Abstract
Zinc oxide nanoparticles (ZnO NPs) have garnered increasing attention in agriculture due to their potential to enhance plant growth and nutrient use. This research investigates the concentration-dependent effects of ZnO NPs on young crabapple (Malus robusta) plants, addressing gaps in understanding how different concentrations influence plant development. A hydroponic experiment was conducted, applying foliar treatments of 200 mg L−1 ZnSO4 (S200) and 200, 500, and 1000 mg L−1 ZnO NPs (N200, N500, N100). The control group (CK) was treated with deionized water (dH2O). Growth parameters, antioxidant enzyme activity, and nutrient contents were measured to evaluate the impact of ZnO NPs on plant development and nutrient uptake. The results showed that N200 enhanced growth, increasing plant height by 22.64%, total dry weight by 49.36%, and root length by 116.07%. In contrast, N500 and N1000 induced oxidative stress, elevating H2O2 and MDA by 32.02~54.43% and inhibiting growth. N200 also improved nutrient uptake, increasing K, Ca, Fe, and Zn uptake fluxes by 84.92%, 112.12%, 185.15%, and 149.92%, respectively, whereas N1000 suppressed overall nutrient uptake but increased root Ca accumulation by 64.59%. These findings suggest that ZnO NPs can enhance plant growth and nutrient utilization at low concentrations, with potential implications for agricultural practices involving nanoparticle (NP)-based fertilizers.
Keywords:
ZnO NPs; hydroponics; foliar application; growth; oxidative stress; root architecture; nutrient use; crabapple 1. Introduction
Crop fertilization is critical for supplying the necessary nutrients required by plants to achieve optimal growth and fruit development. Mineral nutrition, which encompasses the absorption, processing, and utilization of essential elements, plays a fundamental role in maximizing a plant’s biological potential. Fertilization, a key factor influencing crop productivity, is often associated with inefficiencies, as plants typically absorb only 10–60% of the applied fertilizer [1]. This inefficiency not only results in economic losses, but also causes environmental concerns such as nutrient leaching, soil degradation, and groundwater pollution [1]. Consequently, there is growing interest in developing more efficient and environmentally friendly alternatives to traditional fertilization methods, with nano-fertilizers emerging as a promising solution [2].
Nano-fertilizers, which incorporate NPs to deliver essential nutrients, offer numerous advantages over conventional fertilizers [2]. Due to their small size (1–100 nm), NPs exhibit unique properties, such as high surface area, enhanced solubility, and increased reactivity, which improve nutrient delivery efficiency [3]. The enhanced bioavailability and faster uptake of nutrients by plants result in a more efficient fertilization process, reducing the need for large quantities of fertilizers and minimizing environmental impact [4]. Nanofertilizers provide nutrients to plants in two ways: directly and indirectly. Directly, they release essential elements in ionic (e.g., Cu2+) or NPs forms (e.g., Cu or CuO) when they interact with the soil. Indirectly, they enhance nutrient availability by either triggering physicochemical reactions in the soil or improving plant nutrient uptake through changes in plant metabolism and microbiome [5,6]. In addition to improving nutrient delivery, nano-fertilizers can be designed to release nutrients in a controlled and gradual manner, ensuring a steady supply to plants and reducing the risk of nutrient leaching [7,8].
One of the key advantages of nano-fertilizers is their application via foliar spray. Foliar fertilization allows for direct application of nutrients to the aerial parts of the plant, which leads to faster absorption compared to soil applications [9]. This method is especially effective for delivering micronutrients such as zinc (Zn), which is essential for various physiological processes, including enzyme activity, protein synthesis, and photosynthesis [10]. The benefits of foliar nano-fertilizers have been demonstrated in various fruit trees. The foliar application of ZnO NPs in apples, for example, has been shown to improve fruit quality, enhance stress tolerance, and increase antioxidant levels [11]. Similarly, the use of ZnO NPs in mangoes, pomegranates, and olives has resulted in improved growth, higher yields, and better fruit quality [7,12,13]. The application of nano-fertilizers has also been associated with improvements in the nutraceutical quality of fruits. For instance, the foliar application of ZnO NPs and copper NPs has increased the levels of antioxidants and bioactive compounds in fruits such as habanero peppers, tomatoes, and strawberries [14,15]. These improvements not only enhance the nutritional value of the fruit, but also contribute to better postharvest quality, such as increased shelf life and reduced spoilage [16,17].
In addition to improving the quality and quantity of fruit, nano-fertilizers have been found to enhance the resistance of fruit trees to various abiotic stresses, such as water deficit and salinity, as well as biotic stresses like plant diseases [16,17,18]. For example, ZnO NPs have been shown to increase water-use efficiency and improve tolerance to drought conditions in strawberry [19]. Similarly, the application of ZnO NPs can help apple rootstock M9-T337 resist salinity stress, which is becoming an increasingly common issue due to climate change [2]. These effects are attributed to the ability of nano-fertilizers to induce antioxidant activity and other protective mechanisms in plants, helping them cope with environmental stressors [20].
However, ZnO NPs exhibit concentration-dependent effects. Low concentrations can enhance plant growth, chlorophyll content, and antioxidant activity, while high concentrations may induce toxicity, reducing root growth, and nutrient uptake [21,22,23,24]. NPs toxicity can occur directly through plant absorption or adsorption, or indirectly via the release of ions during NPs breakdown or by triggering enhanced responses to other environmental toxins [5]. These dual effects have been observed across various crops, including legumes, vegetables, and cereals [25,26,27,28], highlighting the need for dosage optimization. Based on these findings, we propose the following testable hypotheses: (1) Low ZnO NP doses enhance root architecture and nutrient flux without oxidative stress; (2) Appropriate ZnO NPs improve root Zn accumulation more effectively than ZnSO4 due to their enhanced uptake pathway.
Apples, a globally important fruit crop, are particularly sensitive to micronutrient availability, including Zn [29], as Zn deficiency limits growth and yield, while foliar Zn (especially nano-Zn) application can increase yield by 7~11% and reduce fruit drop by up to 30% [30]. Despite growing interest in ZnO NPs in agriculture, their specific effects on apple young plant growth and nutrient absorption remain unclear. This study aims to fill this gap by evaluating the impact of foliar-applied ZnO NPs, at varying concentrations, on apple young plant growth, antioxidant enzyme activity, and mineral nutrient content under hydroponic conditions. The findings will provide theoretical support for using ZnO NPs as nano-fertilizers in apple cultivation, contributing to more efficient and sustainable agricultural practices.
