
Iris typhifolia Responses to Saline–Alkali Stress: Germination, Antioxidant Activity, Hormones, and Photosynthetic Performance

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Method
2.2.1. Experimental Design
Artificial Salinity−Alkalinity Conditions
Seeds Treatment
Seedlings Treatment
2.2.2. Determination Method
Seed Germination Parameters
Physiological Parameters
Chlorophyll Concentration
Photosynthetic Parameters
Chlorophyll Fluorescence Parameters
2.2.3. Statistical Analysis
3. Results
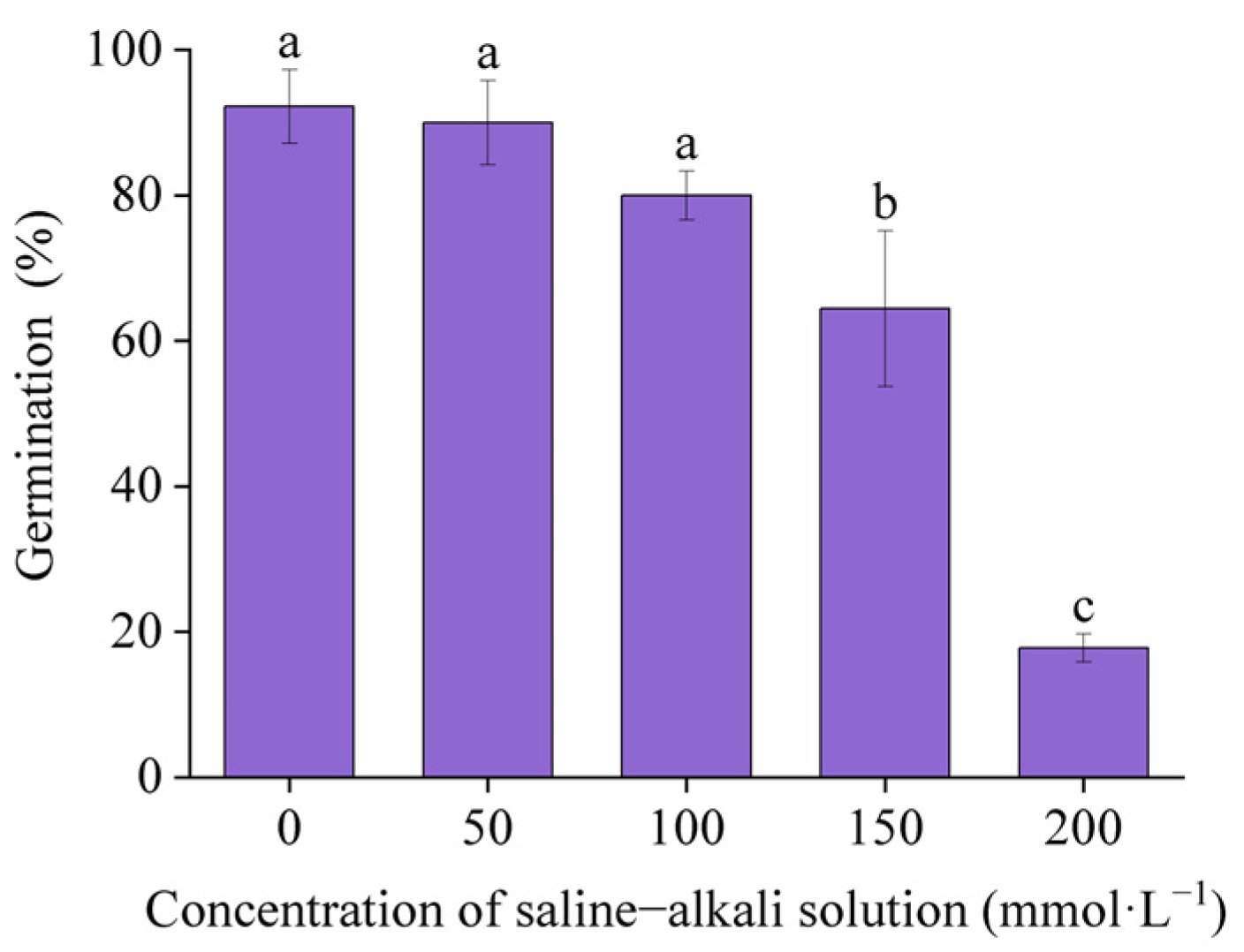
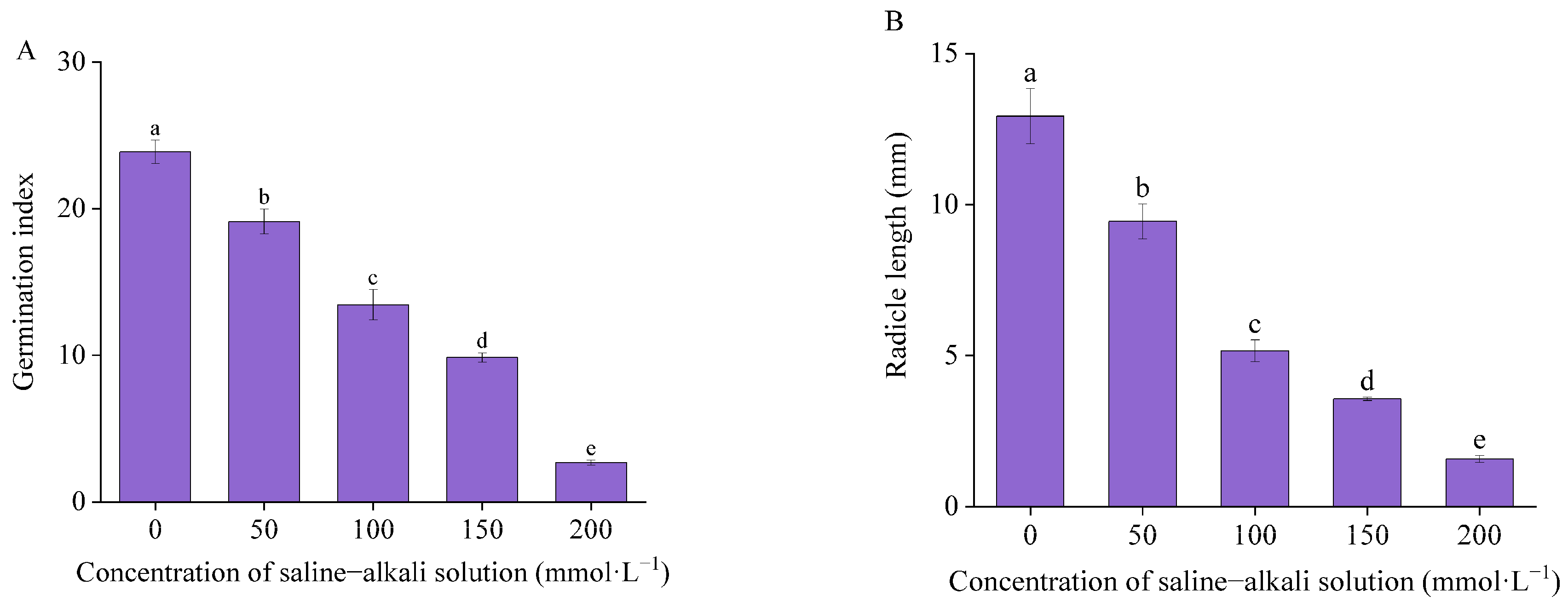
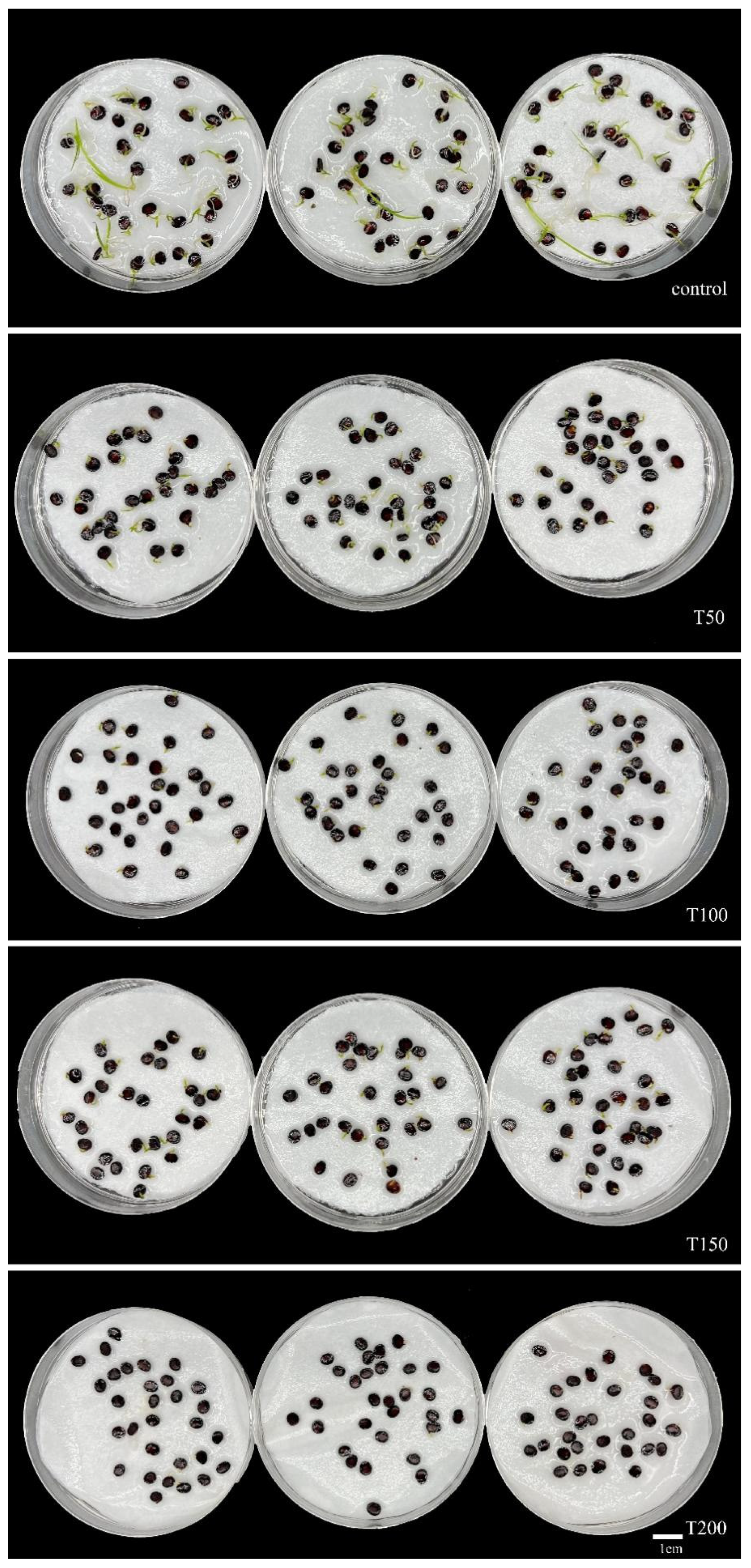
3.1. Seed Germination
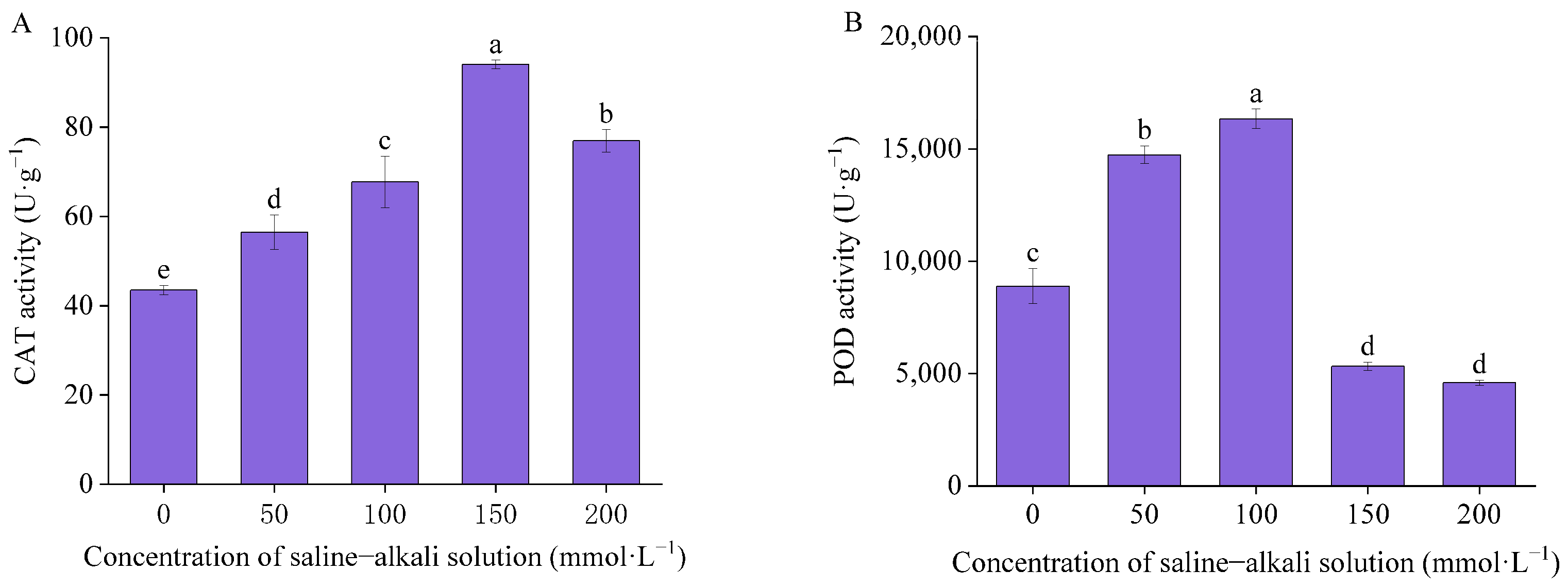
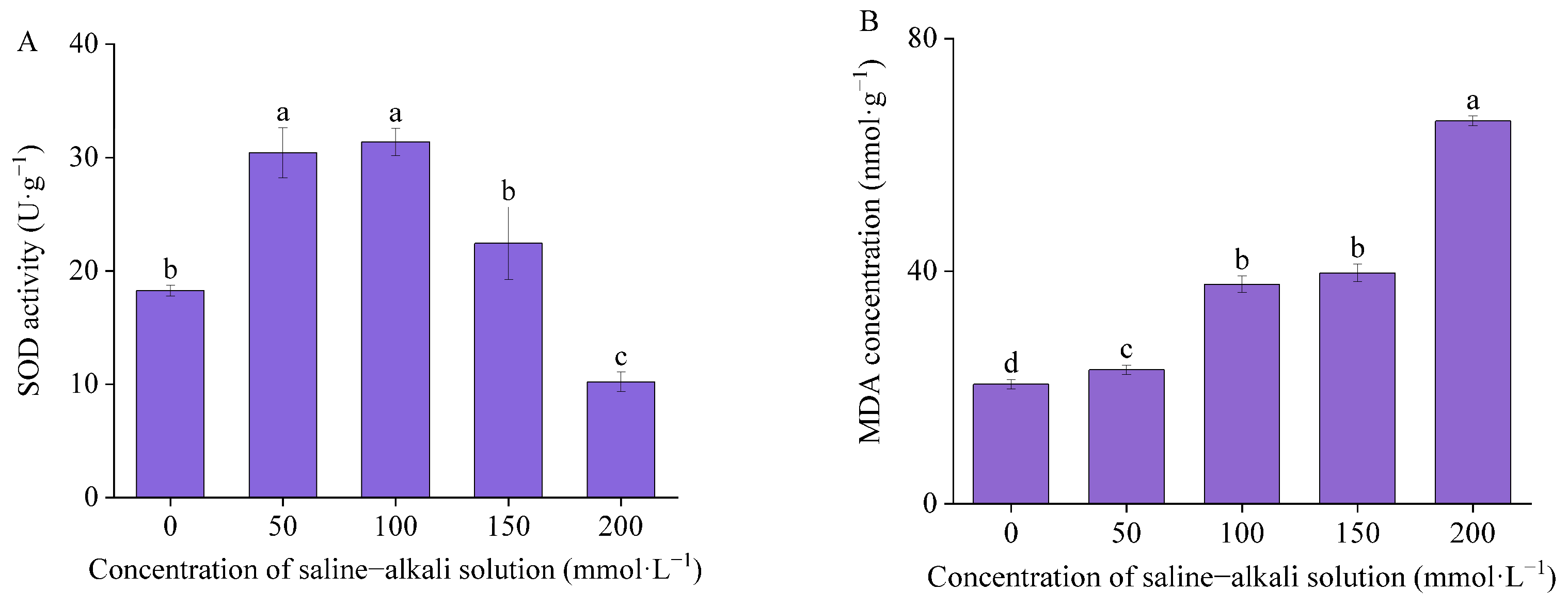
3.2. Antioxidant Enzyme Activity and Malondialdehyde Concentration
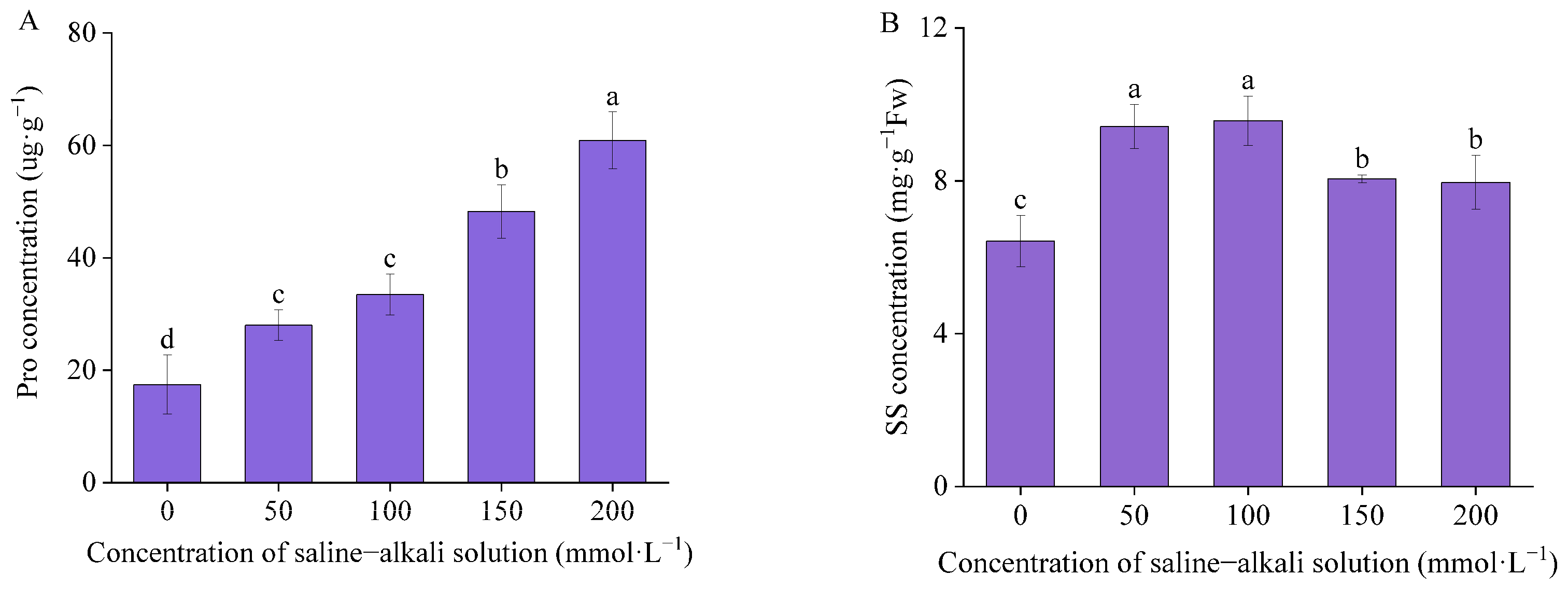
3.3. Osmotic Adjustment Substances
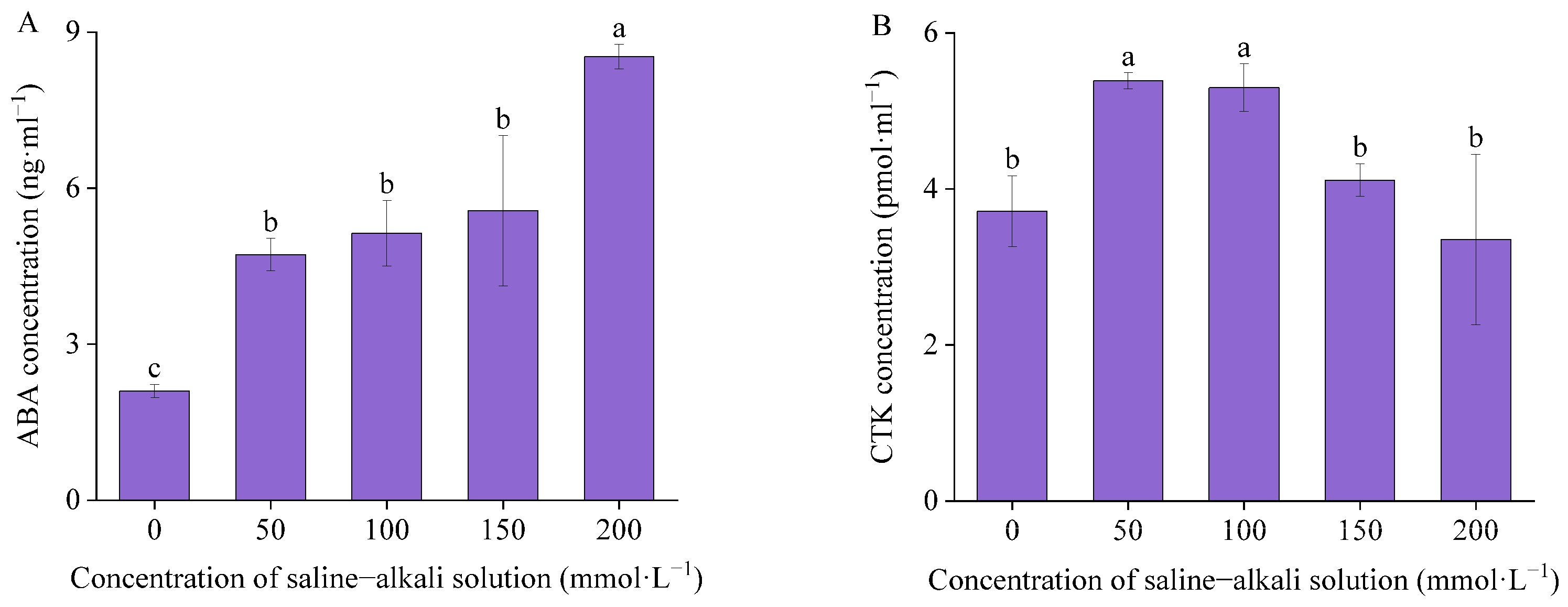
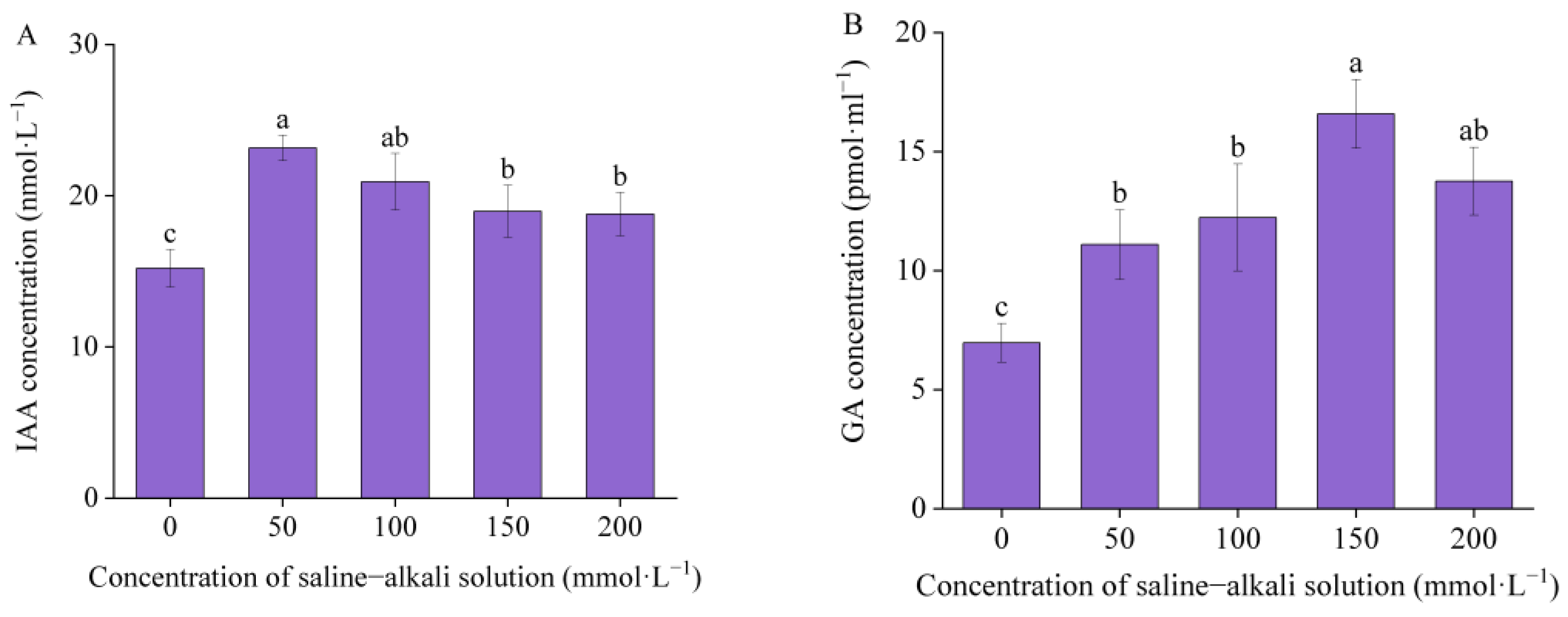
3.4. Endogenous Hormones Concentration
3.5. Determination of Threshold Value of Seed Germination Period
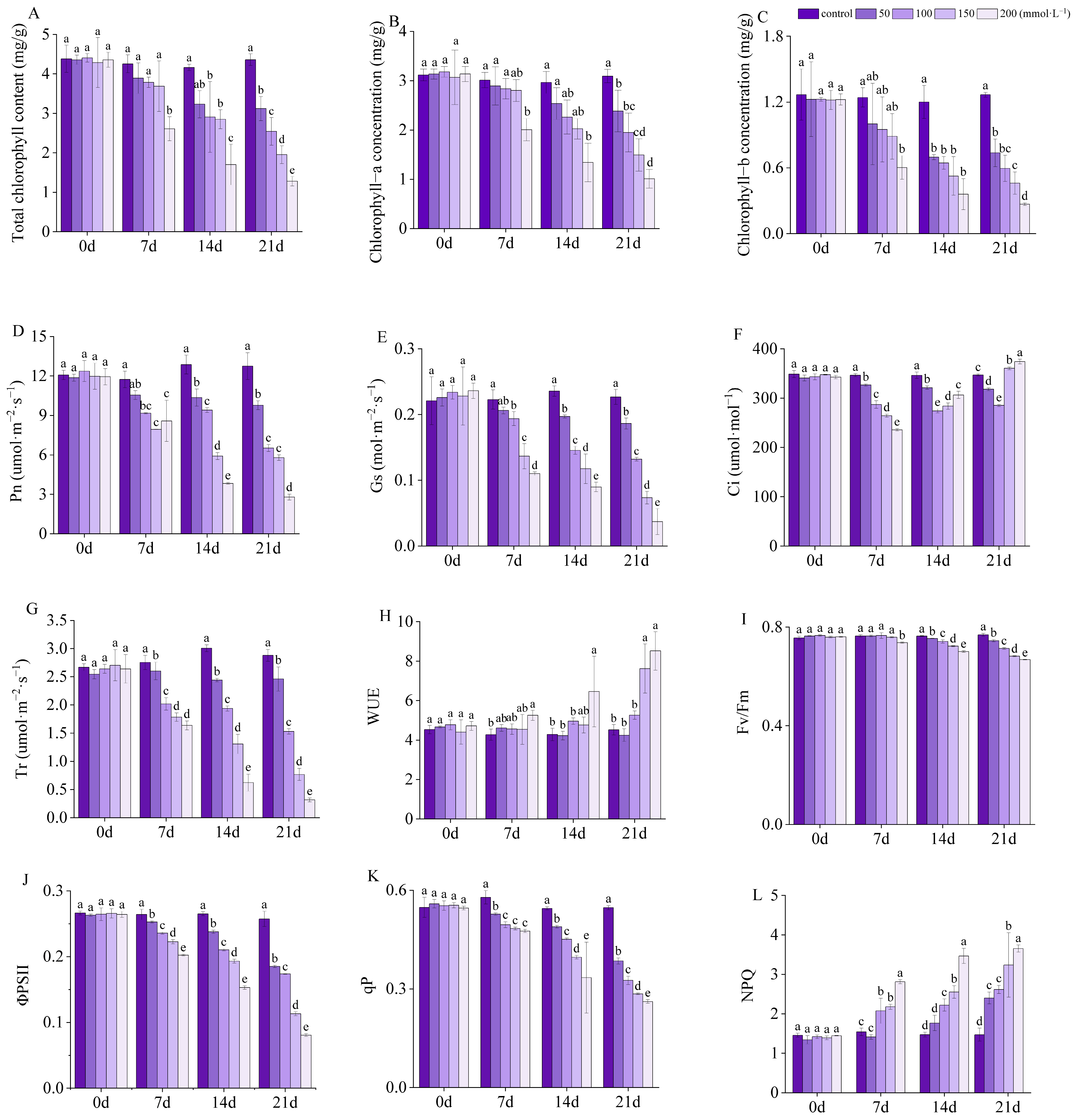
3.6. Chlorophyll Concentration
3.7. Photosynthetic Parameters
3.8. Chlorophyll Fluorescence Parameters
4. Discussion
4.1. Effects of Artificial Saline–Alkali Stress on Seed Germination of I. typhifolia
4.2. Effects of Artificial Saline–Alkali Stress on Antioxidant Enzyme Activity and Malondialdehyde Concentration in Germinated Seeds of I. typhifolia
4.3. Effects of Artificial Saline–Alkali Stress on the Concentration of Osmotic Adjustment Substances in the Germinated Seeds of I. typhifolia
4.4. Effects of Artificial Saline–Alkali Stress on Endogenous Hormones in I. typhifolia Germinated Seeds
4.5. Effects of Artificial Saline–Alkali Stress on Chlorophyll and Photosynthetic Parameters of I. typhifolia Seedlings
4.6. Effects of Artificial Saline–Alkali Stress on Chlorophyll Fluorescence Parameters of I. typhifolia Seedlings
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bai, J.; Li, Y.; Zhang, J.; Xu, F.; Bo, Q.; Wang, Z.; Yue, S. Straw returning and one-time application of a mixture of controlled release and solid granular urea to reduce carbon footprint of plastic film mulching spring maize. J. Clean. Prod. 2021, 280, 124478. [Google Scholar] [CrossRef]
- Tao, C.S.; Yao, L.J.; Chen, Y.C.; Li, L.Y.; Kong, Y.P. The impact of expressway snowmelt agent usage on the environment in an extreme freezing snow and sleet condition. IOP Conf. Ser. Earth Environ. Sci. 2018, 191, 012073. [Google Scholar] [CrossRef]
