

Transcriptome Profiling Reveals Potential Genes Involved in Salicylic Acid-Induced Arbutin Synthesis in Pear
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. SA Treatments
2.3. Hydroquinone Treatments
2.4. Measurement of Arbutin Content
2.5. RNA Extraction, cDNA Library Construction, and Sequencing
2.6. Quantitative Real-Time PCR Analysis
2.7. Transient Transformation of PbUGT72B1 into Pear
2.8. Statistical Analysis
3. Results
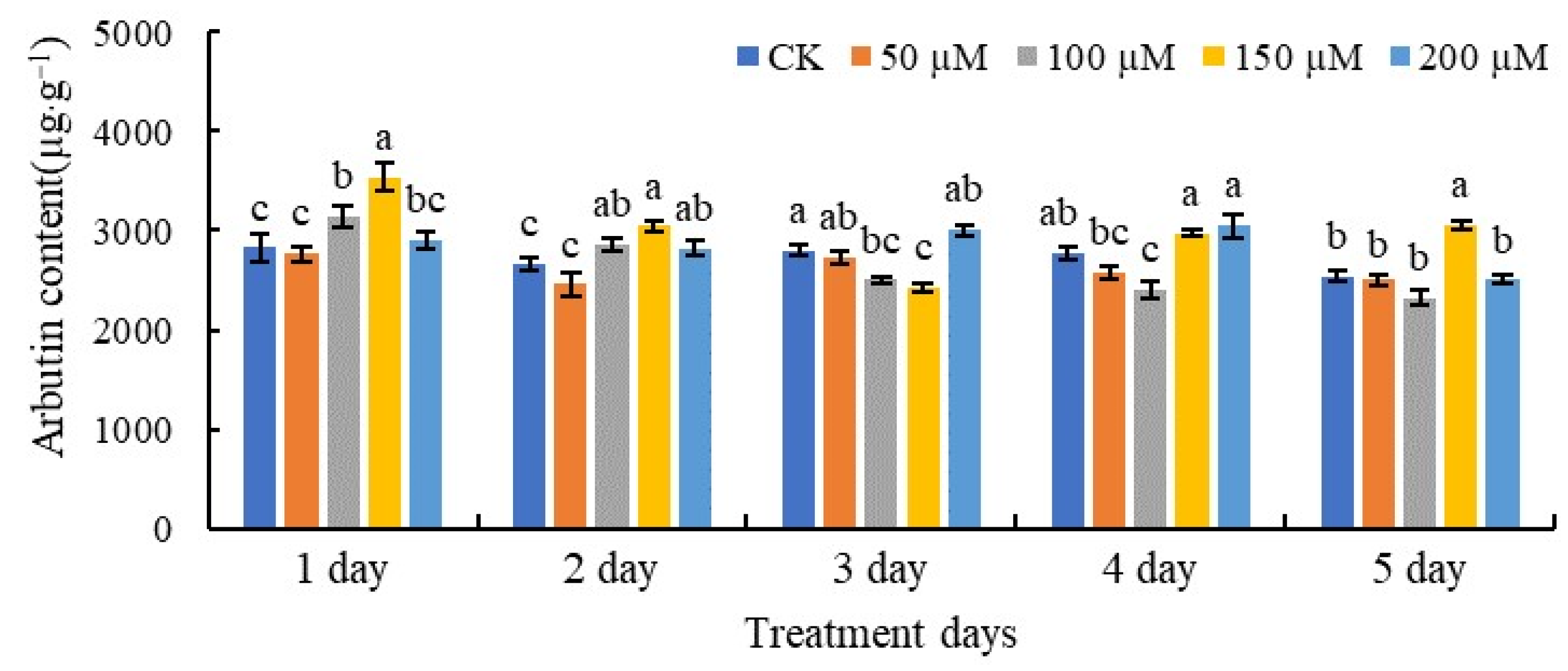
3.1. Arbutin Contents of Pear Leaves Treated with SA
3.2. Arbutin Contents of Fruits Treated with SA
3.3. Arbutin Contents of Calli Treated with SA
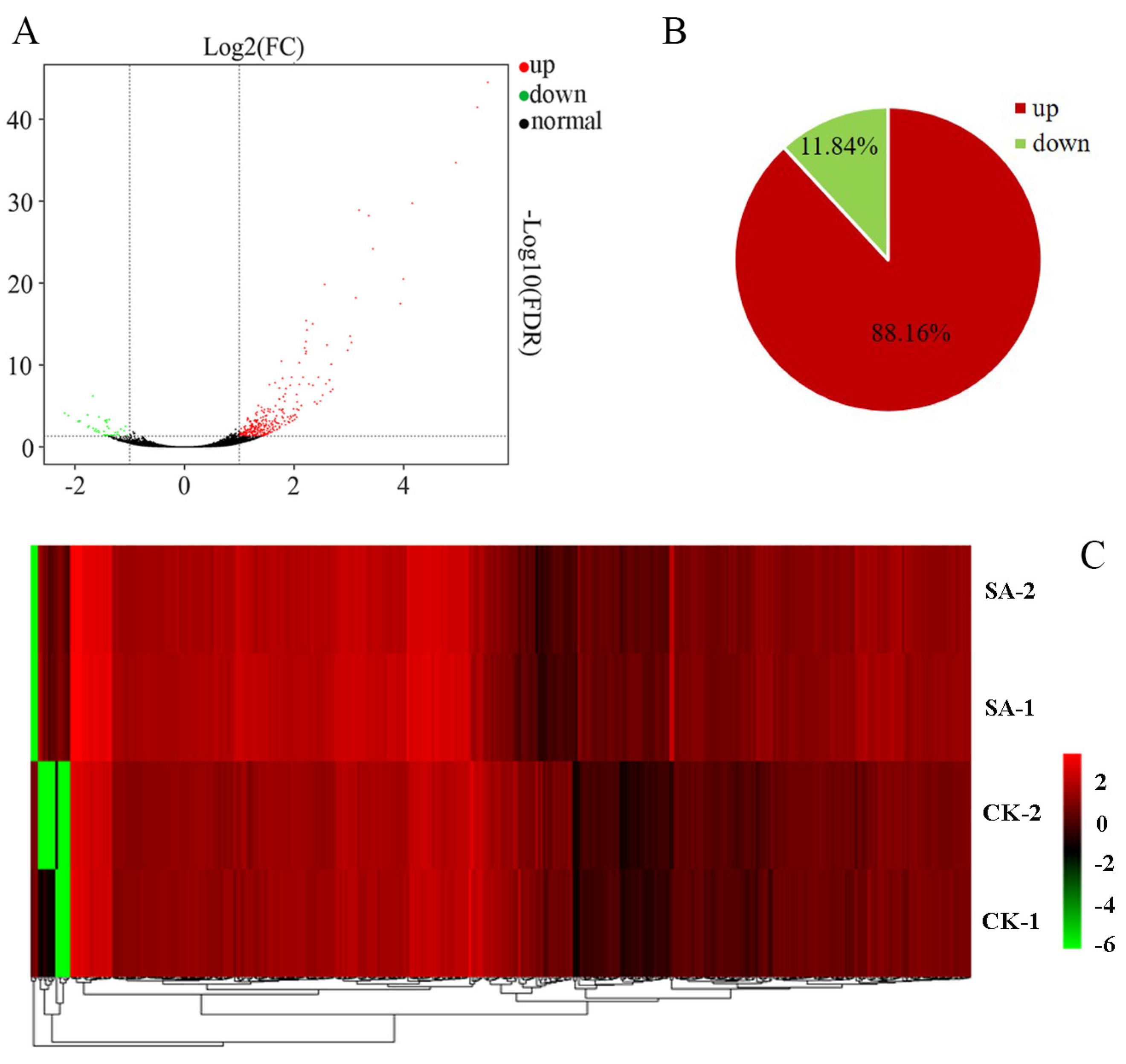
3.4. RNA-Seq of Pear Callus under SA Treatment
3.5. Analysis of DEGs in Response to SA
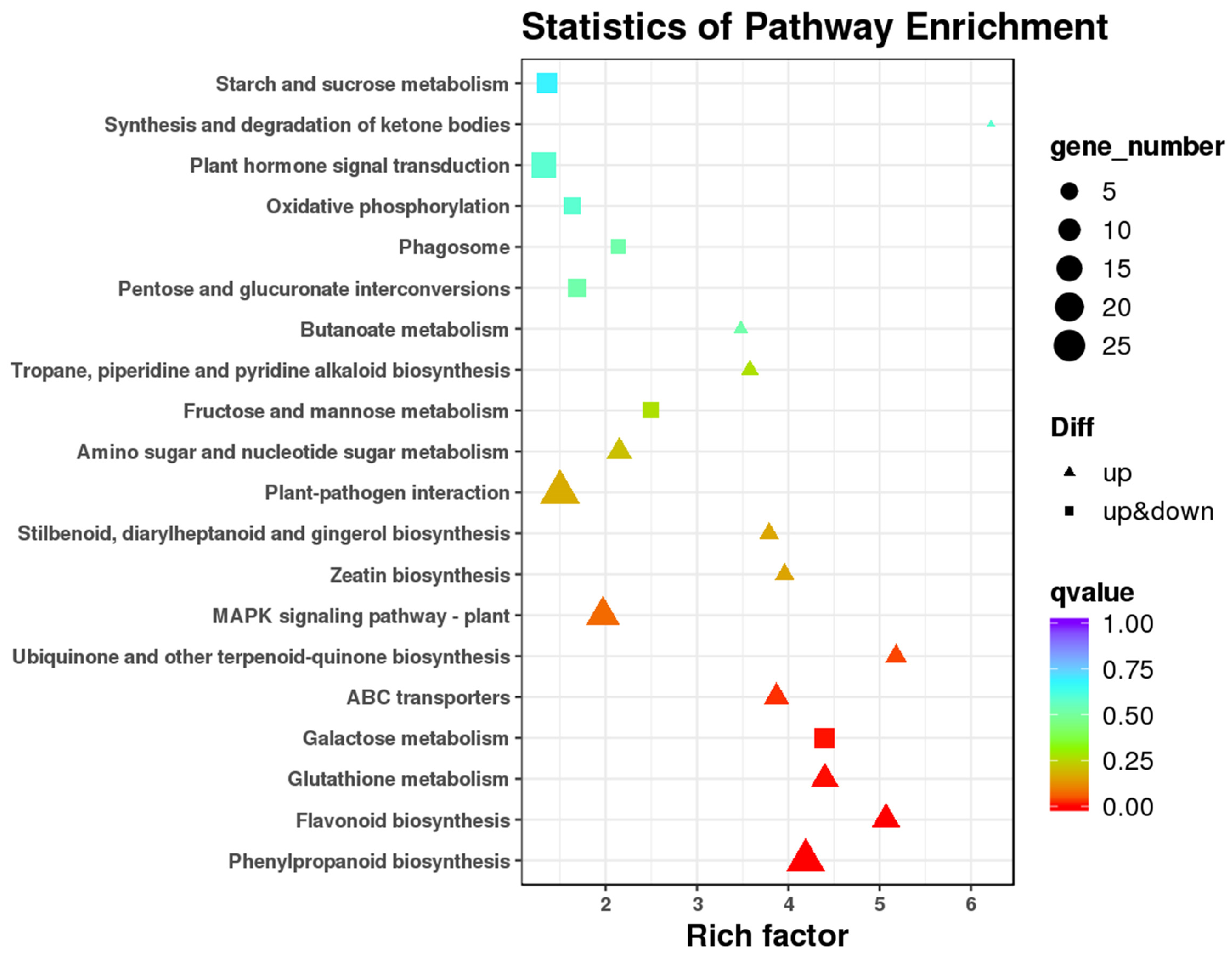
3.6. Functional Analysis of DEGs
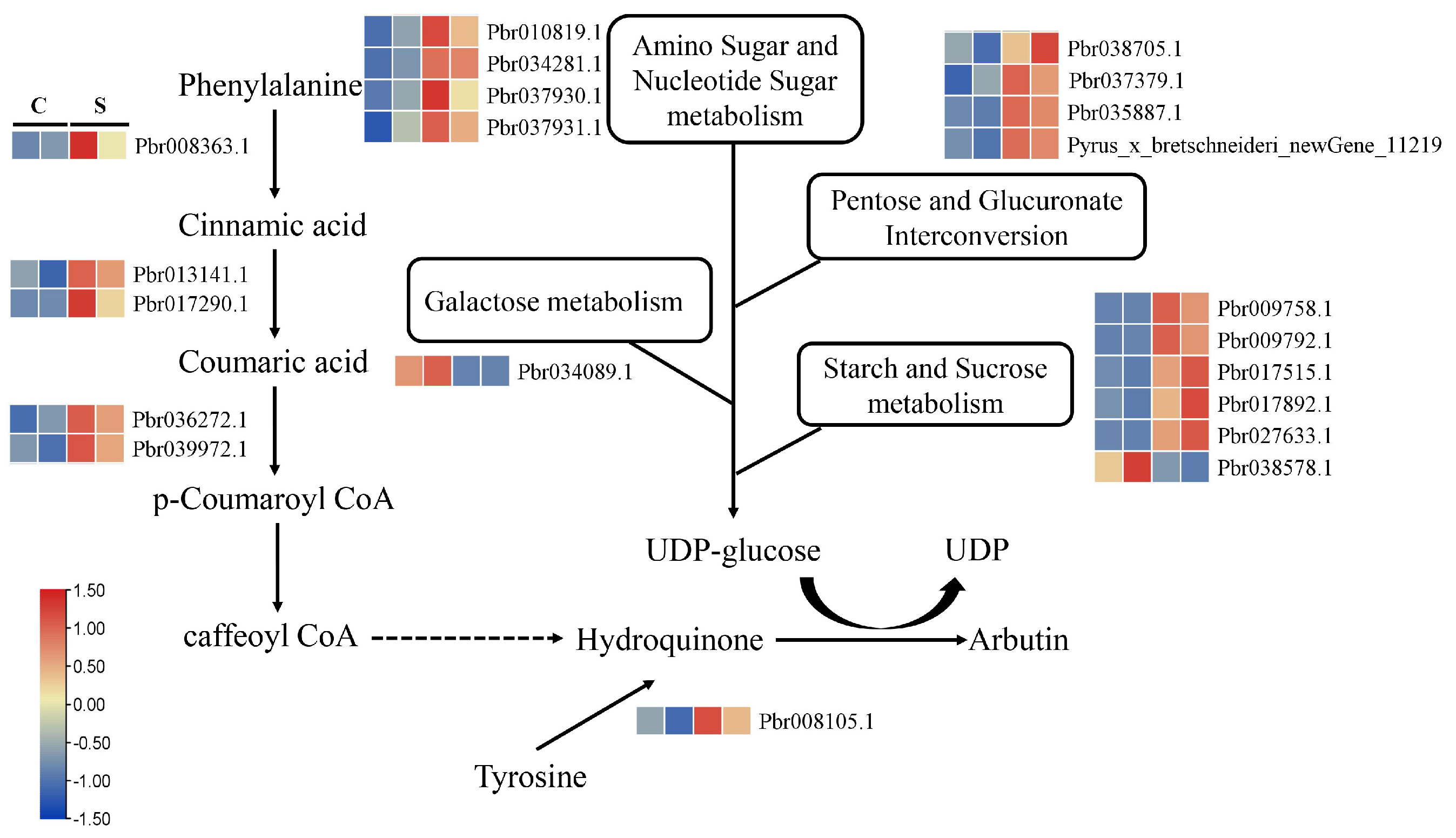
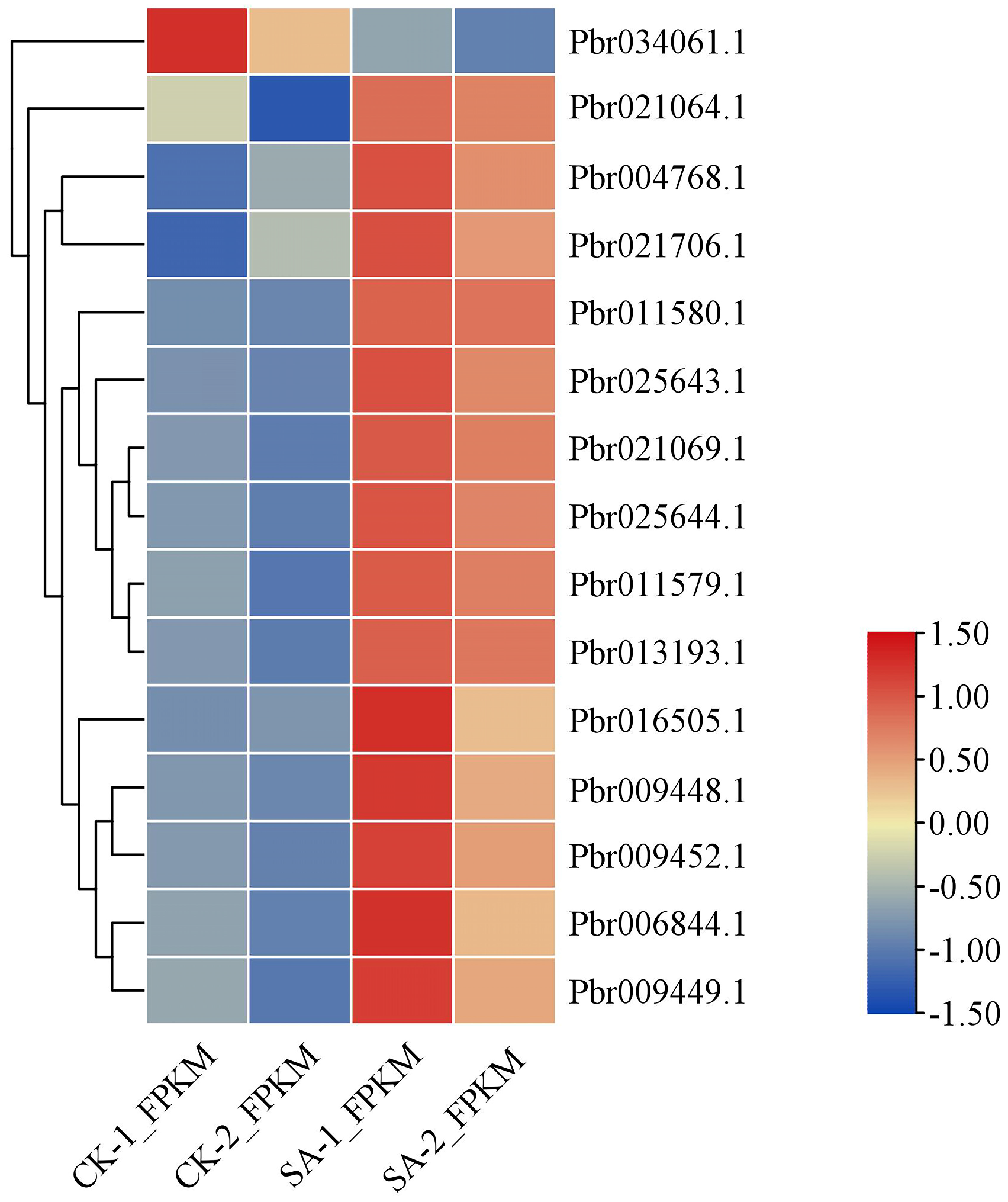
3.7. Key Genes in the Arbutin Biosynthesis Pathway
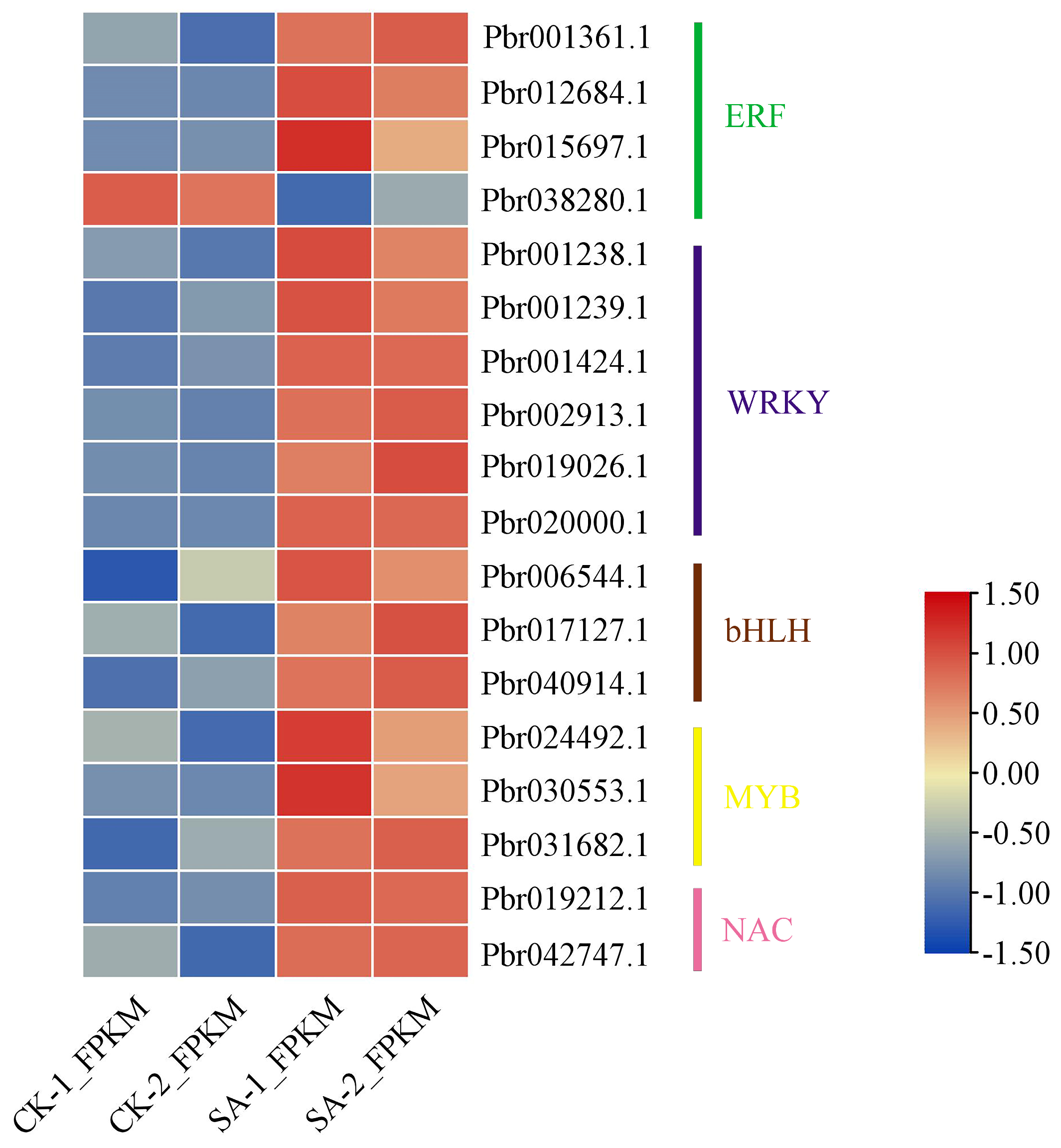
3.8. Transcription Factor Responses to SA Treatment
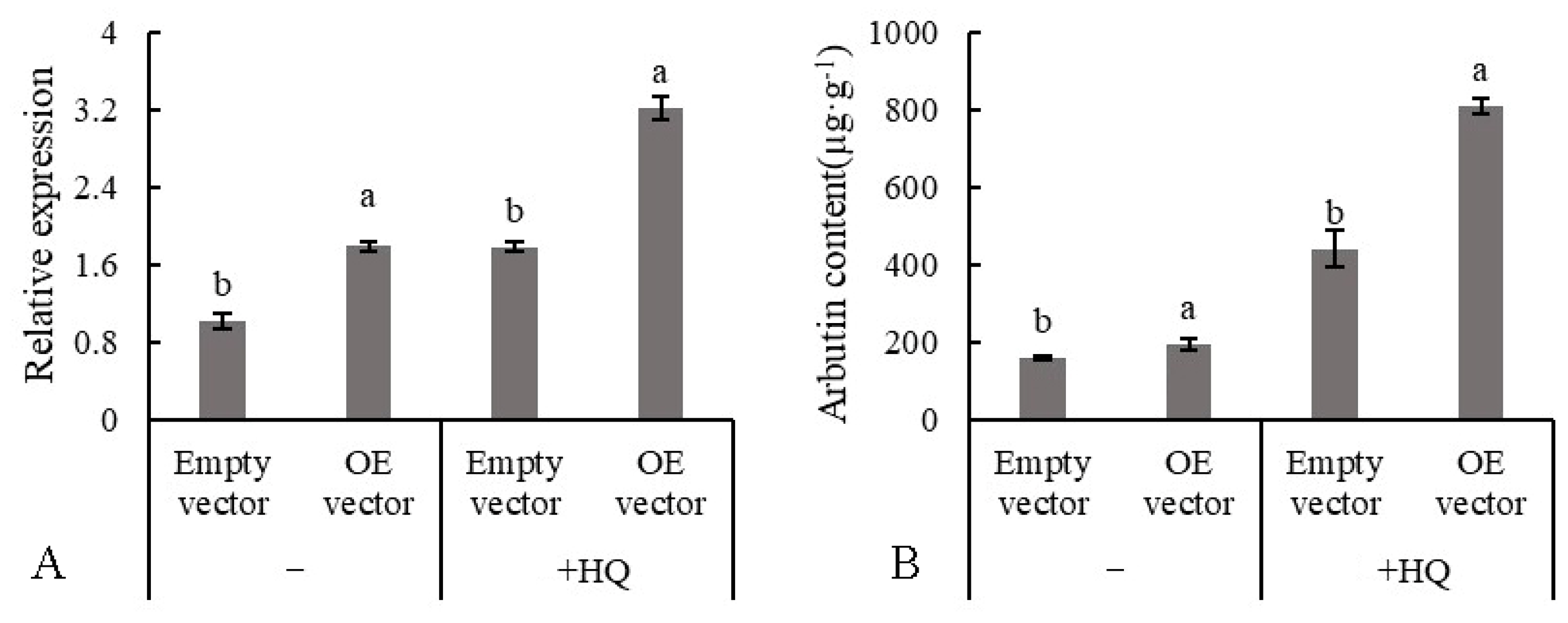
3.9. Effect of Hydroquinone Feeding on Arbutin Content and Gene Expression
3.10. Transient Expression of PbUGT72B1 in Pear Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, J.Y.; Fan, J.B.; Li, Q.H.; Jia, L.T.; Xu, L.L.; Wu, X.; Wang, Z.W.; Li, H.X.; Qi, K.J.; Qiao, X.; et al. Variation of organic acids in mature fruits of 193 pear (Pyrus spp.) cultivars. J. Food Compos. Anal. 2022, 109, 104483. [Google Scholar] [CrossRef]
- Hong, S.Y.; Lansky, E.; Kang, S.S.; Yang, M. A review of pears (Pyrus spp.), ancient functional food for modern time. BMC Complement. Med. Ther. 2021, 21, 219. [Google Scholar] [CrossRef] [PubMed]
