
Time–Course Analysis of the Onset and Progression of Cuticle Cracking in Fruits of Cherry Tomato Cultivar ‘Nene’


Abstract
1. Introduction
2. Materials and Methods
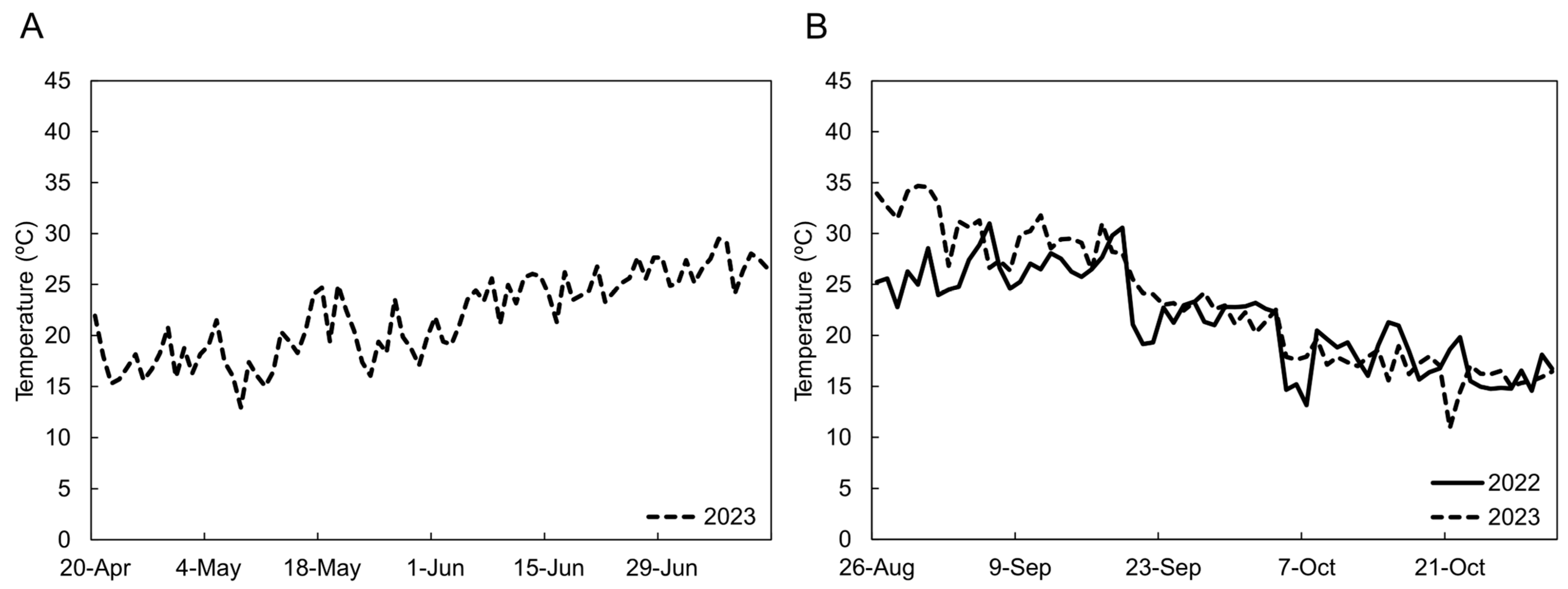
2.1. Plant Materials and Cultivation
2.2. Determination of Fruit Size and Onset of Cuticular Cracks
2.3. Measurement of CM Deposition and Epidermal Cell Density
2.4. Statistical Analyses
3. Results
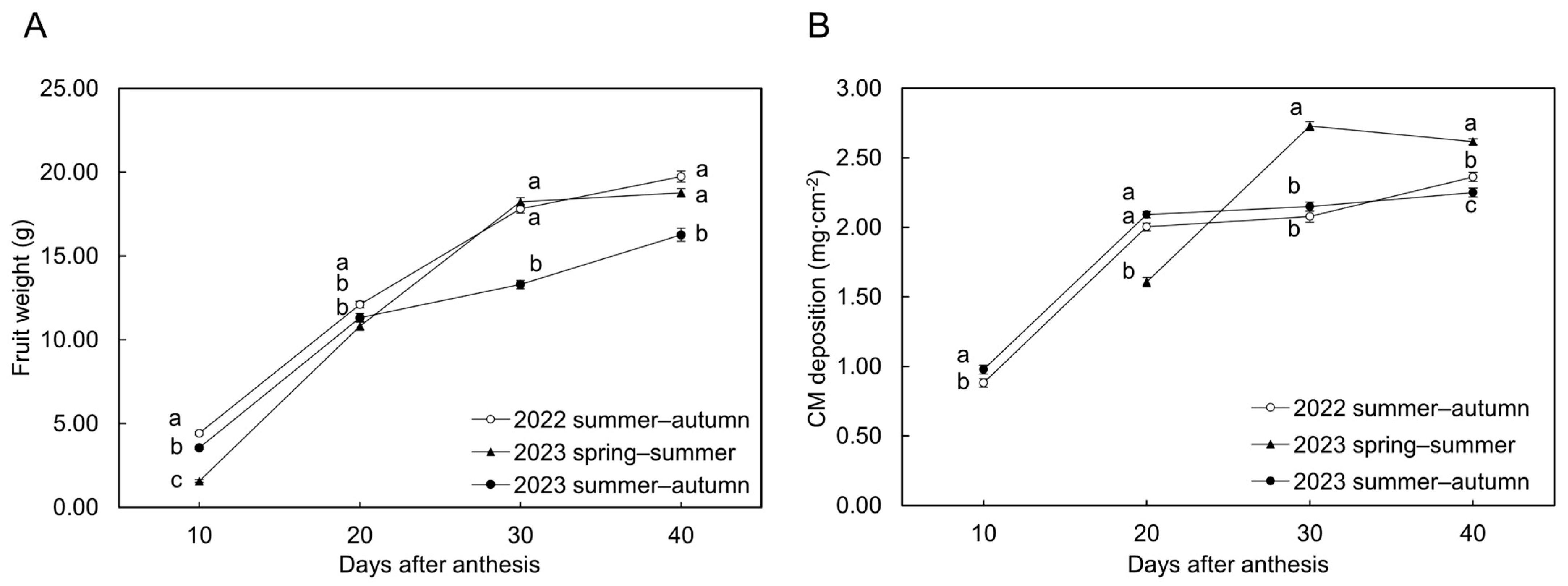
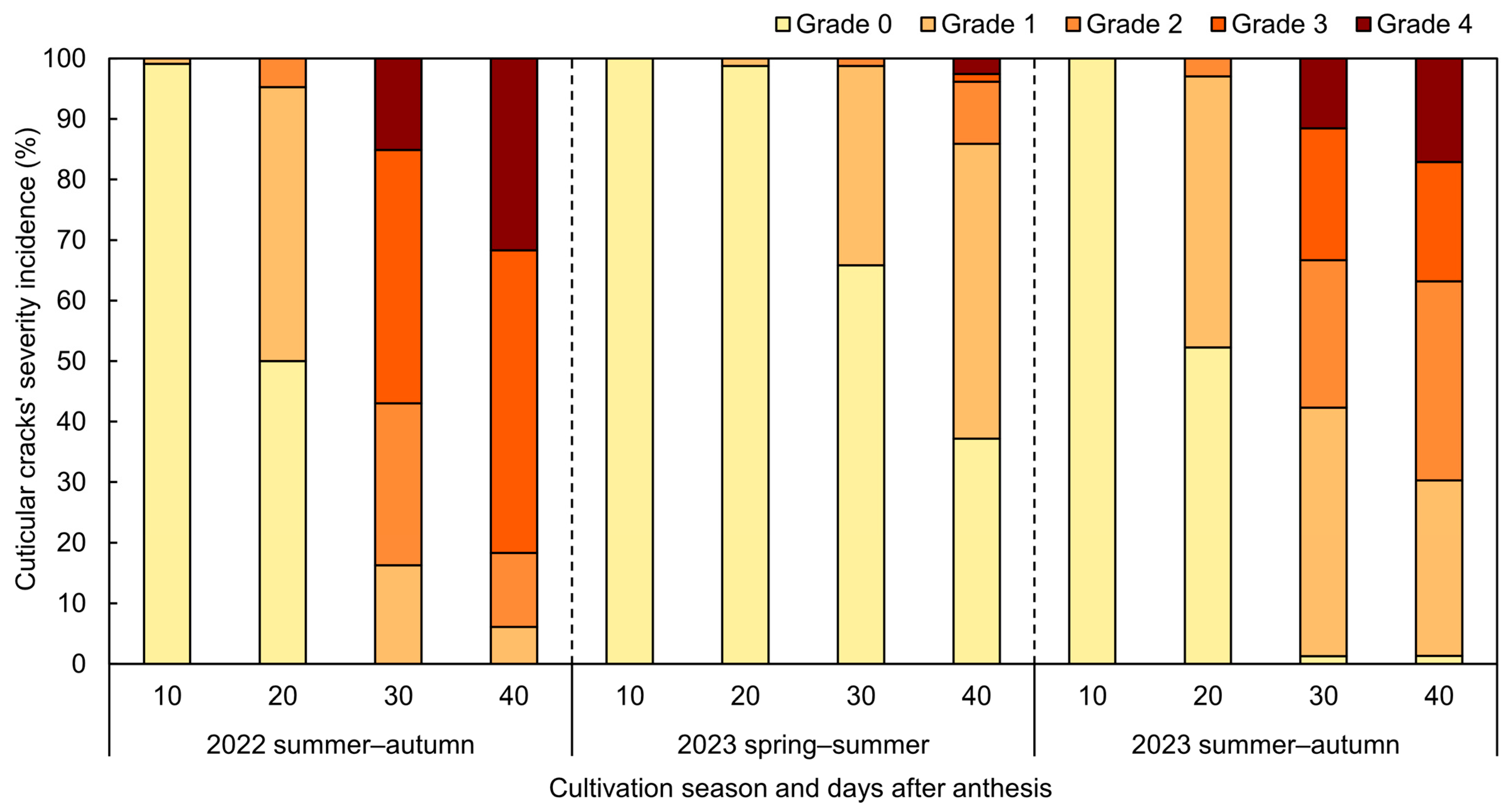
3.1. 2023 Spring–Summer Cultivation
3.2. 2022 and 2023 Summer–Autumn Cultivations
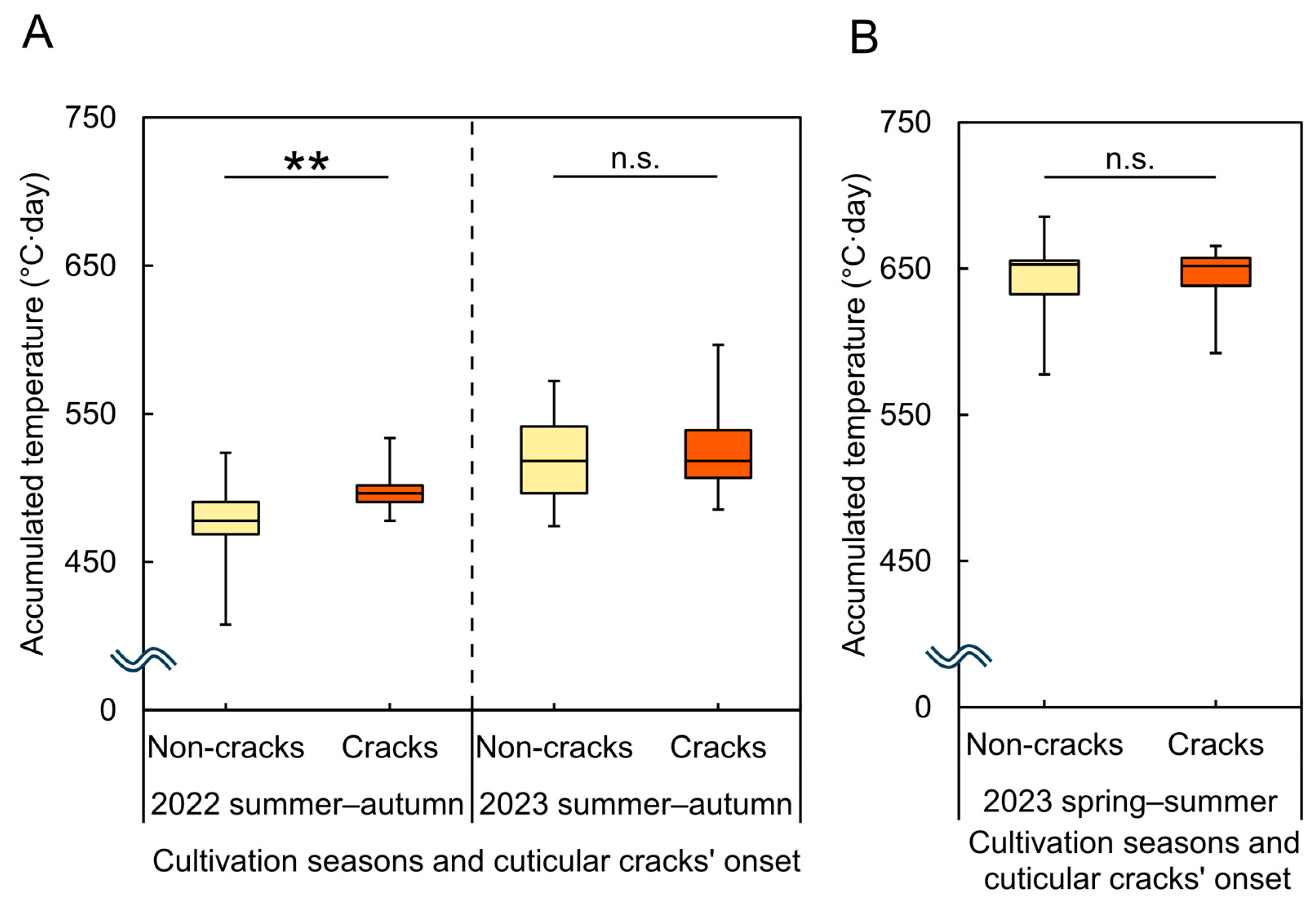
3.3. Analysis of the Relationship Between Cuticular Cracks’ Onset and Environmental Factors
4. Discussion
4.1. Onset of Cuticular Cracks
4.2. Progress of Cuticular Cracks Stages into Severe Net-like Forms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Holloway, P.J. Structure and histochemistry of plant cuticular membranes: An overview. In The Plant Cuticle; Cutler, D.F., Alvin, K.L., Price, C.E., Eds.; Academic Press: London, UK, 1982; pp. 1–32. [Google Scholar]
- Baker, E.A.; Bukovac, M.J.; Hunt, G.M. Composition of tomato fruit cuticle as related to fruit growth and development. In The Plant Cuticle; Cutler, D.F., Alvin, K.L., Price, C.E., Eds.; Academic Press: London, UK, 1982; pp. 33–44. [Google Scholar]
- Jeffree, C.E. The fine structure of the plant cuticle. Annu. Plant Rev. 2006, 23, 11–125. [Google Scholar] [CrossRef]
- Domínguez, E.; Cuartero, J.; Heredia, A. An overview on plant cuticle biomechanics. Plant Sci. 2011, 181, 77–84. [Google Scholar] [CrossRef]
- Yeats, T.H.; Rose, J.K.C. The Formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef]
- Reynoud, N.; Petit, J.; Bres, C.; Lahaye, M.; Rothan, C.; Marion, D.; Bakan, B. The complex architecture of plant cuticles and its relation to multiple biological functions. Front. Plant Sci. 2021, 12, 782773. [Google Scholar] [CrossRef] [PubMed]