- Green, S.M.; Machin, R.; Cresser, M.S. Effect of long-term changes in soil chemistry induced by road salt applications on n-transformations in roadside soils. Environ. Pollut. 2008, 152, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Li, H.; Huang, S.L.; Ren, C.H.; Weng, X.H.; Zhang, S.Z.; Liu, L.Y.; Pei, J.B. Optimal exogenous calcium alleviates the damage of Snow-melting agent to Salix matsudana seedlings. Front. Plant Sci. 2022, 13, 928092. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.P.; Chong, P.F.; Zhao, M. Effect of salt stress on the photosynthetic characteristics and endogenous hormones, and: A comprehensive evaluation of salt tolerance in Reaumuria soongorica seedlings. Plant Signal. Behav. 2022, 17, 2031782. [Google Scholar] [CrossRef] [PubMed]
- Quan, X.Y.; Liang, X.L.; Li, H.M.; Xie, C.J.; He, W.X.; Qin, Y.X. Identification and characterization of wheat germplasm for salt tolerance. Plants 2021, 10, 268. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Rehman, A.; Li, X.W.; Jiang, X.; Tian, C.Y.; Wang, X.Y.; Li, H.G.; Wang, Z.Z.; He, S.P.; Du, X.M. Comprehensive evaluation and transcriptome analysis reveal the salt tolerance mechanism in semi-wild cotton (Gossypium purpurascens). Int. J. Mol. Sci. 2023, 24, 12853. [Google Scholar] [CrossRef]
- Muhammad, N.; Rabail, Z.; Shehzad, K.K.; Noman, Y.; Farhan, Q.M.; Martin, B.; Jiri, H.; Adnan, M. Unveiling the potential of acidified cow dung in combination with plant growth promoting endophytes on growth, physiology, and yield improvement of maize in salt-affected soil. Arab. J. Geosci. 2023, 16, 551. [Google Scholar] [CrossRef]
- Sasan, S.M.; Akbar, K.; Filippo, M. Photosynthesis and chlorophyll fluorescence of Iranian licorice (Glycyrrhiza glabra L.) accessions under salinity stress. Front. Plant Sci. 2022, 13, 984944. [Google Scholar] [CrossRef]
- Mann, A.; Kumar, N.; Lata, C.; Kumar, A.; Meena, B.L.; Kumar, A. Physiological and differential gene expression reveals a trade-off between antioxidant capacity and salt tolerance in Urochondra setulosa and Dichanthium annulatum. Plant Growth Regul. 2024, 102, 555–570. [Google Scholar] [CrossRef]
- Lu, X.; Ma, L.; Zhang, C.C.; Yan, H.K.; Bao, Y.; Gong, M.S.; Wang, W.H.; Li, S.; Ma, S.Y.; Chen, B.H. Grapevine (Vitis vinifera) responses to salt stress and alkali stress: Transcriptional and metabolic profiling. BMC Plant Biol. 2022, 22, 528. [Google Scholar] [CrossRef] [PubMed]
- Zang, W.; Miao, R.Q.; Zhang, Y.; Yuan, Y.; Pang, Q.Y.; Zhou, Z.Q. Metabolic and molecular basis for the salt and alkali responses of Suaeda corniculata. Environ. Exp. Bot. 2021, 192, 104643. [Google Scholar] [CrossRef]