- Reiland, H.; Slavin, J. Systematic Review of Pears and Health. Nutr. Today 2015, 50, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Navaei, N.; Pourafshar, S.; Akhavan, N.S.; Litwin, N.S.; Foley, E.M.; George, K.S.; Hartley, S.C.; Elam, M.L.; Rao, S.; Arjmandi, B.H.; et al. Influence of daily fresh pear consumption on biomarkers of cardiometabolic health in middle-aged/older adults with metabolic syndrome: A randomized controlled trial. Food Funct. 2019, 10, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Chandel, R.; Kumar, V.; Kaur, R.; Kumar, S.; Kumar, A.; Kumar, D.; Kapoor, S. Bioactive compounds, health benefits and valorization of Pyrus pyrifolia (sand pear): A review. Nutr. Food Sci. 2023, 53, 1061–1080. [Google Scholar] [CrossRef]
- Ulaszewska, M.; Vázquez-Manjarrez, N.; Garcia-Aloy, M.; Llorach, R.; Mattivi, F.O.; Dragsted, L.; Praticò, G.; Manach, C. Food intake biomarkers for apple, pear, and stone fruit. Genes Nutr. 2018, 13, 29. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.C.; Malnoy, M.; Hofmann, T.; Schwab, W.; Palmieri, L.; Wehrens, R.; Schuch, L.A.; Muller, M.; Schimmelpfeng, H.; Velasco, R.; et al. F1 hybrid of cultivated apple (Malus × domestica) and European pear (Pyrus communis) with fertile F2 offspring. Mol. Breed. 2014, 34, 817–828. [Google Scholar] [CrossRef]
- Funayama, M.; Arakaw, H.; Yamamoto, R.; Nishino, T.; Shin, T.; Murao, S. Effects of α- and β-Arbutin on Activity of Tyrosinases from Mushroom and Mouse Melanoma. Biosci. Biotechnol. Biochem. 1995, 59, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, T.T.; Zhou, B.; Gao, W.Y.; Cao, J.G.; Huang, L.Q. Chemical composition and antioxidant and anti-inflammatory potential of peels and flesh from 10 different pear varieties (Pyrus spp.). Food Chem. 2014, 152, 531–538. [Google Scholar] [CrossRef]
