
Purslane (Portulaca oleracea L.) Growth, Nutritional, and Antioxidant Status under Different Nitrogen Levels in Hydroponics





Abstract
1. Introduction
2. Materials and Methods
2.1. Cropping System, Plant Material, and Growth Conditions
2.2. Plant Growth and Tissue Analysis
2.3. Plant Physiology and Photosynthetic-Related Parameters
2.4. Plant Tissue Minerals and Water and Nutrient Use Efficiency
2.5. Total Phenols, Total Flavonoids, Antioxidant Activity, Ascorbic Acid, and Total Soluble Sugars
2.6. Lipid Peroxidation, Hydrogen Peroxide, and Enzyme Antioxidant Activity
2.7. Statistical Methods
3. Results
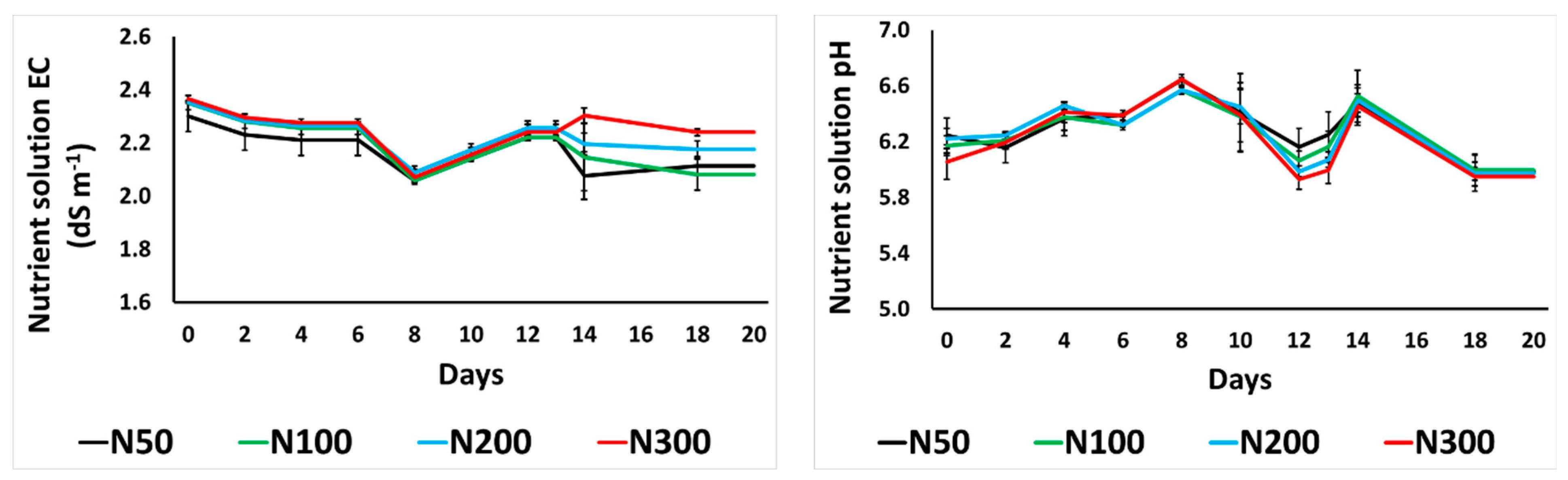
3.1. Electrical Conductivity and pH of the Drainage Nutrient Solution
3.2. Growth Parameters
3.3. Physiological Parameters
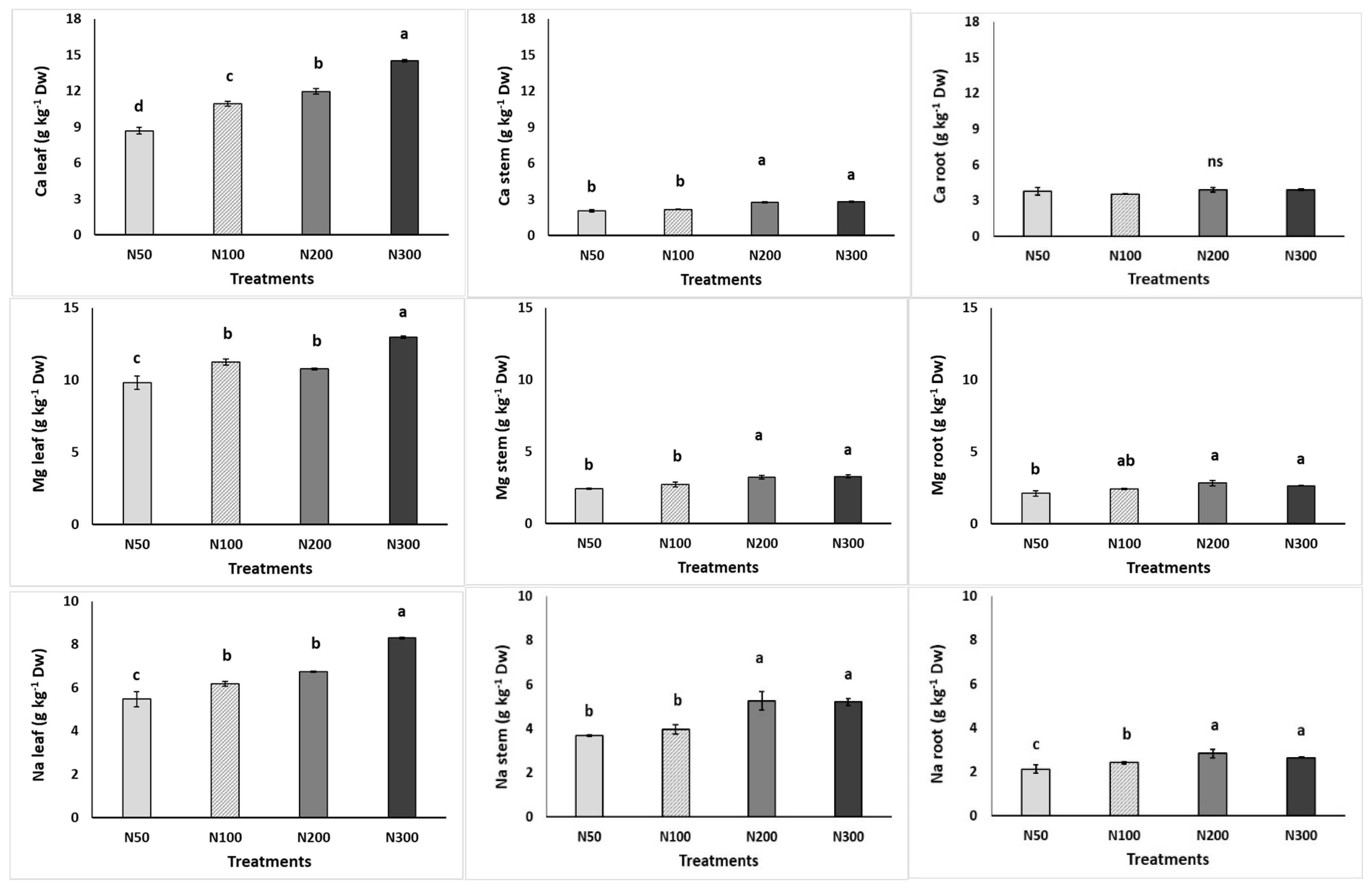
3.4. Leaf and Root Nutrient Content
3.5. Nitrogen and Water Uptake and Use Efficiency
3.6. Correlation of N Concentrations with Individual Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chugh, V.; Mishra, V.; Dwivedi, S.V.; Sharma, K.D. Purslane (Portulaca oleracea L.): An underutilized wonder plant with potential pharmacological value. Pharma Innov. J. 2019, 8, 236–246. [Google Scholar]
- Kumar, A.; Sreedharan, S.; Kashyap, A.K.; Singh, P.; Ramchiary, N. A review on bioactive phytochemicals and ethnopharmacological potential of purslane (Portulaca oleracea L.). Heliyon 2022, 8, e08669. [Google Scholar] [CrossRef] [PubMed]
- Cannavacciuolo, C.; Napolitano, A.; Heiss, E.H.; Dirsch, V.M.; Piacente, S. Portulaca oleracea, a rich source of polar lipids: Chemical profile by LC-ESI/LTQOrbitrap/MS/MSn and in vitro preliminary anti-inflammatory activity. Food Chem. 2022, 388, 132968. [Google Scholar] [CrossRef] [PubMed]
- Jalali, J.; Rahbardar, M.G. Ameliorative effects of Portulaca oleracea L. (purslane) on the metabolic syndrome: A review. J. Ethnopharmacol. 2022, 299, 115672. [Google Scholar] [CrossRef] [PubMed]
- D’Imperio, M.; Durante, M.; Gonnella, M.; Renna, M.; Montesano, F.F.; Parente, A.; Mita, G.; Serio, F. Enhancing the nutritional value of Portulaca oleracea L. by using soilless agronomic biofortification with zinc. Food Res. Int. 2022, 155, 111057. [Google Scholar] [CrossRef] [PubMed]
- Montoya-García, C.O.; García-Mateos, R.; Becerra-Martínez, E.; Toledo-Aguilar, R.; Volke-Haller, V.H.; Jesús Magdaleno-Villar, J. Bioactive compounds of purslane (Portulaca oleracea L.) according to the production system: A review. Sci. Hortic. 2023, 308, 111584. [Google Scholar] [CrossRef]
