
Polyploidization Impact on Plant Architecture of Watermelon (Citrullus lanatus)

 , ,
, ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Location and Climatic Condition
2.3. Phenotypic Data Collection
2.4. Hormone Content Measurement
2.5. Exogenous Application of Plant Hormone
2.6. Histological Analysis
2.7. Statistical Analysis
3. Results
3.1. Comparison of Plant Architectural Traits among Diploid, Triploid, and Tetraploid Watermelon Plants
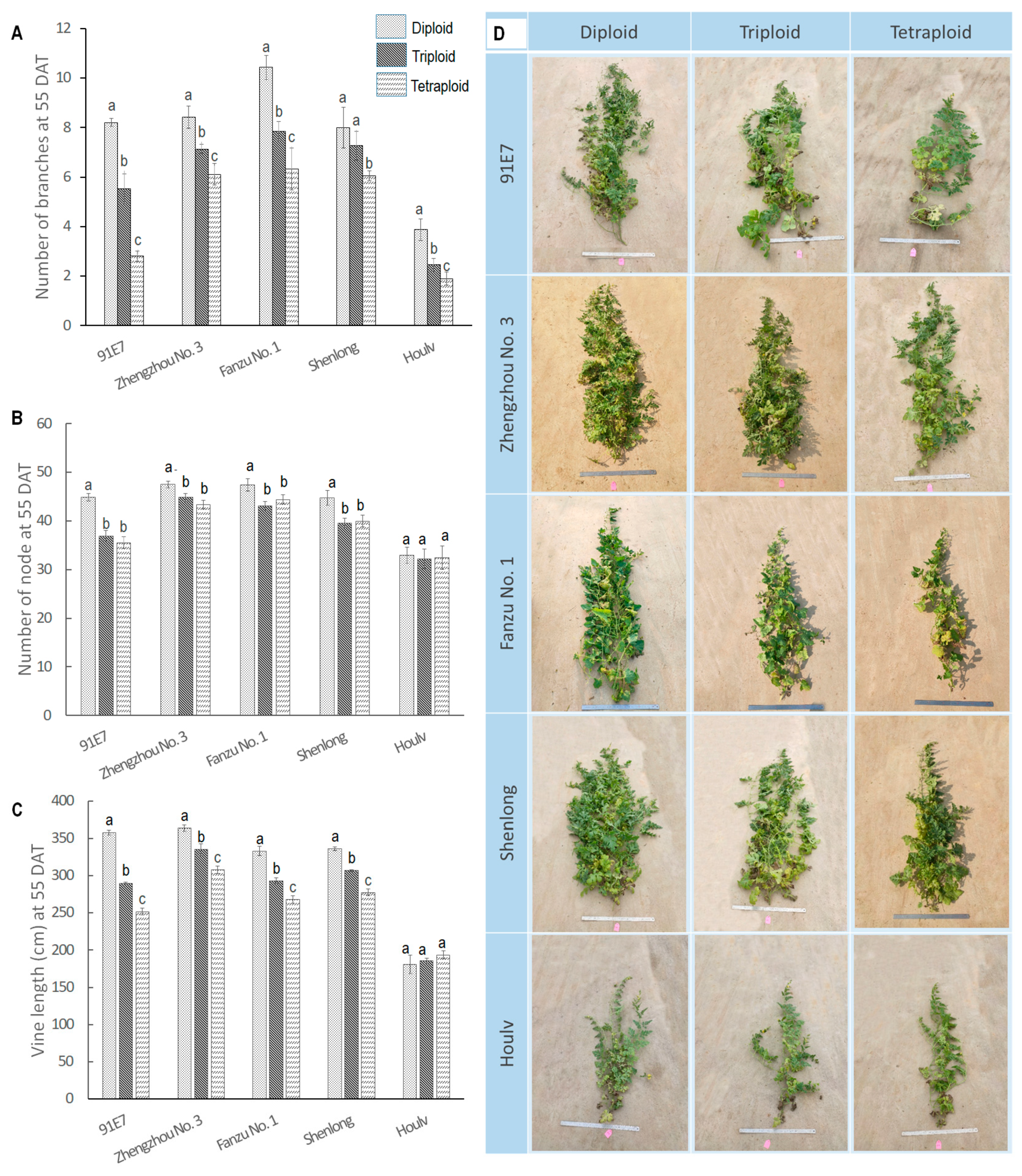
3.1.1. Ploidy Induced Variations in Branching
3.1.2. Ploidy Induced Variations in Node Number
3.1.3. Ploidy Induced Variations in Vine Length
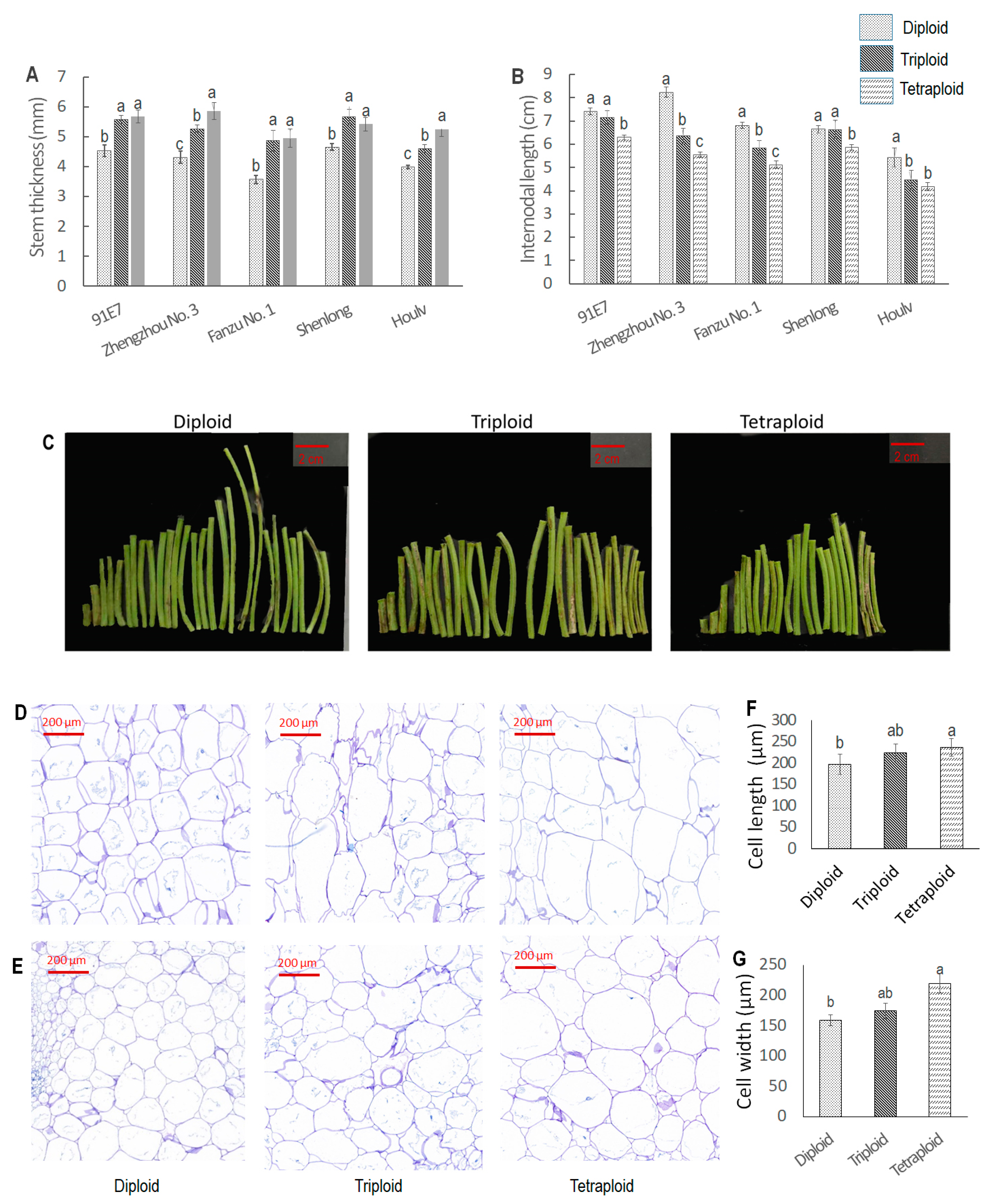
3.1.4. Ploidy-Induced Variations in Stem Thickness and Internodal Length
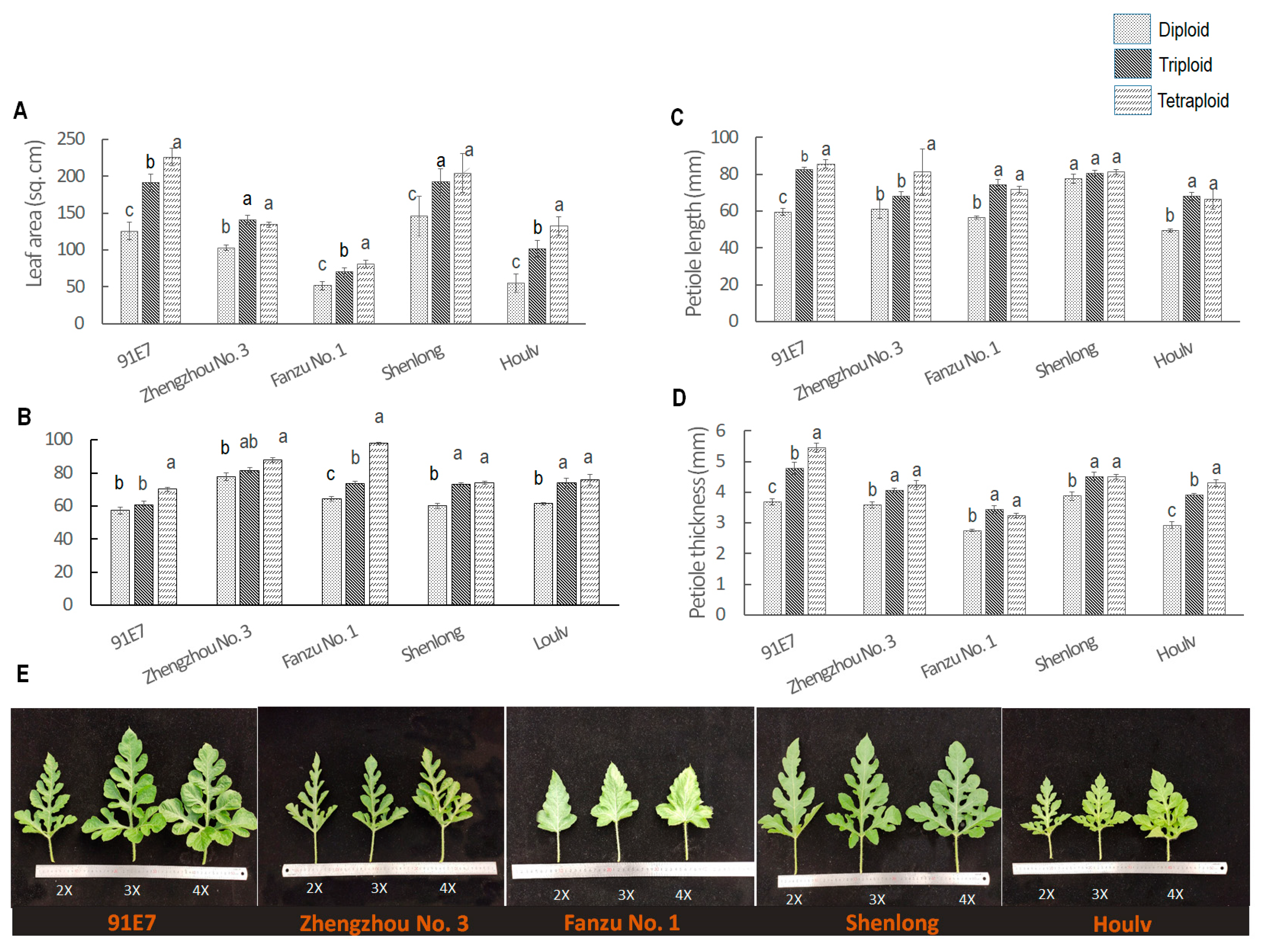
3.1.5. Ploidy-Induced Variations in Leaf Relate Traits
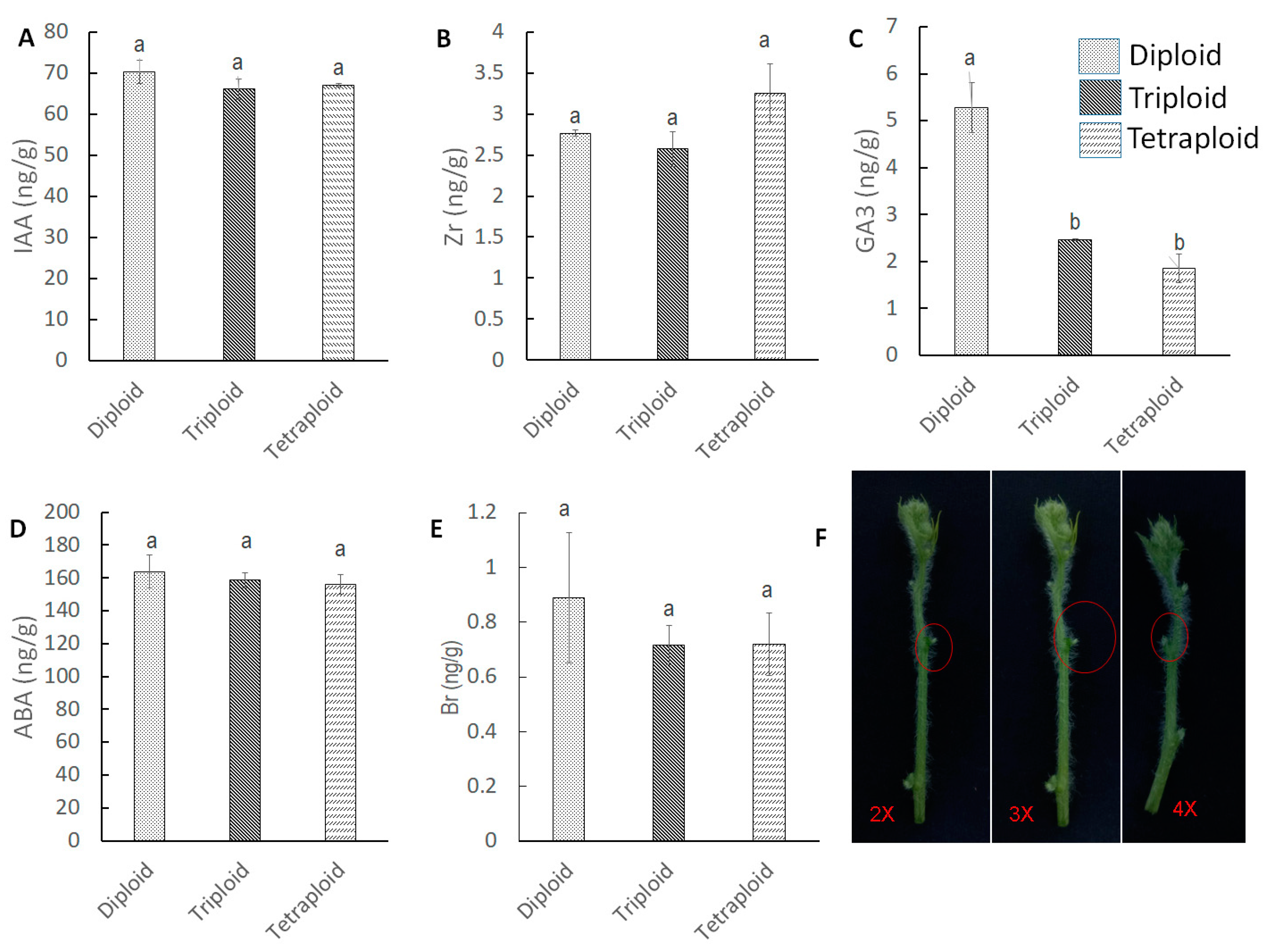
3.2. Quantification of Endogenous Hormones at Different Growth Stages in Different Tissue Parts
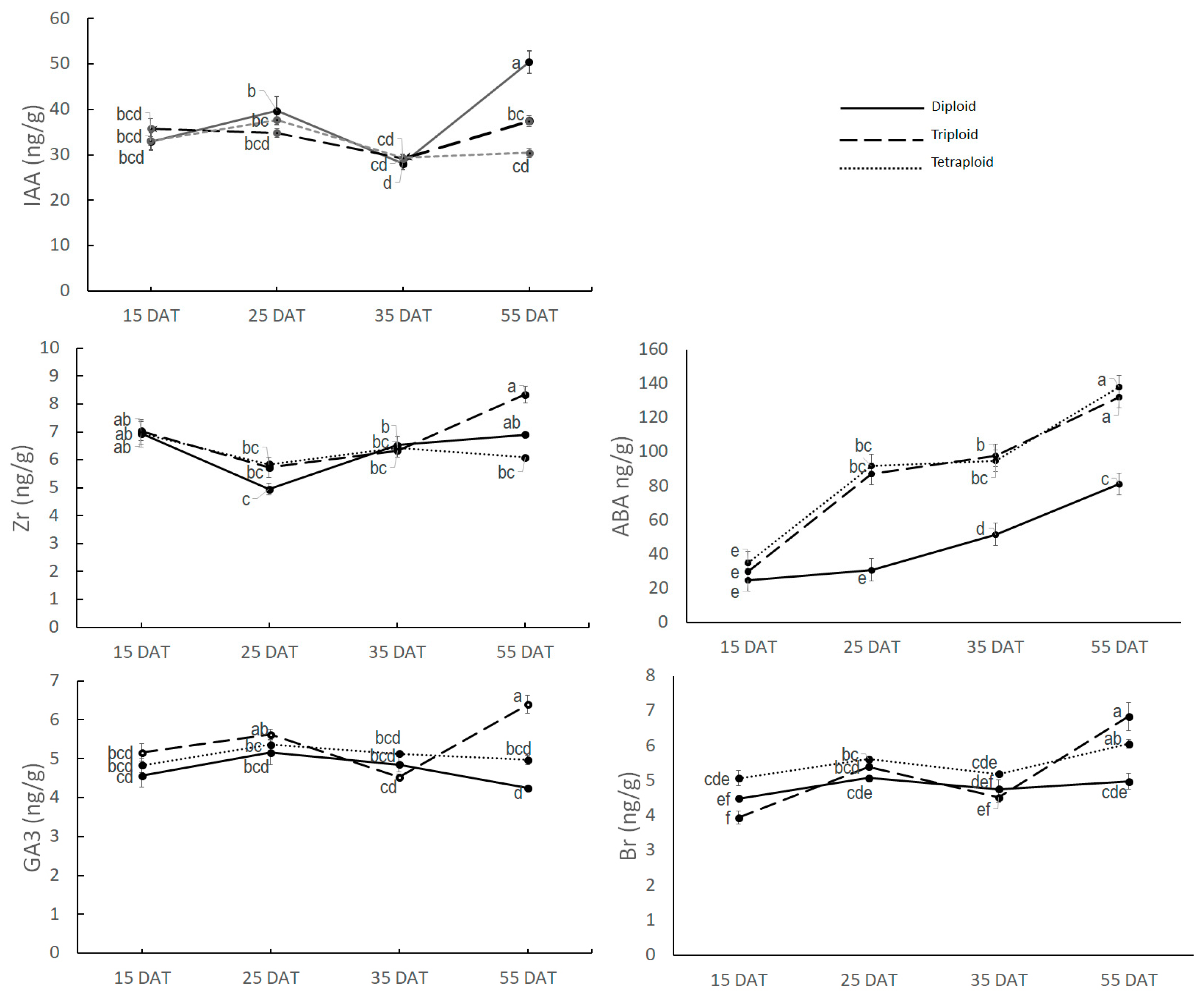
3.2.1. Hormone Quantification at Apical Meristem
3.2.2. Hormone Quantification at Nodal Tissue
3.2.3. Hormone Quantification at Auxiliary Bud
3.3. Correlation Study between the Morphological and Hormonal Traits
3.4. Principal Component Analysis of Morphological and Hormonal Traits
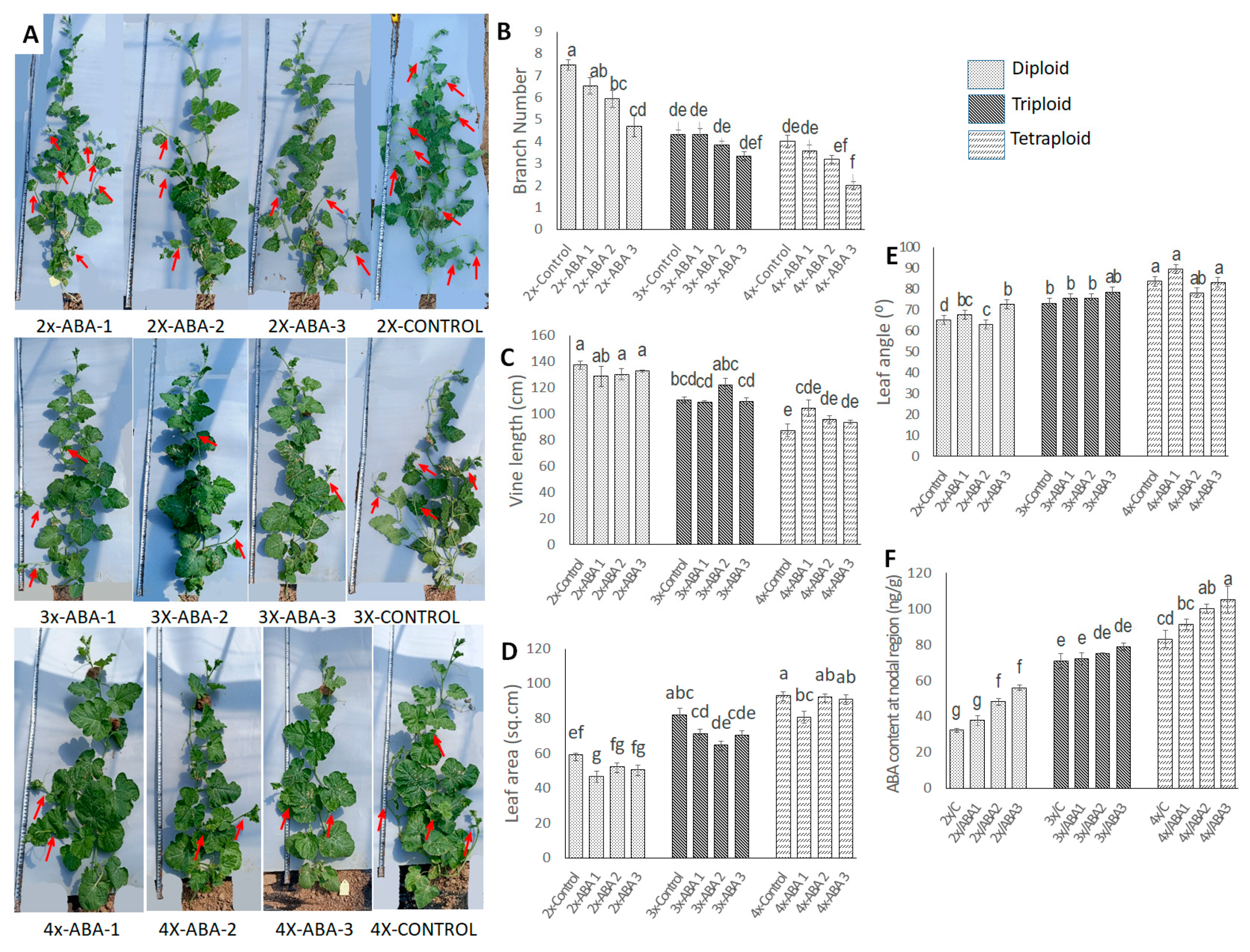
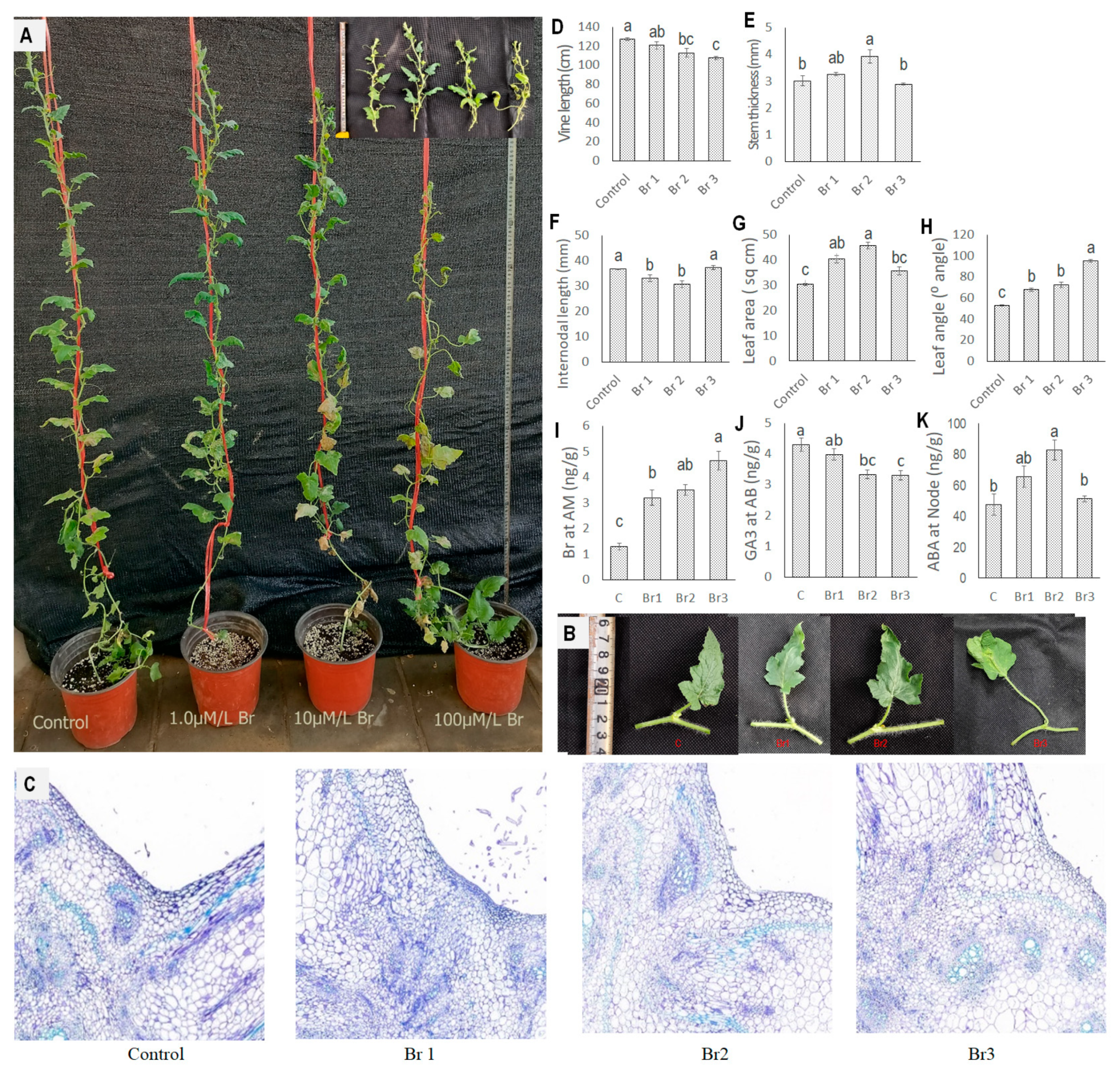
3.5. Effect of Exogenous Application of GA3, ABA, and Br on Plant Architectural Traits
3.5.1. Effect of Exogenous Application of GA3 on Plant Architectural Traits
3.5.2. Effect of Exogenous Application of ABA on Plant Architectural Traits
3.5.3. Effect of Exogenous Application of Br on Plant Architectural Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kihara, H. Triploid Watermelons. Proc. Am. Soc. Hortic. Sci. 1951, 58, 217–230. [Google Scholar]
- Smith, W.K.; Vogelmann, T.C.; DeLucia, E.H.; Bell, D.T.; Shepherd, K.A. Leaf Form and Photosynthesis. Bioscience 1997, 47, 785–793. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, J.; Qiu, B.; Ma, Z.; Lu, T.; Kang, X.; Yang, J. Induction and Characterization of Tetraploid Through Zygotic Chromosome Doubling in Eucalyptus Urophylla. Front. Plant Sci. 2022, 13, 870698. [Google Scholar] [CrossRef]
- Lynch, J.P. Harnessing Root Architecture to Address Global Challenges. Plant J. 2022, 109, 415–431. [Google Scholar] [CrossRef]