- Migas, P.; Krauze-Baranowska, M. The significance of arbutin and its derivatives in therapy and cosmetics. Phytochem. Lett. 2015, 13, 35–40. [Google Scholar] [CrossRef]
- Ragab, D.; Salah Eldin, H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 cytokine storm; what we know so far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef] [PubMed]
- Nahar, L.; Al-Groshi, A.; Kumar, A.; Sarker, S.D. Arbutin: Occurrence in Plants, and Its Potential as an Anticancer Agent. Molecules 2022, 27, 8786. [Google Scholar] [CrossRef]
- Jurica, K.; Gobin, I.; Kremer, D.; Čepo, D.V.; Grubešić, R.J.; Karačonji, I.B.; Kosalec, I. Arbutin and its metabolite hydroquinone as the main factors in the antimicrobial effect of strawberry tree (Arbutus unedo L.) leaves. J. Herb. Med. 2017, 8, 17–23. [Google Scholar] [CrossRef]
- Kuźniak, E.; Wielanek, M.; Chwatko, G.; Głowacki, R.; Libik-Konieczny, M.; Piątek, M.; Gajewska, E.; Skłodowska, M. Salicylic acid and cysteine contribute to arbutin-induced alleviation of angular leaf spot disease development in cucumber. J. Plant Physiol. 2015, 181, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Hou, K.; Zhang, H.H.; Wang, X.Y.; Wu, W. Integrating transcriptomics and metabolomics to studies key metabolism, pathways and candidate genes associated with drought-tolerance in Carthamus tinctorius L. under drought stress. Ind. Crops Prod. 2020, 151, 112465. [Google Scholar] [CrossRef]
