. Plant Soil 2020, 454, 411–430. [Google Scholar] [CrossRef]
- Wang, P.; Zhu, L.X.; Chen, X.X.; Feng, H.; Sun, W.S.; Qin, N. Effects of Platycodon grandiflorum and Allium fistulosum Intercropping on Soil Nutrients, Microorganism and Enzyme Activity. J. Plant Nutr. Fert. 2018, 24, 668–675. [Google Scholar]
- Yan, L.; Riaz, M.; Liu, J.Y.; Yu, M.; Jiang, C.C. The Aluminum Tolerance and Detoxification Mechanisms in Plants; Recent Advances and Prospects. Crit. Rev. Environ. Sci. Technol. 2022, 52, 1491–1527. [Google Scholar] [CrossRef]
- Bian, X.B.; Xiao, S.Y.; Zhao, Y.; Xu, Y.H.; Yang, H.; Zhang, L.X. Comparative Analysis of Rhizosphere Soil Physiochemical Characteristics and Microbial Communities between Rusty and Healthy Ginseng Root. Sci. Rep. 2020, 10, 15756. [Google Scholar] [CrossRef]
- Accoe, F.; Boeckx, P.; Busschaert, J.; Hofman, G.; Van Cleemput, O. Gross N Transformation Rates and Net N Mineralisation Rates Related to the C and N Contents of Soil Organic Matter Fractions in Grassland Soils of Different Age. Soil Biol. Biochem. 2004, 36, 2075–2087. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, Y.Y.; Sun, C.W.; Zheng, P.H.; Gao, M.; Zhang, L.N. Relationship between Soil Nutrient Status and Ginsenosides Content in Different Growth Patterns of Panax ginseng. Acta Agric. Boreali-Occident. Sin. 2012, 21, 146–152. [Google Scholar]
- Ming, Q.L.; Han, T.; Huang, F.; Qin, L.P. Advances in Studies on Ginsenoside Biosynthesis and Its Related Enzymes. Chin. Tradit. Herb. Drugs 2010, 41, 1913–1917. [Google Scholar]
- Chen, W.; Gao, W.Y.; Jia, W.; Duan, H.Q.; Xiao, P.G. Advances in Studies on Tissue and Cell Culture in Medicinal Plants of Panax L. Chin. Tradit. Herb. Drugs 2005, 36, 616–620. [Google Scholar]
- Ma, W.Q.; Wang, H.Y.; Wang, S.; Wan, X.F.; Kang, C.Z.; Guo, L.P. Effects of Ecological Factors on Shape and Ginsenoside of Panax ginseng. China J. Chin. Mater. Med. 2021, 46, 1920–1926. [Google Scholar]
- Yang, Y.W.; Jiang, Y.T. Study on Relationship between Effective Components and Soil Enzyme Activity in Different Growth Patterns of Panax ginseng. China J. Chin. Mater. Med. 2016, 41, 2987–2992. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | pH | SOM (g/kg) | Total Nutrient (g/kg) | Available Nutrient (mg/kg) | |||||
|---|---|---|---|---|---|---|---|---|---|
| TN | TP | TK | NO3−-N | NH4+-N | AP | AK | |||
| MR | 5.24 ± 0.02 c | 20.42 ± 1.92 c | 1.42 ± 0.10 b | 1.02 ± 0.03 a | 41.12 ± 1.01 bc | 34.10 ± 0.10 c | 2.96 ± 0.08 c | 100.03 ± 1.44 b | 81.60 ± 1.39 b |
| MN | 5.35 ± 0.03 ab | 24.84 ± 2.80 ab | 1.56 ± 0.13 ab | 0.81 ± 0.04 b | 40.67 ± 0.02 c | 92.71 ± 1.47 a | 4.07 ± 0.18 bc | 102.80 ± 4.72 b | 82.40 ± 2.40 b |