- Srivastava, R.; Srivastava, V.; Singh, A. Multipurpose Benefits of an Underexplored Species Purslane (Portulaca oleracea L.): A Critical Review. Environ. Manage. 2021, 72, 309–320. [Google Scholar] [CrossRef]
- Hnilickova, H.; Kraus, K.; Vachova, P.; Hnilicka, F. Salinity stress affects photosynthesis, malondialdehyde formation, and proline content in Portulaca oleracea L. Plants 2021, 10, 845. [Google Scholar] [CrossRef]
- de Lacerda, L.P.; Lange, L.C.; Costa França, M.G.; Zonta, E. Salinity Reduction and Biomass Accumulation in Hydroponic Growth of Purslane (Portulaca oleracea). Int. J. Phytoremediat. 2015, 17, 235–241. [Google Scholar] [CrossRef]
- Kong, Y.; Zheng, Y. Hydroponic production of purslane as a sodium-removing vegetable in NaCl-rich nutrient solution. HortScience 2014, 49, 201–206. [Google Scholar] [CrossRef]
- Camalle, M.; Standing, D.; Jitan, M.; Muhaisen, R.; Bader, N.; Bsoul, M.; Ventura, Y.; Soltabayeva, A.; Sagi, M. Effect of salinity and nitrogen sources on the leaf quality, biomass, and metabolic responses of two ecotypes of Portulaca oleracea. Agronomy 2020, 10, 656. [Google Scholar] [CrossRef]
- Peng, J.; Wei, W.; Lu, H.C.; Chen, W.; Li, S.D.; Wang, J.; Duan, C.Q.; He, F. Effect of Covering Crops between Rows on the Vineyard Microclimate, Berry Composition and Wine Sensory Attributes of ‘Cabernet Sauvignon’ (Vitis vinifera L. cv.) Grapes in a Semi-Arid Climate of Northwest China. Horticulturae 2022, 8, 518. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Hajisolomou, E.; Xylia, P.; Tzortzakis, N. Olive-mill and grape-mill waste as a substitute growing media component for unexploded vegetables production. Sustain. Chem. Pharm. 2023, 31, 100940. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Louka, S.; Petropoulos, S.A.; Tzortzakis, N. Soilless Cultivation of Portulaca oleracea Using Medicinal and Aromatic Plant Residues for Partial Peat Replacement. Horticulturae 2023, 9, 474. [Google Scholar] [CrossRef]
- Atrooz, O.M.; Al-Maitah, S.Z. Characterization of the crude extract of Portulaca oleracea and the determination of the polyphenol oxidase kinetics in the presence of Cu and Zn. J. Appl. Biol. Biotechnol. 2022, 10, 28–33. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Hajisolomou, E.; Xylia, P.; Tzortzakis, N. Ammonium to total nitrogen ratio affects the purslane (Portulaca oleracea L.) growth, nutritional, and antioxidant status. Heliyon 2023, 9, e21644. [Google Scholar] [CrossRef]
- Szalai, G.; Dai, N.; Danin, A.; Dudai, N.; Barazani, O. Effect of nitrogen source in the fertilizing solution on nutritional quality of three members of the Portulaca oleracea aggregate. J. Sci. Food Agric. 2010, 90, 2039–2045. [Google Scholar] [CrossRef]