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Lara, I.; Heredia, A.; Domínguez, E. Shelf life potential and the fruit cuticle: The unexpected player. Front. Plant Sci. 2019, 10, 770. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Muñoz, R.; Heredia, A.; Domínguez, E. The role of cuticle in fruit shelf-life. Curr. Opin. Biotechnol. 2022, 78, 102802. [Google Scholar] [CrossRef]
- Reina-Pinto, J.J.; Yephremov, A. Surface lipids and plant defenses. Plant Physiol. Biochem. 2009, 47, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, T.; Kosma, D.K.; Matas, A.J.; Buda, G.J.; He, Y.; Yu, B.; Pravitasari, A.; Batteas, J.D.; Stark, R.E.; Jenks, M.A.; et al. Cutin deficiency in the tomato fruit cuticle consistently affects resistance to microbial infection and biomechanical properties, but not transpirational water loss. Plant J. 2009, 60, 363–377. [Google Scholar] [CrossRef]
- Serrano, M.; Coluccia, F.; Torres, M.; L’Haridon, F.; Métraux, J.P. The cuticle and plant defense to pathogens. Front. Plant Sci. 2014, 5, 274. [Google Scholar] [CrossRef] [PubMed]
- Knoche, M.; Lang, A. Ongoing growth challenges fruit skin integrity. CRC Crit. Rev. Plant Sci. 2017, 36, 190–215. [Google Scholar] [CrossRef]
- Winkler, A.; Athoo, T.; Knoche, M. Russeting of fruits: Etiology and management. Horticulturae 2022, 8, 231. [Google Scholar] [CrossRef]
- Dorais, M.; Demers, D.-A.; Papadopoulos, A.P.; Van Ieperen, W. Greenhouse tomato fruit cuticle cracking. Hortic. Rev. 2004, 30, 163–184. [Google Scholar] [CrossRef]
- Ehret, D.L.; Hill, B.D.; Raworth, D.A.; Estergaard, B. Artificial neural network modelling to predict cuticle cracking in greenhouse peppers and tomatoes. Comput. Electron. Agric. 2008, 61, 108–116. [Google Scholar] [CrossRef]