- Lawas, L.M.F.; Li, X.; Erban, A.; Kopka, J.; Jagadish, S.V.K.; Zuther, E.; Hincha, D.K. Metabolic responses of rice cultivars with different tolerance to combined drought and heat stress under field conditions. GigaScience 2019, 8, giz050. [Google Scholar] [CrossRef] [PubMed]
- Grisdale, S.K.; Towers, G.H.N. Biosynthesis of Arbutin from Some Phenylpropanoid compounds in Pyrus communis. Nature 1960, 188, 1130–1131. [Google Scholar] [CrossRef]
- Arend, J.; Warzecha, H.; Hefner, T.; StÖckigt, J. Utilizing Genetically Engineered Bacteria to Produce Plant-Specific Glucosides. Biotechnol. Bioeng. 2001, 76, 126–131. [Google Scholar] [CrossRef]
- Hefner, T.; Arend, J.; Warzecha, H.; Siems, K.; StÖckigt, J. Arbutin Synthase, a Novel Member of the NRD1β Glycosyltransferase Family, is a Unique Multifunctional Enzyme Converting Various Natural Products and Xenobioticsy. Bioorg. Med. Chem. 2002, 10, 1731–1741. [Google Scholar] [CrossRef]
- Ali, B. Salicylic acid: An efficient elicitor of secondary metabolite production in plants. Biocatal. Agric. Biotechnol. 2021, 31, 101884. [Google Scholar] [CrossRef]
- Cui, K.B.; Shu, C.; Zhao, H.D.; Fan, X.G.; Cao, J.K. Preharvest chitosan oligochitosan and salicylic acid treatments enhance phenol metabolism and maintain the postharvest quality of apricots (Prunus armeniaca L.). Sci. Hortic. 2020, 267, 109334. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Liu, F.R.; Wang, J.J.; Yang, Q.R.; Wang, P.; Zhao, H.J.; Wang, J.; Wang, C.; Xu, X.H. Salicylic acid inhibits the postharvest decay of goji berry (Lycium barbarum L.) by modulating the antioxidant system and phenylpropanoid metabolites. Postharvest Biol. Technol. 2021, 178, 111558. [Google Scholar] [CrossRef]