2. Materials and Methods
2.1. Materials and Treatment
ZnO NPs used in this experiment had a particle size of 30 nm and purity > 99.9%, provided by Macklin Biochemical Co., Ltd. (#Z820772, Shanghai, China). Detailed nanoparticle characterization, including SEM and TEM morphology, zeta potential, and hydrodynamic size, has been previously reported [31]. For experimental preparation, they were suspended in dH2O and stirred for 2 h using a mechanical stirrer. The suspension was then treated with an ultrasonic bath (20 kHz) for 30 min to ensure uniform dispersion of the particles.
One-year-old young crabapple (Malus robusta) plants were used for the experiment. In January 2022, apple seeds were cold-stratified in sand and germinated in the greenhouse of the Horticulture Institute, Shanxi Academy of Agricultural Sciences. Germinated seeds were sown in 30 aperture disks (5 cm × 5 cm, 32 holes per disk) filled with vermiculite, with 1–2 seeds per hole. Young plants were regularly watered to maintain proper substrate moisture. The experiment commenced when plants developed 8–10 true leaves. Young plants with uniform growth were transplanted into hydroponic pots (43 cm × 30 cm × 14 cm) containing 18 L nutrient solution. The solution pH was adjusted to 6.0 ± 0.2 using phosphoric acid, and oxygen levels were maintained using an air pump and air stones. Young plants were first acclimated in half-strength Hoagland solution for 15 d, followed by full-strength solution, replaced every 5 d. Young plants were fixed onto foam boards with cotton and suspended in the nutrient solution. The hydroponic growth chamber maintained a constant temperature of 25 ± 1 °C throughout both day and night cycles, with controlled relative humidity at 60 ± 5% and a fixed 14 h daily photoperiod. Following 15 d of pre-cultivation in full-strength solution for stabilization, morphologically uniform apple young plants were allocated to five treatment groups.
Five treatments were applied: Control (CK, dH2O), applying foliar treatments of 200 mg L−1 ZnSO4 (S200) and 200, 500, and 1000 mg L−1 ZnO NPs (N200, N500, N1000). Each treatment included three replicates, with 20 plants in each replicate. A total of 300 plants were used for the five treatments. ZnO NPs were freshly prepared before each application and sprayed every 3–4 d, with 100 mL applied to the leaves each time. Spraying continued until leaf surfaces were saturated and droplets began to drip. The treatment period lasted 47 d. After treatment, young plants were harvested between 10:00 and 11:00 a.m. to minimize diurnal variation.
2.2. Growth Index Determination
Plant height was determined as the aboveground length using a measuring tape (0.1 cm precision), while stem diameter was measured with a digital caliper (0.01 mm accuracy). For dry weight determination, plants were sequentially subjected to ultrasonic cleaning (three cycles), triple rinsing with deionized water, and blot-drying with filter paper. The samples were then separated into root, stem and leaf components, placed in kraft paper bags, and oven-dried (105 °C for 30 min for enzyme deactivation followed by 65 °C drying to constant weight). The dry mass of each component was measured using an analytical balance (0.001 g precision), with total dry weight calculated as the sum of all components. Finally, the dried samples were homogenized using a ball mill, sieved, and stored in light-proof ziplock bags at low temperature for subsequent mineral element analysis.
2.3. Relative Growth Rate (RGR)
RGR was calculated using the formula:
where DW1 and DW2 are the total dry weights before and after treatment, and t2 − t1 is the treatment duration [32].
2.4. Root Architecture Analysis
The roots were ultrasonically cleaned, rinsed with deionized water, and evenly spread before being scanned at 600 dpi resolution using an Epson Perfection V850 Pro scanner. The acquired images were then analyzed with WinRHIZO Pro software (Version 2019a, Regent Instruments Inc., Québec City, QC, Canada) to quantify multiple root parameters including total root length, average diameter, projected surface area, and root tip number.
2.5. Biochemical Assays (MDA, H2O2, CAT, SOD, POD)
Samples (n = 6 biological replicates, each comprising 5 pooled fully expanded leaves from the 4th–5th leaves) were immediately flash-frozen in liquid nitrogen and stored at −80 °C until analysis. Prior to assays, frozen tissues were homogenized into fine powder using a pre-cooled high-throughput grinder (SCIENTZ-24L Cryogenic High-throughput Tissue Grinder, Ningbo Scientz Biotechnology Co., Ltd., Zhejiang, China) (SCIENTZ-48, 45 Hz, 1 min). For each sample, 0.1 g of powdered tissue was weighed for subsequent kit-based assays.
The contents of malondialdehyde (MDA), hydrogen peroxide (H2O2), catalase (CAT), superoxide dismutase (SOD), and peroxidase (POD) were measured using commercial kits (Beijing Solarbio Science & Technology Co., Ltd., Beijing, China) according to the manufacturer’s instructions.
2.6. Total Nitrogen (N)
Samples (n = 3 biological replicates) were measured as 0.2 g of dried plant material using a 0.0001 g precision balance for analysis. Measured by the Kjeldahl method (Automatic Kjeldahl Nitrogen Analyzer (K1160), Hanon Advanced Technology Group Co., Ltd., Shandong, China), involving sulfuric acid digestion, followed by distillation and titration [32].
2.7. Phosphorus, Potassium, Calcium, Magnesium, and Trace Elements
Samples (n = 3 biological replicates) were measured as 0.2 g of dried plant material using a 0.0001 g precision balance for analysis. Samples were digested with a 2:1 HNO3–HClO4 mixture. Phosphorus was determined by UV spectrophotometry (UV-1900 Double-Beam UV-Vis Spectrophotometer, Shanghai Aoxi Scientific Instrument Co., Ltd., Shanghai, China), potassium by flame spectrophotometry (FP6400A Flame Photometer, Shanghai Aopu Analytical Instrument Co., Ltd., Shanghai, China), and calcium, magnesium, and trace elements by atomic absorption spectrometry (AA-6880F Fatomic-absorption spectrophotometer, Shimadzu Corporation, Japan) [32].
2.8. Element Absorption Flux (Jupt)
Total element absorption was calculated as the sum of element fluxes from roots, stems, and leaves:
where RGR is relative growth rate, DW is tissue dry weight, and C is element concentration, with r for root, s for stem, and l for leaf [32].
TNr = RGR × DWr × Cr
TNs = RGR × DWs × Cs
TNl = RGR × DWl × Cl
Jupt = TNr + TNs + TNl
2.9. Element Accumulation and Transfer
Root accumulation was defined as the total element absorbed per unit root mass per unit time. Transfer was defined as the amount moved to shoots (leaves + stems) per unit root mass per unit time. The transfer/accumulation ratio was calculated as:
where M is total element content, DW is root dry weight, and t is treatment time [32].