- Hayman, G. The hair-like cracking of last season. Grower 1987, 107, 3–5. [Google Scholar]
- Blanke, M.M. Comparative SEM-study of the surface morphology of two solanaceae fruit, tomato and aubergine. Gartenbauwissenschaft 1986, 51, 225–230. [Google Scholar]
- Young, P.A. Cuticle cracks in tomato fruit. Phytopathology 1947, 37, 143–145. [Google Scholar]
- Schilstra-van Veelen, I.M.; Bakker, J.C. Cracking of tomato fruits. In Annual Report 1985; Glasshouse Crops Research and Experiment Station: Naaldwijk, The Netherland, 1985; pp. 39–40. [Google Scholar]
- Emmons, C.L.W.; Scott, J.W. Ultrastructural and anatomical factors associated with resistance to cuticle cracking in tomato (Lycopersicon esculentum Mill.). Int. J. Plant Sci. 1998, 159, 14–22. [Google Scholar] [CrossRef]
- Demers, D.-A.; Dorais, M.; Papadopoulos, A.P. Yield and russeting of greenhouse tomato as influenced by leaf-to-fruit ratio and relative humidity. HortScience 2007, 42, 503–507. [Google Scholar] [CrossRef]
- Gonroudobou, G.B.M.; Nabeshima, T.; Nishizawa, T.; Watanabe, M. Boron deficiency enhances microcracking in tomato fruit during summer. Chiang Mai J. Sci. 2022, 49, 1040–1049. [Google Scholar] [CrossRef]
- Aloni, B.; Karni, L.; Rylski, I.; Cohen, Y.; Lee, Y.; Fuchs, M.; Moreshet, S.; Yao, C. Cuticular cracking in pepper fruit. I. effects of night temperature and humidity. J. Hortic. Sci. Biotechnol. 1998, 73, 743–749. [Google Scholar] [CrossRef]
- Liu, G.S.; Huang, H.; Grierson, D.; Gao, Y.; Ji, X.; Peng, Z.Z.; Li, H.L.; Niu, X.L.; Jia, W.; He, J.L.; et al. NAC transcription factor SlNOR-like1 plays a dual regulatory role in tomato fruit cuticle formation. J. Exp. Bot. 2024, 75, 1903–1918. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, S.; Yoshikawa, H.; Ito, K. Studies on fruit cracking in tomatoes. Bull. Hort. Res. Sta. Ser. C Morioka 1972, 7, 73–138, (In Japanese with English summary). [Google Scholar]