- Ye, J.B.; Mao, D.; Cheng, S.Y.; Zhang, X.; Tan, J.P.; Zheng, J.R.; Xu, F. Comparative transcriptome analysis reveals the potential stimulatory mechanism of terpene trilactone biosynthesis by exogenous salicylic acid in Ginkgo biloba. Ind. Crops Prod. 2020, 145, 112104. [Google Scholar] [CrossRef]
- Chen, W.; Wang, Y.; Xu, L.; Dong, J.H.; Zhu, X.W.; Ying, J.L.; Wang, Q.J.; Fan, L.X.; Li, C.; Liu, L.W. Methyl jasmonate, salicylic acid and abscisic acid enhance the accumulation of glucosinolates and sulforaphane in radish (Raphanus sativus L.) taproot. Sci. Hortic. 2019, 250, 159–167. [Google Scholar] [CrossRef]
- Shi, H.Y.; Cao, L.W.; Yue, X.U.; Yang, X.; Liu, S.L.; Liang, Z.S. Transcriptional profiles underlying the effects of salicylic acid on fruit ripening and senescence in pear (Pyrus pyrifolia nakai). J. Integr. Agric. 2021, 20, 2424–2437. [Google Scholar] [CrossRef]
- Xu, Y.; Huo, L.Y.; Zhao, K.K.; Li, Y.W.; Zhao, X.R.; Wang, H.Y.; Wang, W.L.; Shi, H.Y. Salicylic acid delays pear fruit senescence by playing an antagonistic role toward ethylene, auxin, and glucose in regulating the expression of PpEIN3a. Front. Plant Sci. 2023, 13, 1096645. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, J.S.; Wang, Y.G.; Feng, W.X.; Xu, W.X. Effects of infection of epiphyte caused fruit rot on defense mechanism in pear fruit callus. Acta Bot. Boreali Occident. Sin. 2010, 30, 2219–2224. [Google Scholar]
- Skrzypczak-Pietraszek, E.; Kwiecien, I.; Pietraszek, J. HPLC-DAD analysis of arbutin produced from hydroquinone in a biotransformation process in Origanum majorana L. shoot culture. Phytochem. Lett. 2017, 20, 443–448. [Google Scholar] [CrossRef]