- Valladares, F.; Laanisto, L.; Niinemets, Ü.; Zavala, M.A. Shedding Light on Shade: Ecological Perspectives of Understorey Plant Life. Plant Ecol. Divers. 2016, 9, 237–251. [Google Scholar] [CrossRef]
- Mencuccini, M. Dwarf Trees, Super-Sized Shrubs and Scaling: Why Is Plant Stature so Important? Plant Cell Environ. 2015, 38, 1–3. [Google Scholar] [CrossRef]
- Jaillais, Y.; Chory, J. Unraveling the Paradoxes of Plant Hormone Signaling Integration. Nat. Struct. Mol. Biol. 2010, 17, 642–645. [Google Scholar] [CrossRef]
- Eichmann, R.; Richards, L.; Schäfer, P. Hormones as Go-Betweens in Plant Microbiome Assembly. Plant J. 2021, 105, 518–541. [Google Scholar] [CrossRef]
- Ortiz-García, P.; González Ortega-Villaizán, A.; Onejeme, F.C.; Müller, M.; Pollmann, S. Do Opposites Attract? Auxin-Abscisic Acid Crosstalk: New Perspectives. Int. J. Mol. Sci. 2023, 24, 3090. [Google Scholar] [CrossRef]
- Comai, L. The Advantages and Disadvantages of Being Polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef]
- Tsukaya, H. Controlling Size in Multicellular Organs: Focus on the Leaf. PLoS Biol. 2008, 6, e174. [Google Scholar] [CrossRef] [PubMed]
- del Pozo, J.C.; Ramirez-Parra, E. Whole Genome Duplications in Plants: An Overview from Arabidopsis. J. Exp. Bot. 2015, 66, 6991–7003. [Google Scholar] [CrossRef] [PubMed]
- Tavan, M.; Mirjalili, M.H.; Karimzadeh, G. In Vitro Polyploidy Induction: Changes in Morphological, Anatomical and Phytochemical Characteristics of Thymus persicus (Lamiaceae). Plant Cell Tissue Organ Cult. 2015, 122, 573–583. [Google Scholar] [CrossRef]
- Vergara, F.; Kikuchi, J.; Breuer, C. Artificial Autopolyploidization Modifies the Tricarboxylic Acid Cycle and GABA Shunt in Arabidopsis Thaliana Col-0. Sci. Rep. 2016, 6, 26515. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shi, X.; Hao, B.; Ge, S.; Luo, J. Duplication and DNA Segmental Loss in the Rice Genome: Implications for Diploidization. New Phytol. 2005, 165, 937–946. [Google Scholar] [CrossRef]
- Lavania, U.C. Genomic and Ploidy Manipulation for Enhanced Production of Phyto-Pharmaceuticals. Plant Genet. Resour. 2005, 3, 170–177. [Google Scholar] [CrossRef]