- Bakker, J.C. Russeting (cuticle cracking) in glasshouse tomatoes in relation to fruit growth. J. Hortic. Sci. 1988, 63, 459–463. [Google Scholar]
- Emmons, C.L.W.; Scott, J.W. Environmental and physiological effects on cuticle cracking in tomato. J. Am. Soc. Hortic. Sci. 1997, 122, 797–801. [Google Scholar] [CrossRef]
- Jobin-Lawler, F.; Simard, K.; Gosselin, A.; Papadopoulos, A.P.; Dorais, M. The Influence of solar radiation and boron-calcium fruit application on cuticle cracking of a winter tomato crop grown under supplemental lighting. Acta Hortic. 2002, 580, 235–239. [Google Scholar] [CrossRef]
- Huang, J.-S.; Snapp, S.S. The effect of boron, calcium, and surface moisture on shoulder check, a quality defect in fresh-market tomato. J. Am. Soc. Hortic. Sci. 2004, 129, 599–607. [Google Scholar] [CrossRef]
- Shimazu, M.; Motoki, S. Comparison of tomato distribution in Japan and several other countries. Acta Hortic. 2022, 1353, 247–254. [Google Scholar] [CrossRef]
- Hosoi, R.; Ikeda, K.; Nishizawa, T.; Nabeshima, T. Effect of parthenocarpic fruit development on the occurrence of severe microcracking in cherry tomato. Acta Hortic. 2024, 1404, 1079–1084. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Ehret, D.L.; Helmer, T.; Hall, J.W. Cuticle cracking in tomato fruit. J. Hortic. Sci. 1993, 68, 195–201. [Google Scholar] [CrossRef]
- Khanal, B.P.; Knoche, M. Mechanical properties of cuticles and their primary determinants. J. Exp. Bot. 2017, 68, 5351–5367. [Google Scholar] [CrossRef] [PubMed]
- Bargel, H.; Neinhuis, C. Tomato (Lycopersicon esculentum Mill.) fruit growth and ripening as related to the biomechanical properties of fruit skin and isolated cuticle. J. Exp. Bot. 2005, 56, 1049–1060. [Google Scholar] [CrossRef]