- Chen, Z.; Niu, J.P.; Guo, Z.P.; Sui, X.; Xu, N.; Kareem, H.A.; Hassan, M.U.; Yan, M.; Zhang, Q.; Cui, J.; et al. Graphene enhances photosynthesis and the antioxidative defense system and alleviates salinity and alka-linity stresses in alfalfa (Medicago sativa L.) by regulating gene expression. Environ. Sci.-Nano 2021, 8, 2731–2748. [Google Scholar] [CrossRef]
- Kha, W.; Prithivira, B.; Smit, D.L. Photosynthetic responses of corn and soybean to foliar application of salicylates. J. Plant Physiol. 2003, 160, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Afzal, I.; Basra, S.M.A.; Cheema, M.A.; Farooq, M.; Jafar, M.Z.; Shahid, M.; Yasmeen, A. Seed priming: A shotgun approach for alleviation of salt stress in wheat. Int. J. Agric. Biol. 2013, 15, 1199–1203. [Google Scholar] [CrossRef]
- Pereira, I.C.; Catão, H.C.R.M.; Caixeta, F. Seed physiological quality and seedling growth of pea (Pisum sativum) under water and salt stress. Rev. Bras. Eng. Agríc. Ambient. 2020, 24, 95–100. [Google Scholar] [CrossRef]
- Chen, L.; Meng, Y.; Jiang, D.; Yang, F.; Zhou, Y. Physio–biochemical responses of three Aquilegia species seedlings to salt stress. Agronomy 2022, 12, 2841. [Google Scholar] [CrossRef]
- Fang, S.; Hou, X.; Liang, X. Response mechanisms of plants under saline–alkali stress. Front. Plant Sci. 2021, 12, 667458. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef]
- Li, J.; Xu, H.; Liu, W.; Zhang, X.; Lu, Y. Ethylene inhibits root elongation during alkaline stress through AUXIN1 and associated changes in auxin accumulation. Plant Physiol. 2015, 168, 1777–1791. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yang, A.; Zhang, W. Higher endogenous bioactive gibberellins and α-amylase activity confer greater tolerance of rice seed germination to saline-alkaline stress. Environ. Exp. Bot. 2019, 162, 357–363. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Niu, C.; Yu, Q.; Wang, Q.; Wang, J.; Sun, X.; Wang, Z.; Shan, X. Experimental investigation of the erodibility of soda saline-alkali soil under freeze-thaw cycle from a microscopic view. Catena 2023, 232, 107430. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, R.; Zhu, H.; Cheng, X.; Shutes, B.; Yan, B. Seed germination and early seedling growth of six wetland plant species in saline-alkaline environment. Int. J. Phytoremediation 2020, 22, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Tian, K.; Ban, Z.; Xu, H.; Jia, W.; Zhu, Y.; Chen, H. Analysis of floral fragrance components in different parts of Iris typhifolia. Horticulturae 2023, 9, 1268. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, T.; Cheng, Y.; Wang, F.; Zhao, X. Morphological and metabolic responses of four Iris germanica cultivars under salinity stress. Sci. Hortic. 2021, 281, 109960. [Google Scholar] [CrossRef]