- He, N.; Umer, M.J.; Yuan, P.; Wang, W.; Zhu, H.; Lu, X.; Xing, Y.; Gong, C.; Batool, R.; Sun, X.; et al. Physiological, Biochemical, and Metabolic Changes in Diploid and Triploid Watermelon Leaves during Flooding. Front. Plant Sci. 2023, 14, 1108795. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, S.; Lu, X.; He, N.; Gao, L.; Dou, J.; Bie, Z.; Liu, W. Genome Duplication Improves the Resistance of Watermelon Root to Salt Stress. Plant Physiol. Biochem. 2018, 133, 11–21. [Google Scholar] [CrossRef]
- Tossi, V.E.; Martínez Tosar, L.J.; Laino, L.E.; Iannicelli, J.; Regalado, J.J.; Escandón, A.S.; Baroli, I.; Causin, H.F.; Pitta-Álvarez, S.I. Impact of Polyploidy on Plant Tolerance to Abiotic and Biotic Stresses. Front. Plant Sci. 2022, 13, 869423. [Google Scholar] [CrossRef]
- Ramsey, J.; Schemske, D.W.; Ramsey, J.; Schemske, D.W. Pathways, Mechanisms, and Rates of Polyploid Formation in Flowering Plants. Annu. Rev. Ecol. Syst. 1998, 29, 467–501. [Google Scholar] [CrossRef]
- Davis, A.R.; Webber, C.L.; Liu, W.; Perkins-Veazie, P.; Levi, A.; King, S. Watermelon Quality Traits as Affected by Ploidy. HortScience 2013, 48, 1113–1118. [Google Scholar] [CrossRef]
- Xu, M.; Gao, M.; Amanullah, S.; Guo, Y.; Bao, X.; Duan, Y.; Liu, X.; Liu, J.; Gao, Y.; Luan, F. Fine Genetic Mapping Confers a Major Gene Controlling Leaf Shape Variation in Watermelon. Euphytica 2023, 219, 92. [Google Scholar] [CrossRef]
- Mandizvo, T.; Odindo, A.O.; Mashilo, J.; Sibiya, J.; Beck-Pay, S.L. Phenotypic Variability of Root System Architecture Traits for Drought Tolerance among Accessions of Citron Watermelon (Citrullus lanatus Var. Citroides (L.H. Bailey). Plants 2022, 11, 2522. [Google Scholar] [CrossRef]
- Gebremeskel, H.; Dou, J.; Li, B.; Zhao, S.; Muhammad, U.; Lu, X.; He, N.; Liu, W. Molecular Mapping and Candidate Gene Analysis for GA3 Responsive Short Internode in Watermelon (Citrullus lanatus). Int. J. Mol. Sci. 2020, 21, 290. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, H.; Fan, M.; He, Y.; Guo, P. A Mutation in the Intron Splice Acceptor Site of a GA3ox Gene Confers Dwarf Architecture in Watermelon (Citrullus lanatus L.). Sci. Rep. 2020, 10, 14915. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, A.; He, W.; Li, Q.; Zhao, B.; Zhao, H.; Ke, X.; Guo, Y.; Sun, P.; Yang, T.; et al. GRAS Family Member LATERAL SUPPRESSOR Regulates the Initiation and Morphogenesis of Watermelon Lateral Organs. Plant Physiol. 2023, 193, 2592–2604. [Google Scholar] [CrossRef]