- Montoya-García, C.O.; Volke-Haller, V.H.; Trinidad-Santos, A.; Villanueva-Verduzco, C. Change in the contents of fatty acids and antioxidant capacity of purslane in relation to fertilization. Sci. Hortic. 2018, 234, 152–159. [Google Scholar] [CrossRef]
- Mancosu, N.; Snyder, R.L.; Kyriakakis, G.; Spano, D. Water scarcity and future challenges for food production. Water 2015, 7, 975–992. [Google Scholar] [CrossRef]
- Varlagas, H.; Savvas, D.; Mouzakis, G.; Liotsos, C.; Karapanos, I.; Sigrimis, N. Modelling uptake of Na+ and Cl− by tomato in closed-cycle cultivation systems as influenced by irrigation water salinity. Agric. Water Manag. 2010, 97, 1242–1250. [Google Scholar] [CrossRef]
- Savvas, D.; Neocleous, D. Developments in soilless/hydroponic cultivation of vegetables. In Achieving Sustainable Cultivation of Vegetables; Hochmuth, G., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 211–243. [Google Scholar]
- Tzortzakis, N.; Nicola, S.; Savvas, D.; Voogt, W. Editorial: Soilless Cultivation Through an Intensive Crop Production Scheme. Management Strategies, Challenges and Future Directions. Front. Plant Sci. 2020, 11, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Fei, L.; Huang, D.; Zeng, J.; Chen, L.; Cai, Y. Coupling effects of irrigation and nitrogen levels on yield, water and nitrogen use efficiency of surge-root irrigated jujube in a semiarid region. Agric. Water Manag. 2019, 213, 146–154. [Google Scholar] [CrossRef]
- Akl, I.A.; Savvas, D.; Papadantonakis, N.; Lydakis-Simantiris, N.; Kefalas, P. Influence of Ammonium to Total Nitrogen Supply Ratio on Growth, Yield and Fruit Quality of Tomato Grown in a Closed Hydroponic System. Eur. J. Hortic. Sci. 2003, 68, 204–211. [Google Scholar]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef]
- Castellanos-Morales, V.; Villegas-Moreno, J.; Vierheilig, H.; Cárdenas-Navarro, R. Nitrogen availability drives the effect of Glomus intraradices on the growth of strawberry (Fragaria x ananassa Duch.) plants. J. Sci. Food Agric. 2012, 92, 2260–2264. [Google Scholar] [CrossRef]
- Liu, R.; Yang, Y.; Wang, Y.-S.; Wang, X.-C.; Rengel, Z.; Zhang, W.-J.; Shu, L.-Z. Alternate partial root-zone drip irrigation with nitrogen fertigation promoted tomato growth, water and fertilizer-nitrogen use efficiency. Agric. Water Manag. 2020, 233, 106049. [Google Scholar] [CrossRef]