2.10. Statistical Analysis
Data were analyzed using SPSS 26.0 2019 software (IBM Corporation, Armonk, NY, USA), and figures were created with OriginPro 2021 software (QriginLab Corporation, Northampton, MA, USA) [33]. One-way ANOVA was used to evaluate treatment effects, with Duncan’s test (p < 0.05) for multiple comparisons. Results are reported as mean ± standard error (SE).
3. Results
3.1. Plant Growth and Morphological Parameters
Foliar application of ZnO NPs significantly influenced the growth and development of young crabapple plants in a concentration-dependent manner. The N200 treatment resulted in significant increases in plant height, total dry weight, and RGR compared to CK, with no significant change observed in basal diameter (Figure 1A–E). Specifically, plants treated with N200 exhibited the most pronounced improvements: plant height increased by 22.64% (p = 0.011), total dry weight by 49.36% (p = 0.005), and RGR by 28.92% (p = 0.021) relative to CK (Figure 1B,D,E). In contrast, no significant differences were found between the N500, N1000, and CK in terms of plant height, basal diameter, total dry weight, and RGR. Furthermore, N200 demonstrated superior growth-promoting effects compared to ZnSO4 at equivalent Zn concentrations (Figure 1A–E).
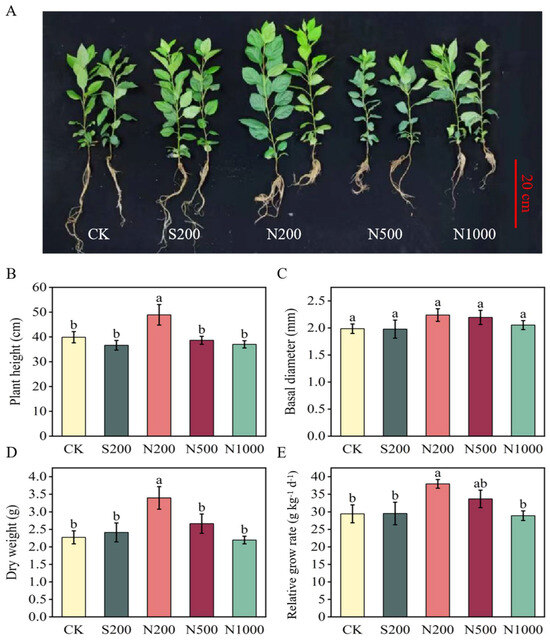
Figure 1.
Effects of foliar spraying with ZnSO4 and ZnO NPs on apple young plant growth. (A) Morphology, (B) plant height, (C) stem diameter, (D) dry weight, and (E) relative growth rate were determined after 47 d of treatment. CK: dH2O; S200: ZnSO4 at 200 mg L−1; N200: ZnO NPs at 200 mg L−1; N500: ZnO NPs at 500 mg L−1; N1000: ZnO NPs at 1000 mg L−1. Data are means ± SE (n = 3). Different letters indicate significant differences (p < 0.05, Duncan’s test).
Root architecture was also markedly improved by N200 treatment. Total root length increased by 116.07% (p = 0.003), average root diameter by 35.71% (p = 0.011), and root tip number by 152.90% (p = 0.001), all significantly higher than CK. However, there was no significant difference in root surface area between N200 and CK (Figure 2). Conversely, N1000 led to a 14.00% decrease in root tip number, but again, this was not statistically significant (Figure 2). No significant differences were observed between S200, N500, and CK in any of the root parameters (Figure 2).
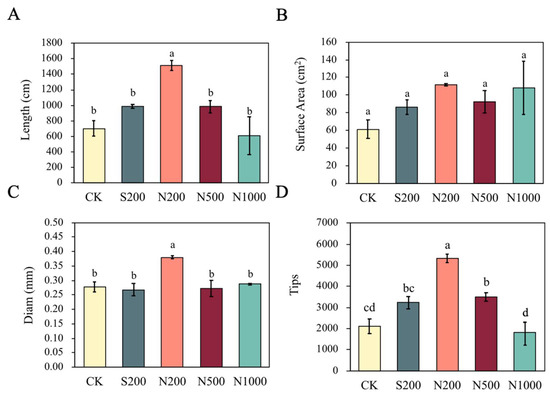
Figure 2.
Effects of foliar spraying with ZnSO4 and ZnO NPs on root architecture of apple young plants. (A) Total root length, (B) total root surface area, (C) average root diameter, and (D) root tip number were measured after 47 d of treatment. CK: dH2O; S200: ZnSO4 at 200 mg L−1; N200: ZnO NPs at 200 mg L−1; N500: ZnO NPs at 500 mg L−1; N1000: ZnO NPs at 1000 mg L−1. Data are means ± SE (n = 3). Different letters indicate significant differences (p < 0.05, Duncan’s test).
3.2. Oxidative Stress Indicators and Antioxidant Enzyme Activities
To evaluate potential stress responses induced by ZnO NPs foliar application, levels of H2O2 and MDA, along with antioxidant enzyme activities, were measured. N500 and N1000 treatments significantly elevated H2O2 levels by 38.46% and 32.02%, respectively, indicating oxidative stress (Figure 3A). Similarly, MDA content increased by 44.47% and 54.43% in N500 and N1000 treatments, respectively, indicating increased lipid peroxidation (Figure 3B). The S200 treatment also increased MDA by 35.02%, but no significant oxidative stress indicators were observed in the low-concentration N200 group (Figure 3A,B). No significant differences in SOD activity were observed between N500, N1000, and CK, while peroxidase POD and CAT activities increased, particularly under N500 (Figure 3C–E).
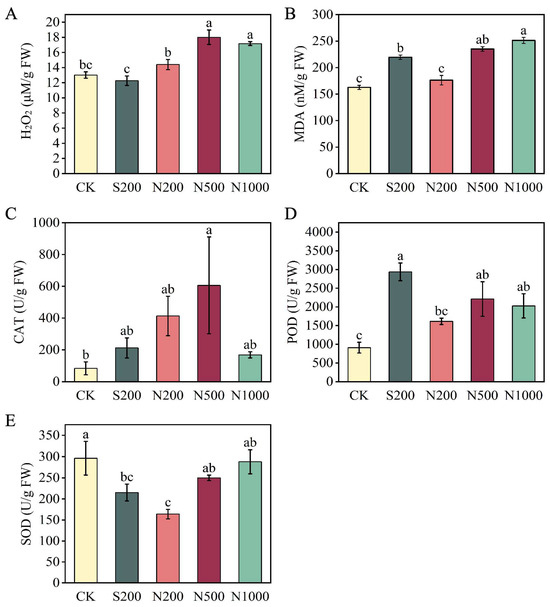
Figure 3.