- Khan, M.N.-E.-A.; Hassan, J.; Biswas, M.S.; Khan, H.I.; Sultana, H.; Suborna, M.N.; Rajib, M.M.R.; Akter, J.; Gomasta, J.; Anik, A.A.M. Morphological and Anatomical Characterization of Colchicine-Induced Polyploids in Watermelon. Hortic. Environ. Biotechnol. 2023, 64, 461–474. [Google Scholar] [CrossRef]
- Zhang, N.; Bao, Y.; Xie, Z.; Huang, X.; Sun, Y.; Feng, G.; Zeng, H.; Ren, J.; Li, Y.; Xiong, J.; et al. Efficient Characterization of Tetraploid Watermelon. Plants 2019, 8, 419. [Google Scholar] [CrossRef]
- Jaskani, M.J.; Kwon, S.W.; Kim, D.H. Comparative Study on Vegetative, Reproductive and Qualitative Traits of Seven Diploid and Tetraploid Watermelon Lines. Euphytica 2005, 145, 259–268. [Google Scholar] [CrossRef]
- Feng, Z.; Bi, Z.; Fu, D.; Feng, L.; Min, D.; Bi, C.; Huang, H. A Comparative Study of Morphology, Photosynthetic Physiology, and Proteome between Diploid and Tetraploid Watermelon (Citrullus lanatus L.). Bioengineering 2022, 9, 746. [Google Scholar] [CrossRef]
- Mo, Z.; Feng, G.; Su, W.; Liu, Z.; Peng, F. Transcriptomic Analysis Provides Insights into Grafting Union Development in Pecan (Carya illinoinensis). Genes 2018, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Corneillie, S.; De Storme, N.; Van Acker, R.; Fangel, J.U.; De Bruyne, M.; De Rycke, R.; Geelen, D.; Willats, W.G.T.; Vanholme, B.; Boerjan, W. Polyploidy Affects Plant Growth and Alters Cell Wall Composition. Plant Physiol. 2019, 179, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Dudits, D.; Török, K.; Cseri, A.; Paul, K.; Nagy, A.V.; Nagy, B.; Sass, L.; Ferenc, G.; Vankova, R.; Dobrev, P.; et al. Response of Organ Structure and Physiology to Autotetraploidization in Early Development of Energy Willow Salix Viminalis. Plant Physiol. 2016, 170, 1504–1523. [Google Scholar] [CrossRef] [PubMed]
- Głowacka, K.; Jezowski, S.; Kaczmarek, Z. In Vitro Induction of Polyploidy by Colchicine Treatment of Shoots and Preliminary Characterisation of Induced Polyploids in Two Miscanthus Species. Ind. Crops Prod. 2010, 32, 88–96. [Google Scholar] [CrossRef]
- Kuspira, J.; Bhambmani, R.N.; Shimada, T. Genetic and Cytogenetic Analyses of the A Genome of Triticum Monococcum. I. Cytology, Breeding Behaviour, Fertility, and Morphology of Induced Autotetraploids. Can. J. Genet. Cytol. 1985, 27, 51–63. [Google Scholar] [CrossRef]