- Cui, T.T.; Li, J.H.; Li, N.; Li, F.; Song, Y.Q.; Li, L.L. Optimization of ultrasonic assisted extraction of arbutin from pear fruitlets using response surface methodology. J. Food Meas. Charact. 2022, 16, 3130–3139. [Google Scholar] [CrossRef]
- Fischer, T.C.; Gosch, C.; Pfeiffer, J.; Halbwirth, H.; Halle, C.; Stich, K.; Forkmann, G. Flavonoid genes of pear (Pyrus communis). Trees 2007, 21, 521–529. [Google Scholar] [CrossRef]
- Shang, Y.Z.; Wei, W.P.; Zhang, P.; Ye, B.C. Engineering Yarrowia lipolytica for enhanced production of arbutin. J. Agric. Food Chem. 2020, 68, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Rafi, S.; Kamili, A.N.; Ganai, B.A.; Parray, J.A.; Jan, S. Variation in root morphology, enhancement in anti-oxidative enzyme responses and improved arbutin and bergenin levels in Bergenia ciliata (Haw.) Sternb. raised in vitro via EMS and gamma irradiations. Plant Cell Tiss Organ Cult. 2021, 145, 43–57. [Google Scholar] [CrossRef]
- Zhang, L.M.; Kamitakahara, H.; Sasaki, R.; Oikawa, A.; Saito, K.; Murayama, H.; Ohsako, T.; Itai, A. Effect of exogenous GA4+7 and BA + CPPU treatments on fruit lignin and primary metabolites in Japanese pear “Gold Nijisseiki”. Sci. Hortic. 2020, 272, 109593. [Google Scholar] [CrossRef]
- Cao, X.Y.; Sun, H.L.; Wang, X.Y.; Li, W.X.; Wang, X.Q. ABA signaling mediates 5-aminolevulinic acid-induced anthocyanin biosynthesis in red pear fruits. Sci. Hortic. 2022, 304, 111290. [Google Scholar] [CrossRef]