- Mahajan, M.; Pal, P.K. Yield response, accumulation of bioactive ingredient and ion uptake of Stevia rebaudiana to different soil-moisture and nitrogen levels. Agric. Water Manag. 2022, 264, 107511. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Nikolaidou, E.; Stamatakis, A.; Tzortzakis, N. Vegetative, physiological, nutritional and antioxidant behavior of spearmint (Mentha spicata L.) in response to different nitrogen supply in hydroponics. J. Appl. Res. Med. Aromat. Plants 2017, 6, 52–61. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Panayiotou, C.; Tzortzakis, N. Nitrogen and phosphorus levels affected plant growth, essential oil composition and antioxidant status of lavender plant (Lavandula angustifolia Mill.). Ind. Crop. Prod. 2016, 83, 577–586. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Tzortzakis, N. Optimising fertigation of hydroponically grown sowthistle (Sonchus oleraceus L.): The impact of the nitrogen source and supply concentration. Agric. Water Manag. 2023, 289, 108528. [Google Scholar] [CrossRef]
- Richardson, A.D.; Duigan, S.P.; Berlyn, G.P. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol. 2002, 153, 185–194. [Google Scholar] [CrossRef]
- Wellburn, A.R.A.R. The spectral determination of Chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Papakyriakou, E.; Petropoulos, S.A.; Tzortzakis, N. The combined and single effect of salinity and copper stress on growth and quality of Mentha spicata plants. J. Hazard. Mater. 2019, 368, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Benimeli, C.S.; Medina, A.; Navarro, C.M.; Medina, R.B.; Amoroso, M.J.; Gómez, M.I. Bioaccumulation of copper by Zea mays: Impact on root, shoot and leaf growth. Water Air Soil Pollut. 2010, 210, 365–370. [Google Scholar] [CrossRef]
- Amin, H.; Arain, B.A.; Jahangir, T.M.; Abbasi, A.R.; Mangi, J.; Abbasi, M.S.; Amin, F. Copper (Cu) tolerance and accumulation potential in four native plant species: A comparative study for effective phytoextraction technique. Geol. Ecol. Landsc. 2021, 5, 53–64. [Google Scholar] [CrossRef]
- Azooz, M.M.; Abou-Elhamd, M.F.; Al-Fredan, M.A. Biphasic effect of copper on growth, proline, lipid peroxidation and antioxidant enzyme activities of wheat (Triticum aestivum cv. Hasaawi) at early growing stage. Aust. J. Crop. Sci. 2012, 6, 688–694. [Google Scholar]
- Fernández, J.E.; Alcon, F.; Diaz-Espejo, A.; Hernandez-Santana, V.; Cuevas, M.V. Water use indicators and economic analysis for on-farm irrigation decision: A case study of a super high density olive tree orchard. Agric. Water Manag. 2020, 237, 106074. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Kloukina, C.; Vassiliou, R.; Tomou, E.-M.E.-M.; Skaltsa, H.; Tzortzakis, N. Cultivation strategy to improve chemical profile and anti-oxidant activity of Sideritis perfoliata L. subsp. perfoliata. Ind. Crop. Prod. 2019, 140, 111694. [Google Scholar] [CrossRef]