- Ma, Y.; Xue, H.; Zhang, L.; Zhang, F.; Ou, C.; Wang, F.; Zhang, Z. Involvement of Auxin and Brassinosteroid in Dwarfism of Autotetraploid Apple (Malus × Domestica). Sci. Rep. 2016, 6, 26719. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.J. Plant Cytogenetics; CRC Press: Boca Raton, FL, USA, 2016; ISBN 1439884196. [Google Scholar]
- Zhang, X.; Chen, K.; Wang, W.; Liu, G.; Yang, C.; Jiang, J. Differences in Leaf Morphology and Related Gene Expression between Diploid and Tetraploid Birch (Betula pendula). Int. J. Mol. Sci. 2022, 23, 12966. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhang, Y.; Han, Q.; Kang, X. Molecular Mechanism of Slow Vegetative Growth in Populus Tetraploid. Genes 2020, 11, 1417. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Zhang, S.; Xu, T.; Kang, X. Morphological, Transcriptome, and Hormone Analysis of Dwarfism in Tetraploids of Populus alba × P. glandulosa. Int. J. Mol. Sci. 2022, 23, 9762. [Google Scholar] [CrossRef]
- Rosellini, D.; Ferradini, N.; Allegrucci, S.; Capomaccio, S.; Zago, E.D.; Leonetti, P.; Balech, B.; Aversano, R.; Carputo, D.; Reale, L.; et al. Sexual Polyploidization in Medicago Sativa L.: Impact on the Phenotype, Gene Transcription, and Genome Methylation. G3 Genes Genomes Genet. 2016, 6, 925–938. [Google Scholar] [CrossRef]
- Van Norman, J.M.; Zhang, J.; Cazzonelli, C.I.; Pogson, B.J.; Harrison, P.J.; Bugg, T.D.H.; Chan, K.X.; Thompson, A.J.; Benfey, P.N. Periodic Root Branching in Arabidopsis Requires Synthesis of an Uncharacterized Carotenoid Derivative. Proc. Natl. Acad. Sci. USA 2014, 111, E1300–E1309. [Google Scholar] [CrossRef]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic Acid: Emergence of a Core Signaling Network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef]
- Yamaguchi, S. Gibberellin Metabolism and Its Regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Müller, M. Hormonal Cross-Talk in Plant Development and Stress Responses. Front. Plant Sci. 2013, 4, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Yuldashev, R.; Avalbaev, A.; Bezrukova, M.; Vysotskaya, L.; Khripach, V.; Shakirova, F. Cytokinin Oxidase Is Involved in the Regulation of Cytokinin Content by 24-Epibrassinolide in Wheat Seedlings. Plant Physiol. Biochem. 2012, 55, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Chone, R.M.S.; Rocha, D.I.; Monte-Bello, C.C.; Pinheiro, H.P.; Dornelas, M.C.; Haddad, C.R.B.; Almeida, J.A.S. Brassinosteroid Increases the Cytokinin Efficiency to Induce Direct Somatic Embryogenesis in Leaf Explants of Coffea arabica L. (Rubiaceae). Plant Cell Tissue Organ Cult. 2018, 135, 63–71. [Google Scholar] [CrossRef]