| IR | 5.33 ± 0.02 b | 27.98 ± 2.16 a | 1.88 ± 0.13 a | 0.84 ± 0.02 b | 43.25 ± 0.88 a | 25.27 ± 0.57 d | 6.67 ± 1.04 a | 109.07 ± 5.79 a | 79.20 ± 1.39 b |
| IN | 5.38 ± 0.03 a | 22.71 ± 0.32 bc | 1.77 ± 0.25 a | 1.08 ± 0.04 a | 42.72 ± 0.87 ab | 90.56 ± 0.67 b | 5.78 ± 2.28 ab | 101.00 ± 2.66 b | 91.19 ± 1.39 a |
| Sample | S-UE (μg/g·d−1) | S-SC (mg/g·d−1) | S-ACP (μmol/g·d−1) | S-CAT (mmol/g·d−1) |
|---|---|---|---|---|
| MR | 197.055 ± 6.249 b | 8.125 ± 0.591 a | 22.143 ± 0.402 b | 6.378 ± 0.056 a |
| MN | 176.581 ± 1.482 c | 6.611 ± 0.303 bc | 23.248 ± 0.504 a | 5.631 ± 0.198 b |
| IR | 235.384 ± 7.787 a | 6.354 ± 0.075 c | 22.632 ± 0.098 ab | 6.323 ± 0.204 a |
| IN | 226.115 ± 5.179 a | 7.179 ± 0.225 b | 23.252 ± 0.491 a | 5.761 ± 0.192 b |
| Sample | MG | IG | |
|---|---|---|---|
| total saponins (TS)/% | 4.321 ± 0.056 | 4.930 ± 0.200 * | |
| Ginsenosides/(g/kg) | Rb1 | 2.147 ± 0.013 | 2.185 ± 0.084 |
| Rg1 | 1.941 ± 0.027 | 2.261 ± 0.011 * | |
| Re | 4.087 ± 0.020 | 4.181 ± 0.009 * | |
| Rb2 | 1.554 ± 0.044 | 1.739 ± 0.018 * | |
| Rb3 | 1.629 ± 0.057 | 1.651 ± 0.018 | |
| Rc | 1.820 ± 0.055 | 2.071 ± 0.041 * | |
| Rd | 1.039 ± 0.005 | 1.445 ± 0.022 * | |
| Rf | 0.620 ± 0.006 | 0.721 ± 0.005 * | |
| Rg2 | 0.387 ± 0.003 | 0.430 ± 0.002 * | |
| Ro | 0.645 ± 0.003 | 0.901 ± 0.016 * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, B.; Sun, H.; Cao, W.; Zhu, J.; Liang, H.; Long, H.; Cui, Y.; Shao, C.; Zhang, Y. Intercropping Between Panax ginseng and Arisaema amurense Improves Ginseng Quality by Improving Soil Properties and Microbial Communities. Horticulturae 2025, 11, 172. https://doi.org/10.3390/horticulturae11020172
Lv B, Sun H, Cao W, Zhu J, Liang H, Long H, Cui Y, Shao C, Zhang Y. Intercropping Between Panax ginseng and Arisaema amurense Improves Ginseng Quality by Improving Soil Properties and Microbial Communities. Horticulturae. 2025; 11(2):172. https://doi.org/10.3390/horticulturae11020172
Chicago/Turabian StyleLv, Bochen, Hai Sun, Weiyu Cao, Jiapeng Zhu, Hao Liang, Hongjie Long, Yanmei Cui, Cai Shao, and Yayu Zhang. 2025. "Intercropping Between Panax ginseng and Arisaema amurense Improves Ginseng Quality by Improving Soil Properties and Microbial Communities" Horticulturae 11, no. 2: 172. https://doi.org/10.3390/horticulturae11020172
APA StyleLv, B., Sun, H., Cao, W., Zhu, J., Liang, H., Long, H., Cui, Y., Shao, C., & Zhang, Y. (2025). Intercropping Between Panax ginseng and Arisaema amurense Improves Ginseng Quality by Improving Soil Properties and Microbial Communities. Horticulturae, 11(2), 172. https://doi.org/10.3390/horticulturae11020172