- López-Casado, G.; Matas, A.J.; Domínguez, E.; Cuartero, J.; Heredia, A. Biomechanics of isolated tomato (Solanum lycopersicum L.) fruit cuticles: The role of the cutin matrix and polysaccharides. J. Exp. Bot. 2007, 58, 3875–3883. [Google Scholar] [CrossRef]
- López-Casado, G.; Salamanca, A.; Heredia, A. Viscoelastic nature of isolated tomato (Solanum lycopersicum) fruit cuticles: A mathematical model. Physiol. Plant. 2010, 140, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Petracek, P.D.; Bukovac, M.J. Rheological properties of enzymatically isolated tomato fruit cuticle. Plant Physiol. 1995, 109, 675–679. [Google Scholar] [CrossRef] [PubMed]
- España, L.; Heredia-Guerrero, J.A.; Segado, P.; Benítez, J.J.; Heredia, A.; Domínguez, E. Biomechanical properties of the tomato (Solanum lycopersicum) fruit cuticle during development are modulated by changes in the relative amounts of its components. New Phytol. 2014, 202, 790–802. [Google Scholar] [CrossRef]
- Marga, F.; Pesacreta, T.C.; Hasenstein, K.H. Biochemical analysis of elastic and rigid cuticles of Cirsium horridulum. Planta 2001, 213, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Bargel, H.; Neinhuis, C. Altered tomato (Lycopersicon esculentum Mill.) fruit cuticle biomechanics of a pleiotropic non ripening mutant. J. Plant Growth Regul. 2004, 23, 61–75. [Google Scholar] [CrossRef]
- Kosma, D.K.; Parsons, E.P.; Isaacson, T.; Lü, S.; Rose, J.K.; Jenks, M.A. Fruit cuticle lipid composition during development in tomato ripening mutants. Physiol. Plant. 2010, 139, 107–117. [Google Scholar] [CrossRef]
- Benítez, J.J.; Guzmán-Puyol, S.; Vilaplana, F.; Heredia-Guerrero, J.A.; Domínguez, E.; Heredia, A. Mechanical performances of isolated cuticles along tomato fruit growth and ripening. Front. Plant Sci. 2021, 12, 787839. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DAA | Cuticular Cracks’ Severity z | Fruit Weight (g) y | CM Deposition (mg·cm−2) y | ||||