- Meyers, K.J.; Watkins, C.B.; Pritts, M.P.; Liu, R.H. Antioxidant and antiproliferative activities of strawberries. J. Agric. Food Chem. 2003, 51, 6887–6892. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 18th ed.; AOAC: Gaithersburg, MD, USA, 2007. [Google Scholar]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- de Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; de Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Botsaris, G.; Tzortzakis, N. Antioxidant and antibacterial activities, mineral and essential oil composition of spearmint (Mentha spicata L.) affected by the potassium levels. Ind. Crop. Prod. 2017, 103, 202–212. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Charalambous, S.; Xylia, P.; Litskas, V.; Stavrinides, M.; Tzortzakis, N. Assessing the biostimulant effects of a novel plant-based formulation on tomato crop. Sustainability 2020, 12, 8432. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Massa, D.; Vandecasteele, B. The Tripartite of Soilless Systems, Growing Media, and Plants through an Intensive Crop Production Scheme. Agronomy 2022, 12, 1896. [Google Scholar] [CrossRef]
- Wenceslau, D.d.S.L.; de Oliveira, D.F.; deO Rabelo, H.; Ferbonink, G.F.; Gomes, L.A.A.; Leonel, É.C.A.; Caione, G. Nitrate concentration and nitrate/ammonium ratio on lettuce grown in hydroponics in Southern Amazon. Afr. J. Agric. Res. 2021, 17, 862–868. [Google Scholar]
- Sapkota, S.; Sapkota, S.; Liu, Z. Effects of nutrient composition and lettuce cultivar on crop production in hydroponic culture. Horticulturae 2019, 5, 72. [Google Scholar] [CrossRef]
- Zhao, D.; Oosterhuis, D.M.; Bednarz, C.W. Influence of potassium deficiency on photosynthesis, chlorophyll content, and chloroplast ultrastructure of cotton plants. Photosynthetica 2001, 39, 103–109. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Straltsova, D.; Medvedev, S.S.; Pozhvanov, G.A.; Sokolik, A.; Yurin, V. Stress-induced electrolyte leakage: The role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef] [PubMed]
- Gajewska, E.; Skłodowska, M. Differential effect of equal copper, cadmium and nickel concentration on biochemical reactions in wheat seedlings. Ecotoxicol. Environ. Saf. 2010, 73, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Phytochemical composition and bioactive compounds of common purslane (Portulaca oleracea L.) as affected by crop management practices. Trends Food Sci. Technol. 2016, 55, 1–10. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, W.; Ying, X.; Stien, D. New flavonoids from Portulaca oleracea L. and their activities. Fitoterapia 2018, 127, 257–262. [Google Scholar] [CrossRef]