- Yu, X.; Liu, Y.; Cao, P.; Zeng, X.; Xu, B.; Luo, f.; Yang, X. Morphological structure and physiological and biochemical responses to drought stress of Iris japonica. Plants 2023, 12, 3729. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, D.; Zhao, X.; Zhang, M. Evaluation of drought resistance and transcriptome analysis for the identification of drought-responsive genes in Iris germanica. Sci. Rep. 2021, 11, 16308. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Z.; Liu, Q.; Tang, J.; Huang, S.; Dhankher, O.P.; Yuan, H. Growth, physiological adaptation, and NHX gene expression analysis of Iris halophila under salt stress. Environ. Sci. Pollut. Res. 2018, 25, 207–25216. [Google Scholar] [CrossRef] [PubMed]
- Alamgir, M.; Hossain, M.K. Effect of pre-sowing treatments on germination and initial seedling development of Albizia saman in the nursery. J. For. Res. 2005, 16, 200–204. [Google Scholar] [CrossRef]
- Holloway, P.S. Seed germination of Alaska Ifs. Hortscience 1987, 22, 898–899. [Google Scholar] [CrossRef]
- Ryuya, M.; Katsumasa, Y.; Daisuke, H.; Yasuhisa, H. Effects of salinity, temperature, and immersion conditions on seed germination of invasive Spartina alterniflora Loisel (Smooth cordgrass) in Japan. Reg. Stud. Mar. Sci. 2023, 57, 102738. [Google Scholar] [CrossRef]
- Önen, H.; Farooq, S.; Tad, S.; Özaslan, C.; Gunal, H.; Chauhan, B.S. The influence of environmental factors on germination of burcucumber (Sicyos angulatus) seeds: Implications for range expansion and management. Weed Sci. 2018, 66, 494–501. [Google Scholar] [CrossRef]
- Jianting, L.; Mengjie, D.; Chuanqi, W.; Yanjun, M. Effects of drought and salt stress on seed germination and seedling growth of Elymus nutans. PeerJ 2023, 11, e15968. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Dong, H.; Deng, Z. Physiological mechanisms of Bretschneidera sinensis hemsl. seed dormancy release and germination. Forests 2023, 14, 2430. [Google Scholar] [CrossRef]
- Wang, X.D.; Shen, H.L.; Yang, L. The Response of hormones, reactive oxygen species and nitric oxide in the Polyethylene-Glycol-Promoted, salt–alkali-stress-induced embryo germination of Sorbus pohuashanensis. Int. J. Mol. Sci. 2024, 25, 5128. [Google Scholar] [CrossRef]
- Sifiso, N.; Oluwaseyi, S.B.; Magnus, A.U.; Ida, R. Optimal chlorophyll extraction conditions and postharvest stability in Moringa (M. Oleifera) leaves. J. Food Meas. Charact. 2023, 18, 1611–1626. [Google Scholar] [CrossRef]
- Kochaphan, V.; Piyada, T.; Supranee, S.; Watanachai, L.; Anoma, D. Photosynthesis performance at different growth stages, growth, and yield of rice in saline fields. Plants 2023, 12, 1903. [Google Scholar] [CrossRef]