Effects of foliar spraying with ZnSO4 and ZnO NPs on the antioxidant system. (A) hydrogen peroxide (H2O2) content, (B) Malondialdehyde (MDA) content, (C) catalase (CAT) activity, (D) peroxidase (POD) activity, and (E) superoxide dismutase (SOD) activity in leaves were measured after 47 d of treatment. FW: fresh weight. CK: dH2O; S200: ZnSO4 at 200 mg L−1; N200: ZnO NPs at 200 mg L−1; N500: ZnO NPs at 500 mg L−1; N1000: ZnO NPs at 1000 mg L−1. Data are means ± SE (n = 6). Different letters indicate significant differences (p < 0.05, Duncan’s test).
3.3. Mineral Nutrient Concentrations
As shown in Table 1, foliar application of ZnSO4 and ZnO NPs had no significant effect on the concentrations of macronutrients—nitrogen (N), phosphorus (P), and potassium (K)—in roots, stems, or leaves. However, ZnO NPs treatments notably altered calcium (Ca), magnesium (Mg), and trace element concentrations (Table 1 and Table 2).

Table 1.
Effects of foliar application of ZnSO4 and ZnO NPs on concentrations of macronutrients nutrients in apple young plants. The parameters were determined at 47 d of treatment. Nutrient contents are expressed as mg·g−1 DW (DW: dry weight).
Table 2.
Effects of foliar application of ZnSO4 and ZnO NPs on concentrations of trace elements in apple young plants. The parameters were determined at 47 d of treatment. Nutrient contents are expressed as μg·g−1 DW (DW: dry weight).
N1000 significantly increased root Ca levels, reaching 6.88 mg g−1-a 64.59% rise compared to the control (p = 0.008). Mg concentrations in roots and stems increased in all Zn-treated groups. Notably, root Mg in N500 rose by 184.21% (p < 0.001), and stem Mg in N500 increased by 229.51%, with the highest leaf Mg observed in N500 (2.26 mg·g−1) (Table 1).
Trace element analysis revealed that root iron (Fe) concentrations increased significantly in all treatments, with the N200 group showing the highest Fe level (197.71 μg·g−1), a 136.75% increase compared to CK (p = 0.031). Conversely, Fe concentrations in stems were significantly reduced in all treatments (p < 0.05). Manganese (Mn) concentrations remained largely unaffected across treatments. N200 significantly decreased copper (Cu) concentrations in stems (−34.68%, p = 0.017) and leaves (−22.37%, p = 0.016) relative to the control. Zn accumulation was markedly enhanced in the N200 group, with root Zn reaching 255.47 μg·g−1 (a 53.39% increase), whereas the S200 treatment resulted in lower root Zn levels than the control. In stems and leaves, Zn concentrations were significantly elevated in N500 and N1000 treatments (Table 2).
Overall, foliar application of ZnO NPs did not affect macronutrient (N, P, K) levels but markedly altered Ca, Mg, and trace element dynamics in young crabapple plants. Moderate doses (N200–N500) enhanced Mg, Fe, and Zn accumulation, while excessive application (N1000) increased root Ca but also reduced Cu in stems and leaves, indicating dose-dependent effects on nutrient uptake and distribution.
3.4. Nutrient Uptake and Allocation Patterns
As shown in Figure 4, foliar application of low concentrations of Zn—especially ZnO NPs—significantly enhanced the nutrient uptake flux of N, P, K, Ca, Mg, Fe, Mn, and Cu in apple young plants, while Zn uptake increased progressively with rising Zn concentration (Figure 4). N200 exhibited the highest uptake fluxes across most elements, with significant increases (p < 0.05) in K, Ca, and Fe compared to the control (Figure 4C,D,F). Specifically, N200 showed increases of 49.60%, 77.80%, 84.92%, 112.12%, 185.15%, 34.61%, and 41.67% in N, P, K, Ca, Fe, Mn, and Cu, respectively (Figure 4A–D,F–H). In contrast, N1000 reduced uptake fluxes of these elements, with N and Cu decreasing by 30.31% and 40.08%, respectively, though not significantly (Figure 4A,H).
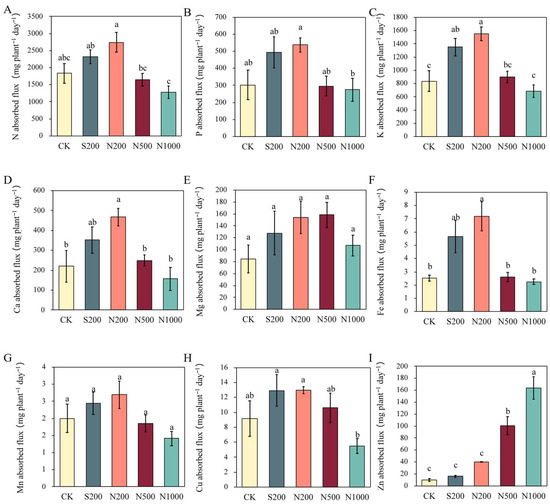
Figure 4.
Elemental absorption fluxes in apple young plants after 47 d of foliar spraying with ZnSO4 and ZnO NPs. (A) Nitrogen (N), (B) Phosphorus (P), (C) Potassium (K), (D) Calcium (Ca), (E) Magnesium (Mg), (F) Iron (Fe), (G) Manganese (Mn), (H) Copper (Cu), (I) Zinc (Zn). CK: dH2O; S200: ZnSO4 at 200 mg L−1; N200: ZnO NPs at 200 mg L−1; N500: ZnO NPs at 500 mg L−1; N1000: ZnO NPs at 1000 mg L−1. Data are means ± SE (n = 3). Different letters indicate significant differences (p < 0.05, Duncan’s test).
Further analysis of nutrient distribution (Figure 5A–I) revealed that S200 resulted in the highest N distribution in leaves (5.93 mg·g−1 DW·day−1) and stem transfer (0.79 mg g−1 DW day−1), whereas N200 showed the greatest N accumulation in roots (0.73 mg g−1 DW day−1, p = 0.007) (Figure 5A). N200 also had the highest K accumulation in roots (0.28 mg g−1 DW day−1), a 189.05% increase over the control, while N1000 showed the lowest stem K transfer (0.20 mg g−1 DW day−1), a significant 52.55% reduction (p = 0.013) (Figure 5C). Mg dynamics were most pronounced in the N500, which exhibited the highest leaf distribution, root accumulation (up by 739.51%), and stem transfer (up by 251.40%) relative to the control (Figure 5E). For Fe, N200 recorded the highest root accumulation (6.02 μg g−1 DW day−1), while N1000 had the lowest stem transfer (−0.27 μg g−1 DW day−1). S200 achieved the highest Fe leaf distribution (12.46 μg g−1 DW day−1), a 75.11% increase (p = 0.026) (Figure 5F). Cu accumulation peaked in N500 (10.02 μg g−1 DW day−1, +55.29%, p = 0.042) (H), while root Zn accumulation was highest in N200 (10.46 μg g−1 DW day−1, +149.92%, p = 0.004), with Zn leaf distribution increasing with Zn concentration (I). No significant differences were observed in the distribution of P, Ca across treatments (Figure 5B,D).