- Fraser, C.M.; Chapple, C. The Phenylpropanoid Pathway in Arabidopsis. Arab. B. 2011, 9, e0152. [Google Scholar] [CrossRef]
- Nguyen, P.M.; Niemeyer, E.D. Effects of nitrogen fertilization on the phenolic composition and antioxidant properties of basil (Ocimum basilicum L.). J. Agric. Food Chem. 2008, 56, 8685–8691. [Google Scholar] [CrossRef]
- Verardo, V.; Riciputi, Y.; Sorrenti, G.; Ornaghi, P.; Marangoni, B.; Caboni, M.F. Effect of nitrogen fertilisation rates on the content of fatty acids, sterols, tocopherols and phenolic compounds, and on the oxidative stability of walnuts. LWT 2013, 50, 732–738. [Google Scholar] [CrossRef]
- He, J.; You, X.; Qin, L. High Salinity Reduces Plant Growth and Photosynthetic Performance but Enhances Certain Nutritional Quality of C4 Halophyte Portulaca oleracea L. Grown Hydroponically Under LED Lighting. Front. Plant Sci. 2021, 12, 651341. [Google Scholar] [CrossRef]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol 1999, 50, 641–664. [Google Scholar] [CrossRef] [PubMed]
- Kaymak, H.C. Effect of nitrogen forms on growth, yield and nitrate accumulation of cultivated purslane (Portulaca oleracea L.). Bulg. J. Agric. Sci. 2013, 19, 444–449. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Parameters | StarterNS | N50 | N100 | N200 | N300 |
|---|---|---|---|---|---|
| EC dS m−1 | 1.75 | 2.32 | 2.33 | 2.34 | 2.36 |
| K+ mmol L−1 | 7.55 | 8.95 | 8.95 | 8.95 | 8.95 |
| Ca2+ mmol L−1 | 3.50 | 3.74 | 3.74 | 3.74 | 3.74 |
| Mg2+ mmol L−1 | 1.00 | 2.88 | 2.88 | 2.88 | 2.98 |
| NH4+ mmol L−1 | 0.50 | 0.21 | 0.42 | 0.83 | 1.23 |
| NO3− mmol L−1 | 13.72 | 3.37 | 6.73 | 13.46 | 20.2 |
| SO42− mmol L−1 | 1.29 | 8.07 | 6.49 | 2.92 | 0.35 |
| H2PO4− mmol L−1 | 1.80 | 2.26 | 2.26 | 2.26 | 2.26 |
| Cl− mmol L−1 | 1.49 | 1.49 | 1.49 | 1.49 | 1.49 |
| Fe μmol L−1 | 30.00 | 71.56 | 71.56 | 71.56 | 71.56 |
| Mn μmol L−1 | 5.00 | 18.21 | 18.21 | 18.21 | 18.21 |
| Zn μmol L−1 | 4.00 | 1.53 | 1.53 | 1.53 | 1.53 |
| Cu μmol L−1 | 1.00 | 4.72 | 4.72 | 4.72 | 4.72 |
| B μmol L−1 | 30.00 | 18.52 | 18.52 | 18.52 | 18.52 |
| Mo μmol L−1 | 0.50 | 0.52 | 0.52 | 0.52 | 0.52 |
| N Concentration | Plant Height | Number of Leaves | Leaf FW | Stem FW | Root FW | Plant Biomass FW | Leaf DW | Stem DW | Root DW | Plant Biomass DW | Leaf DM | Stem DM | Root DM |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N50 | 40.1 ± 0.7 a | 270.5 ± 17.4 a | 47.1 ± 2.7 a | 61.7 ± 10.1 a | 10.5 ± 1.0 a | 123.3 ± 7.1 a | 1.87 ± 0.11 a | 3.27 ± 0.21 a | 0.87 ± 0.09 a | 5.15 ± 0.32 a | 10.95 ± 0.27 a | 7.26 ± 2.67 a | 8.26 ± 0.16 a |