- Nirmalendu, B.; Kumar, A.R.; Parul, S. Assessing soil quality for rehabilitation of salt-affected agroecosystem: A comprehensive review. Front. Environ. Sci. 2022, 10, 935785. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Stoleru, V.; Slabu, C.; Vitanescu, M.; Peres, C.; Cojocaru, A.; Covasa, M.; Mihalache, G. Tolerance of Three quinoa cultivars (Chenopodium quinoa Willd.) to salinity and alkalinity stress during germination stage. Agronomy 2019, 9, 287. [Google Scholar] [CrossRef]
- Javaid, M.M.; Mahmood, A.; Alshaya, D.S.; Alkahtani, M.D.F.; Waheed, H.; Wasaya, A.; Khan, S.A.; Naqve, M.; Haider, I.; Shahid, M.A.; et al. Influence of environmental factors on seed germination and seedling characteristics of perennial ryegrass (Lolium perenne L.). Sci. Rep. 2022, 12, 9522. [Google Scholar] [CrossRef]
- He, L.; Zhao, Y.Y.; Lu, Z.H. Effects of saline-alkaline stress on seed germination and seedling growth of Sorghum bicolor (L.) Moench. Appl. Biochem. Biotechnol. 2014, 173, 1680–1691. [Google Scholar] [CrossRef]
- Shah, F.A.; Wei, X.; Wang, Q.J.; Liu, W.B.; Wang, D.D.; Yao, Y.Y.; Hu, H.; Chen, X.; Huang, S.; Hou, J.; et al. Karrikin improves osmotic and salt stress tolerance via the regulation of the redox homeostasis in the oil plant Sapium sebiferum. Front. Plant Sci. 2020, 11, 216. [Google Scholar] [CrossRef]
- Chen, H.; Shi, J.; Tao, L.; Han, X.; Lin, G.; Chen, X. Exogenous spermidine priming mitigates the osmotic damage in germinating seeds of Leymus chinensis under salt–alkali stress. Front. Plant Sci. 2021, 12, 701538. [Google Scholar] [CrossRef]
- Baily, C. The signaling role of ROS in the regulation of seed germination and dormancy. Biochem. J. 2019, 476, 3019–3032. [Google Scholar] [CrossRef]
- Chen, H.; Tao, L.; Shi, J.; Han, X.; Cheng, X. Exogenous salicylic acid signal reveals an osmotic regulatory role in priming the seed germination of Leymus chinensis under salt–alkali stress. Environ. Exp. Bot. 2021, 188, 104498. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, W.; Ren, C.; Zhan, C.; Wang, C.; Li, J.; Ren, Q.; Liang, X.; Wei, L.; Xiang, D.; et al. Physiological and biochemical mechanisms of exogenous melatonin regulation of saline–alkali tolerance in oats. Agronomy 2023, 13, 1327. [Google Scholar] [CrossRef]
- Wang, Y.; Jie, W.; Peng, X.; Hua, X.; Yan, X.; Zhou, Z.; Lin, J. Physiological adaptive strategies of oil seed crop Ricinus communis early seedlings (Cotyledon vs. True Leaf) under salt and alkali stresses: From the growth, photosynthesis and chlorophyll fluorescence. Front. Plant Sci. 2019, 9, 01939. [Google Scholar] [CrossRef]