- Zu, S.-H.; Jiang, Y.-T.; Chang, J.-H.; Zhang, Y.-J.; Xue, H.-W.; Lin, W.-H. Interaction of Brassinosteroid and Cytokinin Promotes Ovule Initiation and Increases Seed Number per Silique in Arabidopsis. J. Integr. Plant Biol. 2022, 64, 702–716. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.J.; Quittenden, L.J. Interactions between Brassinosteroids and Gibberellins: Synthesis or Signaling? Plant Cell 2016, 28, 829–832. [Google Scholar] [CrossRef]
- Unterholzner, S.J.; Rozhon, W.; Papacek, M.; Ciomas, J.; Lange, T.; Kugler, K.G.; Mayer, K.F.; Sieberer, T.; Poppenberger, B. Brassinosteroids Are Master Regulators of Gibberellin Biosynthesis in Arabidopsis. Plant Cell 2015, 27, 2261–2272. [Google Scholar] [CrossRef]
- Xia, X.; Liu, Y.; Zhang, L.; Qi, Z.; Zhou, Y.; Yu, J. Overexpression of Brassinosteroid Synthesis Gene DWARF Promotes Resistance to Botrytis Cinerea by Inhibiting Gibberellin Synthesis in Solanum lycopersicum L. Plant Stress 2023, 9, 100170. [Google Scholar] [CrossRef]
- Bai, M.Y.; Shang, J.X.; Oh, E.; Fan, M.; Bai, Y.; Zentella, R.; Sun, T.P.; Wang, Z.Y. Brassinosteroid, Gibberellin and Phytochrome Impinge on a Common Transcription Module in Arabidopsis. Nat. Cell Biol. 2012, 14, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Gibberellins and Abscisic Acid Signal Crosstalk: Living and Developing under Unfavorable Conditions. Plant Cell Rep. 2013, 32, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Agharkar, M.; Lomba, P.; Altpeter, F.; Zhang, H.; Kenworthy, K.; Lange, T. Stable Expression of AtGA2ox1 in a Low-Input Turfgrass (Paspalum notatum Flugge) Reduces Bioactive Gibberellin Levels and Improves Turf Quality under Field Conditions. Plant Biotechnol. J. 2007, 5, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Mauriat, M.; Sandberg, L.G.; Moritz, T. Proper Gibberellin Localization in Vascular Tissue Is Required to Control Auxin-Dependent Leaf Development and Bud Outgrowth in Hybrid Aspen. Plant J. 2011, 67, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.K.; Case, D.B.; Jacobs, W.P. Auxin-Gibberellin Interaction in Apical Dominance. Plant Physiol. 1967, 42, 1329–1333. [Google Scholar] [CrossRef]