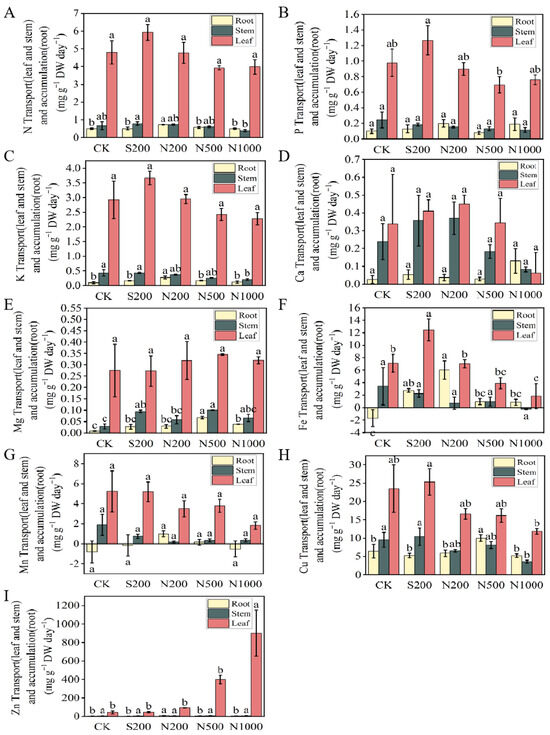
Figure 5.
Element distribution in apple young plants after 47 d of foliar application of ZnSO4 and ZnO NPs. (A) Nitrogen (N), (B) Phosphorus (P), (C) Potassium (K), (D) Calcium (Ca), (E) Magnesium (Mg), (F) Iron (Fe), (G) Manganese (Mn), (H) Copper (Cu), (I) Zinc (Zn). CK: dH2O; S200: ZnSO4 at 200 mg L−1; N200: ZnO NPs at 200 mg L−1; N500: ZnO NPs at 500 mg L−1; N1000: ZnO NPs at 1000 mg L−1. Dw: dry weight. Data are means ± SE (n = 3). Different letters indicate significant differences (p < 0.05, Duncan’s test).
In summary, the N200 treatment improved nutrient uptake and root accumulation, particularly for K, Fe, and Zn, while ZnSO4 favored nutrient distribution to aerial parts. In contrast, the N1000 treatment suppressed overall nutrient uptake but promoted Ca accumulation in roots.
4. Discussion
4.1. Concentration-Dependent Effects of ZnO NPs on Plant Growth Promotion
Our results demonstrate that foliar application of ZnO NPs elicits a concentration-dependent effect on young crabapple plants, with N200 treatment significantly promoting shoot and root development, while higher concentrations (N500, N1000) do not induce statistically significant inhibition compared to the control (Figure 1). This pattern diverges from reports in annual crops like cucumber [34], Spinach [35], maize [36], bean [37], and Alfalfa [38], where higher concentrations often suppress growth, but aligns with the greater physiological resilience of woody fruit trees. The lack of growth inhibition by N500 and N1000 in young crabapple plants in contrast with herbaceous species, highlighting fruit tree-specific tolerance mechanisms. Woody plants often prioritize root structural integrity over rapid growth under stress [39], and our observation of stable root parameters (Figure 2) in high-concentration groups supports this: preserved root function likely maintained nutrient supply, preventing growth declines. In our dataset, N200 treatment markedly improved root architecture, with total root length, average root diameter, and root tip number all significantly higher than CK, while root surface area showed only a non-significant increasing trend (Figure 2). This outcome is most likely attributable to the substantial biological variability inherent in woody perennial plants, which generally exhibit larger individual differences in root system morphology compared with annual herbaceous species. As a result, a higher number of biological replicates may be required to detect statistically significant differences in root surface area.
The growth promotion by N200 is tightly linked to root architectural improvements (Figure 2A,D), which likely expanded the absorptive surface area (Figure 2B, no significant difference in root surface area) and enhanced nutrient uptake capacity (Table 1 and Table 2). This aligns with findings in wheat, where ZnO NPs-induced root development correlated with increased biomass via improved nutrient acquisition [40]. Notably, the negligible response to ZnSO4 (S200) despite equivalent Zn input suggests that nano-sized Zn has superior foliar absorption efficiency. Ionic Zn may form insoluble complexes [41] on leaf surfaces or be excluded by membrane transporters [42], limiting translocation, whereas ZnO NPs can penetrate stomatal pores or cuticular layers [43].
4.2. Antioxidant Defense Mechanisms Mitigating ZnO NP-Induced Oxidative Stress
Our data reveal a nuanced relationship between ZnO NPs, oxidative stress, and growth in young crabapple plants: relatively high concentrations (N500 and N1000) elevated H2O2 and MDA levels (Figure 3A,B), indicating oxidative damage, yet plant growth was not inhibited, likely due to strong antioxidant compensation. This contrasts with annuals, where similar ROS levels correlate with biomass reduction [44], underscoring fruit trees’ superior redox regulation. Consistent with this notion, SOD activity showed no significant differences among N500, N1000, and CK, whereas POD and CAT activities were markedly enhanced, particularly under N500 (Figure 3C–E). This suggests that plants relied on POD- and CAT-mediated detoxification pathways to efficiently decompose excess hydrogen peroxide and mitigate oxidative stress, thereby preventing ROS accumulation from reaching phytotoxic thresholds that would impair growth. Notably, N200 avoided oxidative stress entirely, suggesting a “hormetic” threshold: low doses enhance growth without triggering stress responses, while high doses induce manageable stress via antioxidant upregulation.
4.3. Altered Mineral Nutrient Dynamics Under Foliar-Applied ZnO NPs
Foliar ZnO NPs selectively modulated nutrient concentrations in young crabapple plants, with minimal effects on macronutrients (N, P, K) but significant impacts on Ca, Mg, and trace elements (Table 1 and Table 2)—patterns reflecting both direct ZnO NPs effects and adaptive plant responses. The increase in root Ca under N1000 (Table 1) likely strengthened cell walls, reducing membrane permeability to toxic ions—a common tolerance mechanism in fruit trees [45]. Similarly, elevated Mg in N500 (Table 1) would support chlorophyll synthesis and photosynthesis, offsetting potential growth constraints. This aligns with another apple tree studies, where Mg accumulation under metal stress preserved photosynthetic efficiency [46].