- Shen, H.L.; Wang, Y.T.; Wang, X.D.; Yang, L.Z.; Cai, H. Exogenous ethylene alleviates the inhibition of Sorbus pohuashanensis embryo germination in a saline-alkali environment (NaHCO3). Int. J. Mol. Sci. 2023, 24, 4244. [Google Scholar] [CrossRef]
- Pavla, V.; Helena, H.; Frantisek, H.; Kamil, K. Salinity stress affects photosynthesis, malondialdehyde formation, and proline content in Portulaca Oleracea L. Plants 2021, 10, 845. [Google Scholar] [CrossRef]
- Shu, Y.J.; An, Y.M.; Guo, C.H.; Liu, Y.R.; Song, L.L. De novo transcriptional analysis of alfalfa in response to saline-alkaline stress. Front. Plant Sci. 2016, 7, 931. [Google Scholar] [CrossRef]
- Feng, Z.H.; Lu, G.R.; Sun, M.; Jin, Y.Y.; Xu, Y.; Liu, X.L.; Wang, M.M.; Liu, M.; Yang, H.Y.; Guan, Y.; et al. Comparative study of the priming effect of abscisic acid on tolerance to saline and alkaline stresses in rice seedlings. Agronomy 2023, 13, 2698. [Google Scholar] [CrossRef]
- Hou, R.; Yang, L.; Wuyun, T.; Chen, S.; Zhang, L. Genes related to osmoregulation and antioxidation play important roles in the response of Trollius chinensis seedlings to saline alkali stress. Front. Plant Sci. 2023, 14, 1080504. [Google Scholar] [CrossRef]
- Fatma, M.; Iqbal, M.; Khan, R.; Masood, A.; Khan, N.A. Coordinate changes in assimilatory sulfate reduction are correlated to salt tolerance: Involvement of phytohormones. Annu. Rev. Res. Biol. 2013, 127, 543. [Google Scholar]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signaling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Llanes, A.; Andrade, A.; Masciarelli, O.; Alemano, S.; Luna, V. Drought and salinity alter endogenous hormonal profiles at the seed germination phase. Seed Sci. Res. 2016, 26, 1–13. [Google Scholar] [CrossRef]
- Nasri, N.; Mahmoudi, H.; Baatour, O.; M’rah, S.; Kaddour, R.; Lachâal, M. Effect of exogenous gibberellic acid on germination, seedling growth and phosphatase activities in Lettuce under salt stress. Afr. J. Biotechnol. 2012, 11, 11967–11971. [Google Scholar] [CrossRef]
- Jamil, M.; Rha, E.S. Gibberellic acid (GA3) enhance seed water uptake, germination and early seedling growth in sugar beet under salt stress. Pak. J. Biol. Sci. 2007, 10, 654–658. [Google Scholar] [CrossRef]
- Sano, N.; Marion-Poll, A. ABA metabolism and homeostasis in seed dormancy and germination. Int. J. Mol. Sci. 2021, 22, 5069. [Google Scholar] [CrossRef]