|---|---|---|---|---|---|---|---|
| 2022 Summer–Autumn | 2023 Spring–Summer | 2023 Summer–Autumn | 2022 Summer–Autumn | 2023 Spring–Summer x | 2023 Summer–Autumn | ||
| 10 | Grade 0 | 4.42 ± 0.12 | 1.57 ± 0.09 | 3.55 ± 0.10 | 0.88 ± 0.03 | N.D. | 0.98 ± 0.03 |
| Grade 1 | 5.41 | N.D. | N.D. | 1.11 | N.D. | N.D. | |
| Grade 2 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | |
| Grade 3 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | |
| Grade 4 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | |
| 20 | Grade 0 | 11.90 ± 0.26 a | 10.81 ± 0.24 | 11.25 ± 0.35 a | 1.96 ± 0.04 a | 1.60 ± 0.03 | 2.08 ± 0.02 a |
| Grade 1 | 12.32 ± 0.31 a | 11.11 | 11.44 ± 0.36 a | 2.06 ± 0.03 a | 1.72 | 2.10 ± 0.03 a | |
| Grade 2 | 12.11 ± 0.65 a | N.D. | 10.59 ± 0.16 a | N.D. | N.D. | 2.20 ± 0.24 a | |
| Grade 3 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | |
| Grade 4 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | |
| 30 | Grade 0 | N.D. | 17.51 ± 0.29 b | 9.91 | N.D. | 2.68 ± 0.03 b | 1.65 |
| Grade 1 | 17.01 ± 0.46 a | 19.65 ± 0.38 a | 13.18 ± 0.38 a | 2.06 ± 0.07 a | 2.87 ± 0.07 a | 2.01 ± 0.04 b | |
| Grade 2 | 18.22 ± 0.59 a | 18.57 | 13.43 ± 0.48 a | 2.15 ± 0.09 a | 18.57 | 2.19 ± 0.06 ab | |
| Grade 3 | 17.85 ± 0.38 a | N.D. | 13.81 ± 0.60 a | 2.05 ± 0.06 a | N.D. | 2.28 ± 0.03 a | |
| Grade 4 | 17.87 ± 0.74 a | N.D. | 12.82 ± 0.63 a | 1.98 | N.D. | 2.37 ± 0.10 a | |
| 40 | Grade 0 | N.D. | 18.17 ± 0.39 a | 9.60 | N.D. | 2.61 ± 0.03 a | 2.25 |
| Grade 1 | 21.69 ± 1.91 ab | 19.15 ± 0.35 a | 15.38 ± 0.76 a | 2.00 ± 0.09 b | 2.62 ± 0.03 a | 2.18 ± 0.06 a | |
| Grade 2 | 22.08 ± 1.15 a | 19.46 ± 0.69 a | 16.17 ± 0.64 a | 2.22 ± 0.05 b | 2.52 ± 0.05 a | 2.21 ± 0.05 a | |
| Grade 3 | 19.58 ± 0.43 ab | 20.32 | 17.57 ± 0.98 a | 2.34 ± 0.04 ab | N.D. | 2.29 ± 0.07 a | |