| N100 | 42.5 ± 1.2 a | 217.6 ± 17.4 ab | 41.6 ± 2.3 a | 62.3 ± 2.8 a | 8.80 ± 0.5 ab | 108.7 ± 4.7 ab | 1.72 ± 0.08 a | 2.73 ± 0.14 ab | 0.66 ± 0.04 b | 4.45 ± 0.21 a | 10.79 ± 0.41 a | 4.38 ± 0.13 a | 7.49 ± 0.10 b |
| N200 | 39.2 ± 1.9 a | 201.5 ± 24.4 b | 50.4 ± 9.8 a | 57.3 ± 4.9 a | 6.88 ± 0.5 b | 101.2 ± 8.1 ab | 1.51 ± 0.12 a | 2.55 ± 0.26 b | 0.56 ± 0.04 b | 4.07 ± 0.37 a | 9.25 ± 1.34 a | 4.41 ± 0.09 a | 8.21 ± 0.14 a |
| N300 | 38.8 ± 1.5 a | 201.6 ± 18.6 b | 38.8 ± 25.5 a | 52.6 ± 5.9 a | 7.72 ± 0.7 b | 95.6 ± 11.1 b | 1.62 ± 0.19 a | 2.42 ± 0.28 b | 0.63 ± 0.05 b | 4.04 ± 0.47 a | 10.45 ± 0.39 a | 4.59 ± 0.06 a | 8.34 ± 0.43 a |
| N Concentration | Stomatal Conductance | SPAD | Chlorophyll Fluorescence | Chlorophyll a | Chlorophyll b | Total Chlorophyll | Carotenoids | Electrolyte Leakage |
|---|---|---|---|---|---|---|---|---|
| N50 | 36.3 ± 12.1 ab | 29.7 ± 0.8 a | 0.769 ± 0.003 b | 0.287 ± 0.022 a | 0.044 ± 0.003 a | 0.332 ± 0.025 a | 0.054 ± 0.003 a | 33.2 ± 2.3 a |
| N100 | 37.5 ± 6.6 ab | 30.7 ± 0.9 a | 0.771 ± 0.006 b | 0.310 ± 0.010 a | 0.050 ± 0.000 a | 0.357 ± 0.011 a | 0.058 ± 0.002 a | 27.5 ± 2.9 ab |
| N200 | 50.4 ± 9.6 a | 32.4 ± 1.1 a | 0.784 ± 0.003 a | 0.342 ± 0.025 a | 0.055 ± 0.002 a | 0.397 ± 0.029 a | 0.057 ± 0.005 a | 29.3 ± 3.5 ab |
| N300 | 19.7 ± 6.2 b | 30.8 ± 0.4 a | 0.775 ± 0.002 ab | 0.257 ± 0.064 a | 0.040 ± 0.010 a | 0.300 ± 0.073 a | 0.049 ± 0.011 a | 22.8 ± 1.4 b |
| N Concentration | Total Phenols | DPPH | FRAP | ABTS | Flavonoids | Ascorbic Acid | TSS |
|---|---|---|---|---|---|---|---|
| N50 | 0.29 ± 0.01 a | 0.205 ± 0.011 a | 0.269 ± 0.013 a | 0.089 ± 0.004 b | 0.219 ± 0.009 a | 4.11 ± 0.36 b | 2.00 ± 0.00 a |
| N100 | 0.25 ± 0.01 b | 0.208 ± 0.012 a | 0.245 ± 0.006 a | 0.092 ± 0.005 b | 0.251 ± 0.023 a | 5.19 ± 0.31 ab | 1.86 ± 0.03 b |
| N200 | 0.24 ± 0.02 b | 0.165 ± 0.008 b | 0.212 ± 0.010 b | 0.080 ± 0.005 b | 0.201 ± 0.017 a | 5.07 ± 0.34 ab | 1.90 ± 0.00 b |
| N300 | 0.30 ± 0.01 a | 0.213 ± 0.010 a | 0.262 ± 0.011 a | 0.106 ± 0.001 a | 0.114 ± 0.020 b | 5.42 ± 0.33 a | 1.90 ± 0.01 b |
| N Concentration | H2O2 | MDA | SOD | CAT | POD |
|---|---|---|---|---|---|
| N50 | 0.141 ± 0.005 bc | 6.07 ± 0.26 a | 0.477 ± 0.021 b | 9.81 ± 0.20 a | 0.800 ± 0.056 a |
| N100 | 0.171 ± 0.005 a | 4.99 ± 0.16 b | 0.475 ± 0.012 b | 10.10 ± 0.25 a | 0.880 ± 0.041 a |
| N200 | 0.131 ± 0.004 c | 5.80 ± 0.31 ab | 0.631 ± 0.010 a | 9.32 ± 0.52 a | 0.584 ± 0.019 b |