- Lu, C.; Li, L.; Liu, X.; Chen, M.; Wan, S.; Li, G. Salt stress inhibits photosynthesis and destroys chloroplast structure by downregulating chloroplast development–related genes in Robinia pseudoacacia seedlings. Plants 2023, 12, 1283. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Li, X.; He, G.; Che, Y.; Teng, Z.; Shao, J.; Xu, N.; Sun, G. Chlorophyll synthesis and the photoprotective mechanism in leaves of mulberry (Morus alba L.) seedlings under NaCl and NaHCO3 stress revealed by TMT–based proteomics analyses. Ecotoxicol. Environ. Safe 2020, 190, 110164. [Google Scholar] [CrossRef]
- Sarvandi, S.; Nia, A.E.; Nejad, A.R.; Azimi, M.H. Morpho–physiological responses of some Iris cultivars under drought and salinity stresses. J. Agric. Sci. Technol. 2020, 22, 535–546. [Google Scholar]
- Li, C.; Mur, L.A.J.; Wang, Q.; Hou, X.; Zhao, C.; Chen, Z.; Wu, J.; Guo, Q. ROS scavenging and ion homeostasis is required for the adaptation of halophyte Karelinia caspia to high salinity. Front. Plant Sci. 2022, 13, 979956. [Google Scholar] [CrossRef]
- Müller, B.; Arcoverde Cerveira Sterner, V.; Papp, L.; May, Z.; Orlóci, L.; Gyuricza, C.; Sági, L.; Solti, Á.; Fodor, F. Alkaline salt tolerance of the biomass plant Arundo donax. Agronomy 2022, 12, 1589. [Google Scholar] [CrossRef]
- Ma, Q.; Wu, C.; Liang, S.; Yuan, Y.; Liu, C.; Liu, J.; Feng, B. The alkali tolerance of broomcorn millet (Panicum miliaceum L.) at the germination and seedling stage: The case of 296 broomcorn millet genotypes. Front. Plant Sci. 2021, 12, 711429. [Google Scholar] [CrossRef]
- Song, X.; Zhou, G.; He, Q.; Zhou, H. Stomatal limitations to photosynthesis and their critical water conditions in different growth stages of maize under water stress. Agric. Water Manag. 2020, 241, 106330. [Google Scholar] [CrossRef]
- Shi, C.; Yang, F.; Liu, Z.; Li, Y.; Di, X.; Wang, J.; Lin, J. Uniform water potential induced by salt, alkali, and drought stresses has different impacts on the seedling of Hordeum jubatum: From growth, photosynthesis, and chlorophyll fluorescence. Front. Plant Sci. 2021, 12, 733236. [Google Scholar] [CrossRef]
- Guidi, L.; Piccolo, E.L.; Landi, M. Chlorophyll fluorescence, photoinhibition and abiotic stress: Does it make any difference the fact to be a C3 or C4 species? Front. Plant Sci. 2019, 10, 00174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, H.; Jiang, Y.; Li, H.; Zhang, Z.; Xu, Z.; Xu, B.; Huang, B. Natural variation of physiological traits, molecular markers, and chlorophyll catabolic genes associated with heat tolerance in perennial ryegrass accessions. BMC Plant Biol. 2020, 20, 520. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, M.; Gu, W.; Chen, Z.; Gu, Y.; Pei, L.; Tian, R. Effect of drought on photosynthesis, total antioxidant capacity, bioactive component accumulation, and the transcriptome of Atractylodes lancea. BMC Plant Biol. 2021, 21, 293. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | SSC | GR | GI | RL | CAT | POD | SOD | MDA | Pro | SS | ABA | CTK | IAA | GA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SSC | 1 | |||||||||||||