- Zawaski, C.; Busov, V.B. Roles of Gibberellin Catabolism and Signaling in Growth and Physiological Response to Drought and Short-Day Photoperiods in Populus Trees. PLoS ONE 2014, 9, e86217. [Google Scholar] [CrossRef] [PubMed]
- Choubane, D.; Rabot, A.; Mortreau, E.; Legourrierec, J.; Péron, T.; Foucher, F.; Ahcène, Y.; Pelleschi-Travier, S.; Leduc, N.; Hamama, L.; et al. Photocontrol of Bud Burst Involves Gibberellin Biosynthesis in Rosa sp. J. Plant Physiol. 2012, 169, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Elfving, D.C.; Visser, D.B.; Henry, J.L. Gibberellins Stimulate Lateral Branch Development in Young Sweet Cherry Trees in the Orchard. Int. J. Fruit Sci. 2011, 11, 41–54. [Google Scholar] [CrossRef]
- Koorneef, M.; Elgersma, A.; Hanhart, C.J.; van Loenen-Martinet, E.P.; Van Rijn, L.; Zeevaart, J.A.D. A Gibberellin Insensitive Mutant of Arabidopsis Thaliana. Physiol. Plant. 1985, 65, 33–39. [Google Scholar] [CrossRef]
- Marth, P.C.; Audia, W.V.; Mitchell, J.W. Effects of Gibberellic Acid on Growth and Development of Plants of Various Genera and Species. Bot. Gaz. 1956, 118, 106–111. [Google Scholar] [CrossRef]
- Ni, J.; Gao, C.; Chen, M.-S.; Pan, B.-Z.; Ye, K.; Xu, Z.-F. Gibberellin Promotes Shoot Branching in the Perennial Woody Plant Jatropha Curcas. Plant Cell Physiol. 2015, 56, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- González-Grandío, E.; Pajoro, A.; Franco-Zorrilla, J.M.; Tarancón, C.; Immink, R.G.H.; Cubas, P. Abscisic Acid Signaling Is Controlled by a BRANCHED1/HD-ZIP I Cascade in Arabidopsis Axillary Buds. Proc. Natl. Acad. Sci. USA 2017, 114, E245–E254. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Finlayson, S.A. Abscisic Acid Is a General Negative Regulator of Arabidopsis Axillary Bud Growth. Plant Physiol. 2015, 169, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Friedrichsen, D.M.; Joazeiro, C.A.P.; Li, J.; Hunter, T.; Chory, J. Brassinosteroid-Insensitive-1 Is a Ubiquitously Expressed Leucine-Rich Repeat Receptor Serine/Threonine Kinase. Plant Physiol. 2000, 123, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Y.; Khan, R.; Wu, X.; Zhou, L.; Xu, N.; Du, S.; Ma, X. Exogenous Application of Brassinosteroids Regulates Tobacco Leaf Size and Expansion via Modulation of Endogenous Hormones Content and Gene Expression. Physiol. Mol. Biol. Plants 2021, 27, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Fan, X.