- Dong, J.; Wan, G.W.; Liang, Z.S. Accumulation of salicylic acid-induced phenolic compounds and raised activities of secondary metabolic and antioxidative enzymes in Salvia miltiorrhiza cell culture. J. Biotechnol. 2010, 148, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Gomes, E.P.; Borges, C.V.; Monteiro, G.C.; Belin, M.A.F.; Minatel, I.O.; Junior, A.P.; Tecchio, M.A.; Lima, G.P.P. Preharvest salicylic acid treatments improve phenolic compounds and biogenic amines in ‘Niagara Rosada’ table grape. Postharvest Biol. Technol. 2021, 176, 111505. [Google Scholar] [CrossRef]
- Zhai, R.; Liu, X.T.; Feng, W.T.; Chen, S.S.; Xu, L.F.; Wang, Z.G.; Zhang, J.L.; Li, P.M.; Ma, F.W. Different Biosynthesis Patterns among Flavonoid 3-glycosides with Distinct Effects on Accumulation of Other Flavonoid Metabolites in Pears (Pyrus bretschneideri Rehd.). PLoS ONE 2014, 9, e91945. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.J.; Fang, X.; Li, C.Y.; Yang, L.; Chen, X.Y. General and specialized tyrosine metabolism pathways in plants. aBIOTECH 2019, 1, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Xiong, H.; Li, N.; Song, J.; Zheng, Y.L.; Liu, X.Z.; Zhao, P. De novo Transcriptome Sequencing of Vaccinium dunalianum Wight to Investigate Arbutin and 6′-O-Caffeoylarbutin Synthesis. Russ. J. Plant Physiol. 2017, 64, 260–282. [Google Scholar] [CrossRef]
- Islam, M.Z.; Hu, X.M.; Jin, L.F.; Liu, Y.Z.; Peng, S.A. Genome-wide identification and expression profile analysis of citrus sucrose synthase genes: Investigation of possible roles in the regulation of sugar accumulation. PLoS ONE 2014, 9, e113623. [Google Scholar] [CrossRef]
- Xu, Z.S.; Lin, Y.Q.; Xu, J.; Zhu, B.; Zhao, W.; Peng, R.H.; Yao, Q.H. Selective Detoxification of Phenols by Pichia pastoris and Arabidopsis thaliana Heterologously Expressing the PtUGT72B1 from Populus trichocarpa. PLoS ONE 2013, 8, e66878. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Ding, Y.; Zhu, D.Y.; Yin, J.T.; Liu, X.Z.; Zhao, P. Cloning and Bioinformatics Analysis of VdAS1 Gene in Vaccinium dunalianum (Ericaceae). Plant Divers. Resour. 2015, 37, 71–77. [Google Scholar]