| GR | −0.901 * | 1 | ||||||||||||
| GI | −0.994 ** | 0.932 * | 1 | |||||||||||
| RL | −0.982 ** | 0.82 | 0.971 ** | 1 | ||||||||||
| CAT | 0.855 | −0.588 | −0.801 | −0.876 | 1 | |||||||||
| POD | −0.532 | 0.674 | 0.537 | 0.392 | −0.481 | 1 | ||||||||
| SOD | −0.432 | 0.738 | 0.489 | 0.277 | −0.148 | 0.875 | 1 | |||||||
| MDA | 0.939 * | −0.975 ** | −0.971 ** | −0.896 * | 0.639 | −0.547 | −0.611 | 1 | ||||||
| Pro | 0.99 ** | −0.934 * | −0.984 ** | −0.947 * | 0.825 | −0.613 | −0.521 | 0.941 * | 1 | |||||
| SS | 0.211 | 0.079 | −0.168 | −0.343 | 0.235 | 0.671 | 0.715 | 0.063 | 0.129 | 1 | ||||
| ABA | 0.944 * | −0.908 * | −0.948 * | −0.917 * | 0.681 | −0.386 | −0.389 | 0.933 * | 0.947 * | 0.339 | 1 | |||
| CTK | −0.341 | 0.615 | 0.39 | 0.193 | −0.163 | 0.914 * | 0.968 ** | −0.49 | −0.432 | 0.834 | −0.235 | 1 | ||
| IAA | 0.159 | 0.089 | −0.102 | −0.252 | 0.187 | 0.592 | 0.663 | 0.007 | 0.115 | 0.951 * | 0.329 | 0.802 | 1 | |
| GA | 0.851 | −0.563 | −0.789 | −0.881 * | 0.983 * | −0.371 | −0.046 | 0.619 | 0.82 | 0.385 | 0.724 | −0.029 | 0.359 | 1 |
| Index | Regression Equation | R2 | Threshold Value |
|---|---|---|---|
| Germination index | y = −0.1023x + 24.023 | 0.987 | 120.8 |
| Pro | y = −0.1436x + 16.1876 | 0.978 | 87.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Yu, J.; Lu, X.; Wang, Q.; Wang, S.; Shan, Y.; Liu, Y.; Meng, Y.; Zhou, Y. Iris typhifolia Responses to Saline–Alkali Stress: Germination, Antioxidant Activity, Hormones, and Photosynthetic Performance. Horticulturae 2024, 10, 588. https://doi.org/10.3390/horticulturae10060588
Chen L, Yu J, Lu X, Wang Q, Wang S, Shan Y, Liu Y, Meng Y, Zhou Y. Iris typhifolia Responses to Saline–Alkali Stress: Germination, Antioxidant Activity, Hormones, and Photosynthetic Performance. Horticulturae. 2024; 10(6):588. https://doi.org/10.3390/horticulturae10060588
Chicago/Turabian StyleChen, Lifei, Jiahui Yu, Xi Lu, Qi Wang, Shizhuo Wang, Yuze Shan, Yang Liu, Yuan Meng, and Yunwei Zhou. 2024. "Iris typhifolia Responses to Saline–Alkali Stress: Germination, Antioxidant Activity, Hormones, and Photosynthetic Performance" Horticulturae 10, no. 6: 588. https://doi.org/10.3390/horticulturae10060588
APA StyleChen, L., Yu, J., Lu, X., Wang, Q., Wang, S., Shan, Y., Liu, Y., Meng, Y., & Zhou, Y. (2024). Iris typhifolia Responses to Saline–Alkali Stress: Germination, Antioxidant Activity, Hormones, and Photosynthetic Performance. Horticulturae, 10(6), 588. https://doi.org/10.3390/horticulturae10060588