This distribution pattern likely reflects an adaptive nutrient management strategy in young crabapple plants. The marked Fe accumulation in roots under N200 (Figure 5F; Table 2) suggests that ZnO NPs enhanced Fe uptake from the solution, but the retained Fe was preferentially stored in root tissues rather than translocated to shoots. Moreover, N200 treatment may have altered Fe transport dynamics by modulating root membrane transporters or apoplastic binding sites, leading to localized Fe sequestration. Such retention likely secures Fe for essential root metabolic functions while minimizing the risk of Fe-induced oxidative stress in aerial tissues. This aligns with [47], who reported excess Fe in the root apoplast can be sequestered in the cell wall, thereby restricting Fe translocation and reducing oxidative stress in shoots.
Zn concentrations in stems and leaves increased linearly with ZnO NP dosage (N500> N200 > CK), while root and leaves Zn in N200 exceeded S200 despite equal Zn input (Table 2). This confirms foliar-applied NPs as the primary source, as Zn from the nutrient solution would not exhibit such concentration dependence. Cui et al., (2023) used single-particle ICP-MS with isotopic tracing to quantify ZnO NP uptake in rice, showing that Zn derived from foliar-applied ZnO NPs contributed a major share of leaf Zn and was translocated via xylem and phloem, supporting our conclusion [48].
4.4. Mechanisms of Nutrient Uptake and Allocation Responses to ZnO NP Treatments
Our findings highlight that ZnO NPs regulate nutrient uptake and allocation in a dose-dependent manner, with N200 optimizing uptake efficiency and high concentrations triggering adaptive redistribution. N200’s enhancement of uptake fluxes (Figure 4) directly links to root architectural improvements: more root tips (Figure 2) likely increased contact with soil nutrients, while ZnO NPs may upregulate metal transporter activity [49]. In contrast, N1000’s mild uptake suppression was offset by strategic allocation—for example, prioritizing Mg transport to stems/leaves to sustain photosynthesis (Figure 5E).
Notably, S200 favored nutrient distribution to aerial parts, while N200 enhanced root accumulation (Figure 5). This divergence reflects differing Zn forms: ionic Zn may be rapidly translocated to shoots, whereas ZnO NPs trigger signaling pathways that prioritize root nutrient storage—critical for perennial fruit trees, as woody plants tend to prioritize root structural integrity over rapid growth when under stress [39].
4.5. Integrative Adaptive Strategies of Young Crabapple Plants Responses to ZnO NPs
Taken together, our findings suggest that young crabapple plants respond to foliar-applied ZnO NPs through a combination of morphological, physiological, and biochemical adjustments. At optimal concentrations (N200), ZnO NPs stimulate root system development, thereby enhancing nutrient uptake efficiency and promoting growth. At higher concentrations (N500 and N1000), although oxidative stress is induced, plants activate antioxidant defenses (mainly POD and CAT) and adjust nutrient allocation (e.g., Ca accumulation for membrane stability, Mg transport for photosynthesis, and Fe sequestration in roots), which collectively mitigate stress damage and sustain growth. This integrative response reflects the typical strategy of perennial woody plants, which prioritize root structural integrity, redox homeostasis, and nutrient redistribution to adapt to environmental challenges [39,50,51].
5. Conclusions
Foliar-applied ZnO NPs exhibited clear concentration-dependent effects on young crabapple plants. The N200 treatment significantly promoted growth by improving root architecture and enhancing the uptake of K, Fe, and Zn without inducing oxidative stress. Higher concentrations (N500 and N1000) triggered oxidative stress but did not suppress growth, likely due to strong antioxidant responses and adaptive nutrient sequestration. ZnSO4 was comparatively less effective, resulting in lower root Zn accumulation. Overall, N200 represents an optimal foliar strategy to improve nutrient acquisition in apple young plants, providing a practical recommendation for hydroponic apple propagation and offering promising implications for sustainable fruit tree cultivation.
Author Contributions
Q.Z., L.L. (Lei Li) and Q.L. designed the experiments; Q.Z., M.Q. (Meimei Qin), Q.L. and S.L. performed experiments; M.Q. (Meimei Qin), Q.L., M.Q. (Mengyao Qin) and D.Z. analyzed data; D.Z., L.L. (Lizhi Liu) and Q.Z. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Key Research and Development Project of Shanxi Province (202302140601010), Shanxi Provincial Basic Research Program (202303021222053), the Science and Technology Innovation Project for Higher Education Institutions in Shanxi Province (2022L106, 2023L035), and the Earmarked fund for the China Agricultural Research System (CARS-27).
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Baligar, V.C.; Fageria, N.K.; He, Z.L. Nutrient use efficiency in plants. Commun. Soil Sci. Plant Anal. 2001, 32, 921–950. [Google Scholar] [CrossRef]
- Zhai, J.; Xian, X.; Zhang, Z.; Wang, Y. Nano-zinc oxide can enhance the tolerance of apple rootstock M9-T337 seedlings to saline alkali stress by initiating a variety of physiological and biochemical pathways. Plants 2025, 14, 233. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, A.; Sharma, I.; Singh, R.; Rana, M. Nanotechnology for sustainability of agriculture and environment: Green synthesis and application of nanoparticles—A review. Res. Crops 2024, 25, 578. [Google Scholar] [CrossRef]
- Kourgialas, N.; Karatzas, G.; Koubouris, G. A GIS policy approach for assessing the effect of fertilizers on the quality of drinking and irrigation water and wellhead protection zones (Crete, Greece). J. Environ. Manag. 2017, 189, 150–159. [Google Scholar] [CrossRef]