-Y.; Cao, D.-M.; Tang, W.; He, K.; Zhu, J.-Y.; He, J.-X.; Bai, M.-Y.; Zhu, S.; Oh, E.; et al. Integration of Brassinosteroid Signal Transduction with the Transcription Network for Plant Growth Regulation in Arabidopsis. Dev. Cell 2010, 19, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Szekeres, M.; Németh, K.; Koncz-Kálmán, Z.; Mathur, J.; Kauschmann, A.; Altmann, T.; Rédei, G.P.; Nagy, F.; Schell, J.; Koncz, C. Brassinosteroids Rescue the Deficiency of CYP90, a Cytochrome P450, Controlling Cell Elongation and de-Etiolation in Arabidopsis. Cell 1996, 85, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Kang, M.B. Brassinosteroid-Mediated Regulation of Agronomic Traits in Rice. Plant Cell Rep. 2014, 33, 683–696. [Google Scholar] [CrossRef]
- Ji, X.; Gao, Q.; Zhuang, Z.; Chang, F.; Peng, Y. WGCNA Analysis of the Effect of Exogenous BR on Leaf Angle of Maize Mutant Lpa1. Sci. Rep. 2024, 14, 5238. [Google Scholar] [CrossRef]
- Perez, M.B.M. Genetic and Brassinosteroid Control Underlying Leaf Angle Variation across the Canopy in Sorghum. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2018. [Google Scholar]
- Wang, Z.-Y.; Bai, M.-Y.; Oh, E.; Zhu, J.-Y. Brassinosteroid Signaling Network and Regulation of Photomorphogenesis. Annu. Rev. Genet. 2012, 46, 701–724. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmud, E.; Zhu, H.; Kaseb, M.O.; Sajjad, M.Z.; He, N.; Lu, X.; Liu, W. Polyploidization Impact on Plant Architecture of Watermelon (Citrullus lanatus). Horticulturae 2024, 10, 569. https://doi.org/10.3390/horticulturae10060569
Mahmud E, Zhu H, Kaseb MO, Sajjad MZ, He N, Lu X, Liu W. Polyploidization Impact on Plant Architecture of Watermelon (Citrullus lanatus). Horticulturae. 2024; 10(6):569. https://doi.org/10.3390/horticulturae10060569
Chicago/Turabian StyleMahmud, Eftekhar, Hongju Zhu, Mohamed Omar Kaseb, Muhammad Zeeshan Sajjad, Nan He, Xuqiang Lu, and Wenge Liu. 2024. "Polyploidization Impact on Plant Architecture of Watermelon (Citrullus lanatus)" Horticulturae 10, no. 6: 569. https://doi.org/10.3390/horticulturae10060569
APA StyleMahmud, E., Zhu, H., Kaseb, M. O., Sajjad, M. Z., He, N., Lu, X., & Liu, W. (2024). Polyploidization Impact on Plant Architecture of Watermelon (Citrullus lanatus). Horticulturae, 10(6), 569. https://doi.org/10.3390/horticulturae10060569