- Li, J.H.; Liu, S.H.; Cui, T.T.; Ding, B.P.; Zhou, Z.Q.; Li, L.L. Identification and Expression Characteristics of UDP-Glycosyltransferase Genes in Pear and Their Correlation with Arbutin Accumulation. Russ. J. Plant Physiol. 2022, 69, 82. [Google Scholar] [CrossRef]
- Wu, B.P.; Cao, X.M.; Liu, H.J.; Zhu, C.Q.; Klee, H.; Zhang, B.; Chen, K.S. UDP-glucosyltransferase PpUGT85A2 controls volatile glycosylation in peach. J. Exp. Bot. 2019, 70, 925–936. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Ma, Y.; Cui, T.; Liu, S.; Li, L. Transcriptome Profiling Reveals Potential Genes Involved in Salicylic Acid-Induced Arbutin Synthesis in Pear. Horticulturae 2024, 10, 622. https://doi.org/10.3390/horticulturae10060622
Li J, Ma Y, Cui T, Liu S, Li L. Transcriptome Profiling Reveals Potential Genes Involved in Salicylic Acid-Induced Arbutin Synthesis in Pear. Horticulturae. 2024; 10(6):622. https://doi.org/10.3390/horticulturae10060622
Chicago/Turabian StyleLi, Junhao, Yuchen Ma, Tingting Cui, Shaohua Liu, and Liulin Li. 2024. "Transcriptome Profiling Reveals Potential Genes Involved in Salicylic Acid-Induced Arbutin Synthesis in Pear" Horticulturae 10, no. 6: 622. https://doi.org/10.3390/horticulturae10060622
APA StyleLi, J., Ma, Y., Cui, T., Liu, S., & Li, L. (2024). Transcriptome Profiling Reveals Potential Genes Involved in Salicylic Acid-Induced Arbutin Synthesis in Pear. Horticulturae, 10(6), 622. https://doi.org/10.3390/horticulturae10060622