- Morales-Díaz, A.; Ortega-Ortíz, H.; Juárez-Maldonado, A.; Cadenas-Pliego, G.; González-Morales, S.; Benavides-Mendoza, A. Application of nanoelements in plant nutrition and its impact in ecosystems. Adv. Nat. Sci. Nanosci. Nanotechnol. 2017, 8, 013001. [Google Scholar] [CrossRef]
- Raliya, R.; Saharan, V.; Dimkpa, C.; Biswas, P. Nanofertilizer for precision and sustainable agriculture: Current state and future perspectives. J. Agric. Food Chem. 2018, 66, 6487–6503. [Google Scholar] [CrossRef]
- Davarpanah, S.; Tehranifar, A.; Davarynejad, G.; Abadía, J.; Khorasani, R. Effects of foliar applications of zinc and boron nano-fertilizers on pomegranate (Punica granatum cv. Ardestani) fruit yield and quality. Sci. Hortic. 2016, 210, 57–64. [Google Scholar] [CrossRef]
- Kah, M.; Kookana, R.; Gogos, A.; Bucheli, T. A critical evaluation of nanopesticides and nanofertilizers against their conventional analogues. Nat. Nanotechnol. 2018, 13, 677–684. [Google Scholar] [CrossRef]
- Eichert, T.; Kurtz, A.; Steiner, U.; Goldbach, H.E. Size exclusion limits and lateral heterogeneity of the stomatal foliar uptake pathway for aqueous solutes and water-suspended nanoparticles. Physiol. Plant 2008, 134, 151–160. [Google Scholar] [CrossRef]
- Das, S.; Green, A. Zinc in crops and human health. In Biofortification Ofood Crops; Singh, U., Praharaj, C., Singh, S., Singh, N., Eds.; Springer: New Delhi, India, 2016; pp. 31–40. [Google Scholar]
- Ranjbar, S.; Rahemi, M.; Ramezanian, A. Comparison of nano-calcium and calcium chloride spray on postharvest quality and cell wall enzymes activity in apple cv. Red Delicious. Sci. Hortic. 2018, 240, 57–64. [Google Scholar] [CrossRef]
- Elsheery, N.; Helaly, M.; El-Hoseiny, H.; Alam-Eldein, S. Zinc oxide and silicone nanoparticles to improve the resistance mechanism and annual productivity of salt-stressed mango trees. Agronomy 2020, 10, 558. [Google Scholar] [CrossRef]
- Genaidy, E.; Abd-Alhamid, N.; Hassan, H.; Hassan, A.; Hagagg, L. Effect of foliar application of boron trioxide and zinc oxide nanoparticles on leaves chemical composition, yield and fruit quality of Olea europaea L. cv. Picual. Bull. Natl. Res. Cent. 2020, 44, 106. [Google Scholar] [CrossRef]
- García-López, J.; Niño-Medina, G.; Olivares-Sáenz, E.; Lira-Saldivar, R.; Barriga-Castro, E.; Vázquez-Alvarado, R.; Rodríguez-Salinas, P.; Zavala-García, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compounds in habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- López-Vargas, E.; Ortega-Ortíz, H.; Cadenas-Pliego, G.; De Alba Romenus, K.; Cabrera de la Fuente, M.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Foliar application of copper nanoparticles increases the fruit quality and the content of bioactive compounds in tomatoes. Appl. Sci. 2018, 8, 1020. [Google Scholar] [CrossRef]
- Singh, G.; Kalia, A. Nano-enabled technological interventions for sustainable production, protection, and storage of fruit crops. In Nanoscience for Sustainable Agriculture; Pudake, R., Chauhan, N., Kole, C., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 299–322. [Google Scholar]
- Verma, R.; Chettri, N.; Kumar, R.; Roshan, R.; Kumari, P.; Sonkar, S.; Kumari, S.; Rongpi, A. Nanotechnology interventions in fruit production: Enhancing production and quality. Int. J. Plant Soil Sci. 2024, 36, 575–586. [Google Scholar] [CrossRef]
- Manzoor, M.; Korolev, K.; Ahmad, R. Zinc oxide nanoparticles: Abiotic stress tolerance in fruit crops focusing on sustainable production. Phyton-Int. J. Exp. Bot. 2025, 94, 1401–1418. [Google Scholar] [CrossRef]
- Anwar, T.; Safdar, A.; Qureshi, H.; Siddiqi, E.H.; Ullah, N.; Naseem, M.T.; Soufan, W. Synergistic effects of Vachellia nilotica-derived zinc oxide nanoparticles and melatonin on drought tolerance in Fragaria × ananassa. BMC Plant Biol. 2025, 25, 82. [Google Scholar] [CrossRef]
- Sodhi, G.; Wijesekara, T.; Kumawat, K.; Adhikari, P.; Joshi, K.; Singh, S.; Farda, B.; Djebaili, R.; Sabbi, E.; Ramila, F.; et al. Nanomaterials–plants–microbes interaction: Plant growth promotion and stress mitigation. Front. Microbiol. 2025, 15, 1516794. [Google Scholar] [CrossRef]
- Adil, M.; Bashir, S.; Bashir, S.; Aslam, Z.; Ahmad, N.; Younas, T.; Asghar, R.M.A.; Alkahtani, J.; Dwiningsih, Y.; Elshikh, M.S. Zinc oxide nanoparticles improved chlorophyll contents, physical parameters, and wheat yield under salt stress. Front. Plant Sci. 2022, 13, 932861. [Google Scholar] [CrossRef]
- Azarin, K.; Usatov, A.; Minkina, T.; Plotnikov, A.; Kasyanova, A.; Fedorenko, A.; Duplii, N.; Vechkanov, E.; Rajput, V.D.; Mandzhieva, S.; et al. Effects of ZnO nanoparticles and its bulk form on growth, antioxidant defense system and expression of oxidative stress related genes in Hordeum vulgare L. Chemosphere 2022, 287, 132167. [Google Scholar] [CrossRef]
- Ji, H.; Guo, Z.; Wang, G.; Wang, X.; Liu, H. Effect of ZnO and CuO nanoparticles on the growth, nutrient absorption, and potential health risk of the seasonal vegetable Medicago polymorpha L. Peer J. 2022, 10, e14038. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Wang, R.; Wang, R.; Ju, Q.; Wang, Y.; Xu, J. Comparative physiological and transcriptomic analyses reveal the toxic effects of ZnO nanoparticles on plant growth. Environ. Sci. Technol. 2019, 53, 4235–4244. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Garg, T.; Joshi, A.; Awasthi, A.; Kumar, V.; Kumar, A. Potential effects of metal oxide nanoparticles on leguminous plants: Practical implications and future perspectives. Sci. Hortic. 2024, 331, 113146. [Google Scholar] [CrossRef]
- Rani, S.; Kumari, N.; Sharma, V. Zinc oxide nanoparticles improve photosynthesis by modulating antioxidant system and psb a gene expression under arsenic stress in different cultivars of Vigna radiata. Bionanoscience 2025, 15, 242. [Google Scholar] [CrossRef]
- Sarkhosh, S.; Kahrizi, D.; Darvishi, E.; Tourang, M.; Haghighi-Mood, S.; Vahedi, P.; Ercisli, S. Effect of zinc oxide nanoparticles (ZnO-NPs) on seed germination characteristics in two brassicaceae family species: Camelina sativa and Brassica napus L. J. Nanomater. 2022, 2022, 1892759. [Google Scholar] [CrossRef]