| N300 | 0.157 ± 0.010 ab | 5.95 ± 0.24 a | 0.591 ± 0.016 a | 9.46 ± 0.88 a | 0.660 ± 0.016 b |
| N Concentration | Nutrient Uptake | Water Uptake | NUE | WUE |
|---|---|---|---|---|
| N50 | 29.8 ± 0.71 bc | 7.72 ± 0.44 a | 0.12 ± 0.01 a | 0.47 ± 0.08 b |
| N100 | 32.4 ± 1.78 ab | 7.53 ± 0.49 a | 0.13 ± 0.01 a | 0.59 ± 0.06 ab |
| N200 | 35.0 ± 0.54 a | 7.15 ± 0.08 a | 0.17 ± 0.03 a | 0.84 ± 0.16 a |
| N300 | 28.5 ± 0.02 c | 7.80 ± 0.39 a | 0.14 ± 0.02 a | 0.53 ± 0.04 ab |
| N Concentration | Accumulation Rate | Bioaccumulation Coefficient | Translocation Factor | Tolerance Indices | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaves | Stems | Roots | Leaves | Stems | Total Biomass | Plant Height | Leaf No | Fresh Leaves | Fresh Stems | ||
| N50 | 39.1 ± 0.2 a | 862.7 ± 10.6 a | 481.8 ± 3.2 a | 622.4 ± 10.7 a | 1.38 ± 0.04 a | 0.77 ± 0.01 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a |
| N100 | 25.4 ± 0.1 b | 437.5 ± 4.2 b | 227.9 ± 2.3 b | 330.9 ± 4.4 b | 1.32 ± 0.03 ab | 0.69 ± 0.01 b | 70.7 ± 3.1 b | 101.4 ± 2.9 a | 69.9 ± 5.6 b | 73.1 ± 4.1 a | 80.9 ± 3.7 b |
| N200 | 20.1 ± 0.1 d | 211.4 ± 2.6 c | 108.8 ± 0.9 c | 160.1 ± 1.3 c | 1.32 ± 0.02 ab | 0.68 ± 0.01 b | 79.4 ± 6.3 b | 93.6 ± 4.5 a | 64.8 ± 7.8 b | 88.8 ± 17.3 a | 74.5 ± 6.4 b |
| N300 | 22.5 ± 0.1 c | 144.7 ± 1.3 d | 83.1 ± 0.9 d | 112.8 ± 1.6 d | 1.28 ± 0.02 b | 0.74 ± 0.02 a | 62.3 ± 7.2 b | 92.6 ± 3.6 a | 64.8 ± 5.9 b | 68.2 ± 8.4 a | 68.4 ± 7.7 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrysargyris, A.; Xylia, P.; Zengin, G.; Tzortzakis, N. Purslane (Portulaca oleracea L.) Growth, Nutritional, and Antioxidant Status under Different Nitrogen Levels in Hydroponics. Horticulturae 2024, 10, 1007. https://doi.org/10.3390/horticulturae10091007
Chrysargyris A, Xylia P, Zengin G, Tzortzakis N. Purslane (Portulaca oleracea L.) Growth, Nutritional, and Antioxidant Status under Different Nitrogen Levels in Hydroponics. Horticulturae. 2024; 10(9):1007. https://doi.org/10.3390/horticulturae10091007
Chicago/Turabian StyleChrysargyris, Antonios, Panayiota Xylia, Gokhan Zengin, and Nikolaos Tzortzakis. 2024. "Purslane (Portulaca oleracea L.) Growth, Nutritional, and Antioxidant Status under Different Nitrogen Levels in Hydroponics" Horticulturae 10, no. 9: 1007. https://doi.org/10.3390/horticulturae10091007
APA StyleChrysargyris, A., Xylia, P., Zengin, G., & Tzortzakis, N. (2024). Purslane (Portulaca oleracea L.) Growth, Nutritional, and Antioxidant Status under Different Nitrogen Levels in Hydroponics. Horticulturae, 10(9), 1007. https://doi.org/10.3390/horticulturae10091007