| Grade 4 | 18.70 ± 0.39 b | 16.78 ± 3.10 a | 16.95 ± 0.68 a | 2.50 ± 0.06 a | 2.79 ± 0.11 a | 2.39 ± 0.07 a | |
| Cultivation Season | DAA | Cuticular Cracks’ Occurrence | nz | Epidermal Cell Density (Number/Observed Area (275.53 µm × 206.65 µm)) y | |
|---|---|---|---|---|---|
| 2022 summer–autumn | 20 | Without cuticular cracks | 21 | 42.26 ± 2.03 | n.s. |
| With cuticular cracks | 31 | 39.35 ± 1.10 | |||
| 2023 spring–summer | 30 | Without cuticular cracks | 5 | 43.88 ± 4.44 | n.s. |
| With cuticular cracks | 5 | 38.08 ± 3.27 | |||
| 40 | Without cuticular cracks | 5 | 37.20 ± 2.79 | n.s. | |
| With cuticular cracks | 5 | 37.38 ± 3.38 | |||
| 2023 summer–autumn | 20 | Without cuticular cracks | 5 | 35.65 ± 2.13 | n.s. |
| With cuticular cracks | 5 | 33.70 ± 3.20 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosoi, R.; Ikeda, K.; Nishizawa, T.; Nabeshima, T. Time–Course Analysis of the Onset and Progression of Cuticle Cracking in Fruits of Cherry Tomato Cultivar ‘Nene’. Horticulturae 2025, 11, 89. https://doi.org/10.3390/horticulturae11010089
Hosoi R, Ikeda K, Nishizawa T, Nabeshima T. Time–Course Analysis of the Onset and Progression of Cuticle Cracking in Fruits of Cherry Tomato Cultivar ‘Nene’. Horticulturae. 2025; 11(1):89. https://doi.org/10.3390/horticulturae11010089
Chicago/Turabian StyleHosoi, Ryosuke, Kazuo Ikeda, Takashi Nishizawa, and Tomoyuki Nabeshima. 2025. "Time–Course Analysis of the Onset and Progression of Cuticle Cracking in Fruits of Cherry Tomato Cultivar ‘Nene’" Horticulturae 11, no. 1: 89. https://doi.org/10.3390/horticulturae11010089
APA StyleHosoi, R., Ikeda, K., Nishizawa, T., & Nabeshima, T. (2025). Time–Course Analysis of the Onset and Progression of Cuticle Cracking in Fruits of Cherry Tomato Cultivar ‘Nene’. Horticulturae, 11(1), 89. https://doi.org/10.3390/horticulturae11010089