- Srivastav, A.; Ganjewala, D.; Singhal, R.; Rajput, V.; Minkina, T.; Voloshina, M.; Srivastava, S.; Shrivastava, M. Effect of ZnO nanoparticles on growth and biochemical responses of wheat and maize. Plants 2021, 10, 2556. [Google Scholar] [CrossRef]
- Fu, C.; Li, M.; Zhang, Y.; Zhang, Y.; Yan, Y.; Wang, Y. Morphology, photosynthesis, and internal structure alterations in field apple leaves under hidden and acute zinc deficiency. Sci. Hortic. 2015, 193, 47–54. [Google Scholar] [CrossRef]
- Ananthakrishnan, S.; Sharma, J.C.; Sharma, N.; Ranjha, R.; Vishnu Shankar, S. Comparative efficiency of nano, chelated, and sulfate zinc foliar applications in high-density apple orchards. J. Plant Nutr. 2025, 48, 1646–1661. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, J.; Liu, G.; Yao, Q.; Wang, Z.; Liu, N.; He, Y.; Il, D.; Tusupovich, J.; Jiang, Z. ZnO NPs impair the viability and function of porcine granulosa cells through autophagy regulated by ROS production. Antioxidants 2024, 13, 1295. [Google Scholar] [CrossRef]
- Liang, B.; Gao, T.; Zhao, Q.; Ma, C.; Chen, Q.; Wei, Z.; Li, C.; Li, C.; Ma, F. Effects of exogenous dopamine on the uptake, transport, and resorption of apple ionome under moderate drought. Front. Plant Sci. 2018, 9, 755. [Google Scholar] [CrossRef]
- Zhou, H.; Li, J.; Su, Y.; Sun, Y.; Lu, S.; Li, R.; Ma, L.; Chen, C.; Peng, Z.; Zhang, L. Integrated efficacy evaluation of mist and drip irrigation on vegetative performance of young apple trees. Sci. Hortic. 2025, 346, 114182. [Google Scholar] [CrossRef]
- Moghaddasi, S.; Fotovat, A.; Khoshgoftarmanesh, A.; Karimzadeh, F.; Khazaei, H.; Khorassani, R. Bioavailability of coated and uncoated ZnO nanoparticles to cucumber in soil with or without organic matter. Ecotoxicol. Environ. Saf. 2017, 144, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Kumar, A. Impact of irrigation using water containing CuO and ZnO nanoparticles on spinach oleracea grown in soil media. Bull. Environ. Contam. Toxicol. 2016, 97, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, X.; Shi, Z.; Tong, R.; Adams, C.; Shi, X. Arbuscular mycorrhizae alleviate negative effects of zinc oxide nanoparticle and zinc accumulation in maize plants—A soil microcosm experiment. Chemosphere 2016, 147, 88–97. [Google Scholar] [CrossRef]
- Dimkpa, C.; Hansen, T.; Stewart, J.; McLean, J.; Britt, D.; Anderson, A. ZnO nanoparticles and root colonization by a beneficial pseudomonad influence essential metal responses in bean (Phaseolus vulgaris). Nanotoxicology 2015, 9, 271–278. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Plascencia-Villa, G.; Mukherjee, A.; Rico, C.M.; José-Yacamán, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Comparative phytotoxicity of ZnO NPs, bulk ZnO, and ionic zinc onto the alfalfa plants symbiotically associated with Sinorhizobium meliloti in soil. Sci. Total Environ. 2015, 515–516, 60–69. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Uscola, M.; Jacobs, D. The role of stored carbohydrates and nitrogen in the growth and stress tolerance of planted forest trees. New For. 2015, 46, 813–839. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; Zia ur Rehman, M.; Waris, A. Zinc and iron oxide nanoparticles improved the plant growth and reduced the oxidative stress and cadmium concentration in wheat. Chemosphere 2019, 214, 269–277. [Google Scholar] [CrossRef]
- Gupta, N.; Ram, H.; Kumar, B. Mechanism of Zinc absorption in plants: Uptake, transport, translocation and accumulation. Rev. Environ. Sci. Bio/Technol. 2016, 15, 89–109. [Google Scholar] [CrossRef]
- Krishna, T.; Maharajan, T.; Ceasar, S. The role of membrane transporters in the biofortification of zinc and iron in plants. Biol. Trace Elem. Res. 2023, 201, 464–478. [Google Scholar] [CrossRef]
- Zhu, J.; Li, J.; Shen, Y.; Liu, S.; Zeng, N.; Zhan, X.; White, J.; Gardea-Torresdey, J.; Xing, B. Mechanism of zinc oxide nanoparticle entry into wheat seedling leaves. Environ. Sci. Nano 2020, 7, 3901–3913. [Google Scholar] [CrossRef]
- Rane, J.; Singh, A.; Tiwari, M.; Prasad, P.; Jagadish, S. Effective use of water in crop plants in dryland agriculture: Implications of reactive oxygen species and antioxidative system. Front. Plant Sci. 2022, 12, 778270. [Google Scholar] [CrossRef] [PubMed]
- Larocca, G.; Baldi, E.; Toselli, M. Understanding the role of calcium in kiwifruit: Ion transport, signaling, and fruit quality. Horticulturae 2025, 11, 335. [Google Scholar] [CrossRef]
- Lyu, M.; Liu, J.; Xu, X.; Liu, C.; Qin, H.; Zhang, X.; Tian, G.; Jiang, H.; Jiang, Y.; Zhu, Z.; et al. Magnesium alleviates aluminum-induced growth inhibition by enhancing antioxidant enzyme activity and carbon–nitrogen metabolism in apple seedlings. Ecotoxicol. Environ. Saf. 2023, 249, 114421. [Google Scholar] [CrossRef]
- Hu, H.; He, Y.; Gao, Y.; Chen, S.; Gu, T.; Peng, J. NnMTP10 from Nelumbo nucifera acts as a transporter mediating manganese and iron efflux. Plant Mol. Biol. 2025, 115, 26. [Google Scholar] [CrossRef]
- Cui, H.; Zhang, Z.; Wang, T.; Hong, J.; Lei, L.; Weia, S. Uptake and translocation of ZnO nanoparticles in rice tissues studied by single particle-ICP-MS. At. Spectrosc. 2023, 44, 343–353. [Google Scholar] [CrossRef]
- Skiba, E.; Michlewska, S.; Pietrzak, M.; Wolf, W. Additive interactions of nanoparticulate ZnO with copper, manganese and iron in Pisum sativum L., a hydroponic study. Sci. Rep. 2020, 10, 13574. [Google Scholar] [CrossRef]
- Li, S.; Lu, S.; Wang, J.; Chen, Z.; Zhang, Y.; Duan, J.; Liu, P.; Wang, X.; Guo, J. Responses of physiological, morphological and anatomical traits to abiotic stress in woody plants. Forests 2023, 14, 1784. [Google Scholar] [CrossRef]
- Rosell, J.; Marcati, C.; Olson, M.; Lagunes, X.; Vergilio, P.; Jiménez-Vera, C.; Campo, J. Inner bark vs sapwood is the main driver of nitrogen and phosphorus allocation in stems and roots across three tropical woody plant communities. New Phytol. 2023, 239, 1665–1678. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).