
Echeveria Leaf Morpho-Anatomical Analysis and Its Implications for Environmental Stress Conditions


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Morphological Evaluation
2.3. Free-Hand Sectioning and Fixation Method
2.4. Leaf-Morpho-Anatomical Observation
2.5. Moisture Content Measurement
2.6. Statistical and Cluster Analysis
3. Results
3.1. Leaf Morphology Evaluation
3.2. Leaf Morpho-Anatomical Evaluation and Its Environmental Implication
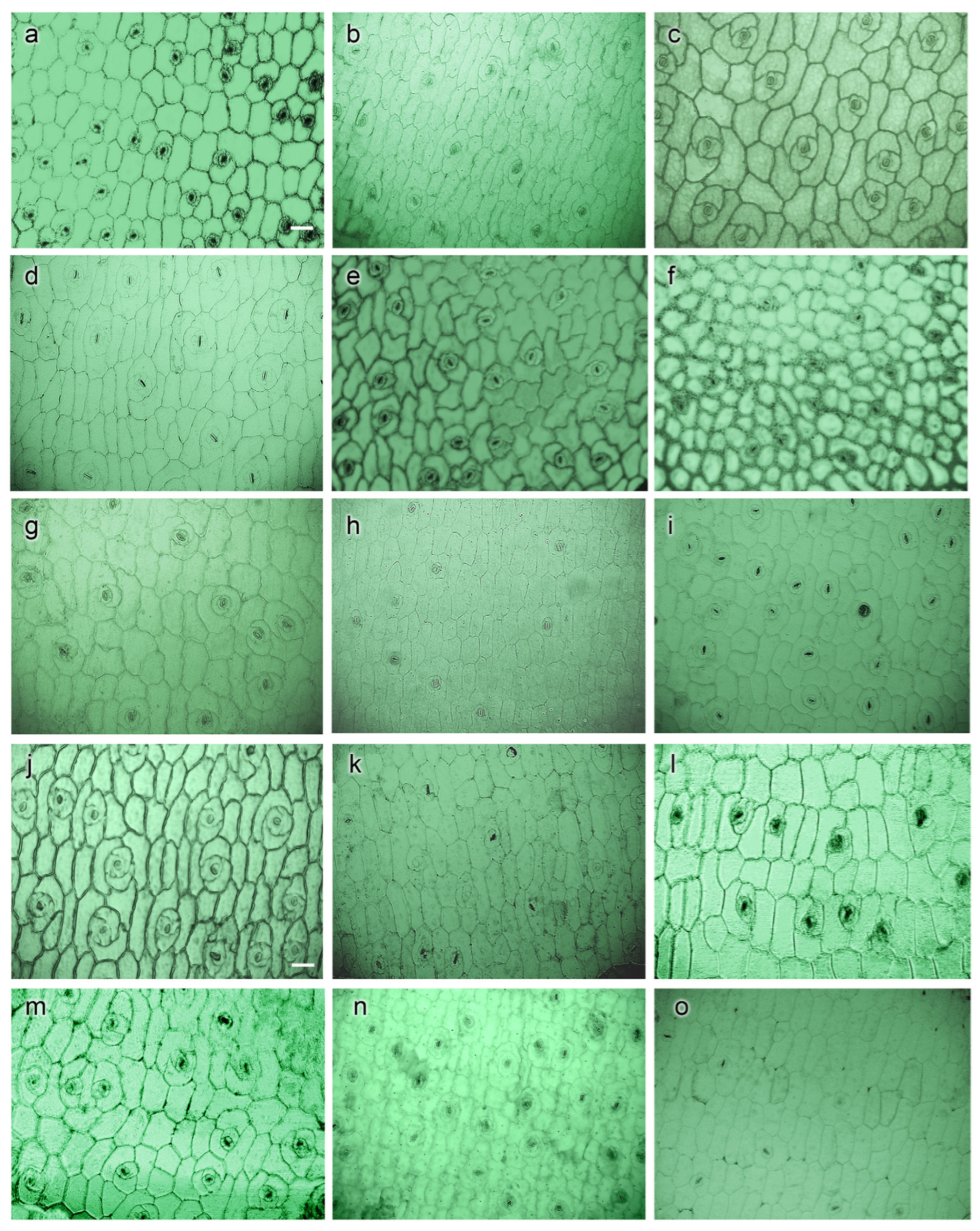
3.2.1. Epidermis and Hypodermis
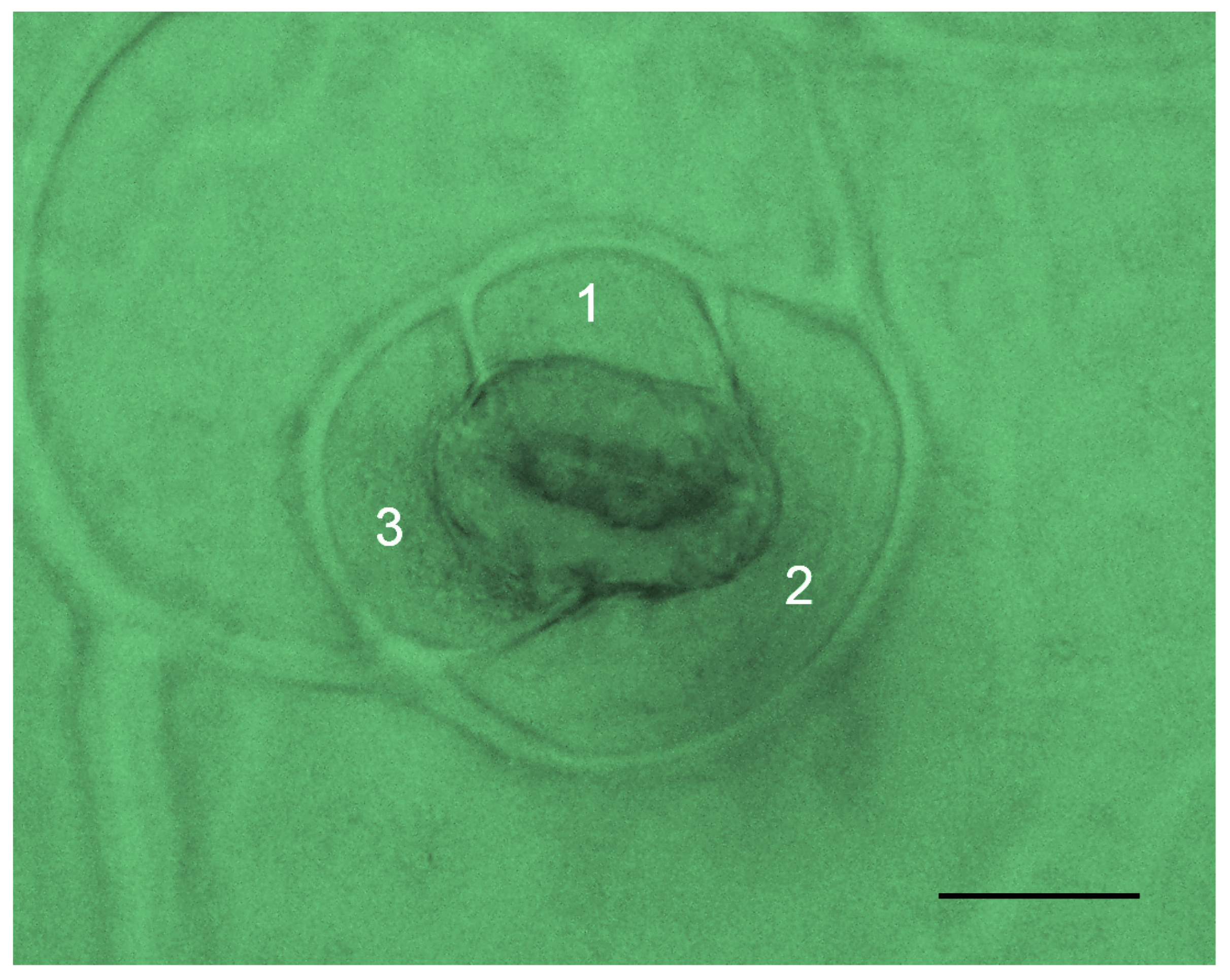
3.2.2. Stomata Characteristics
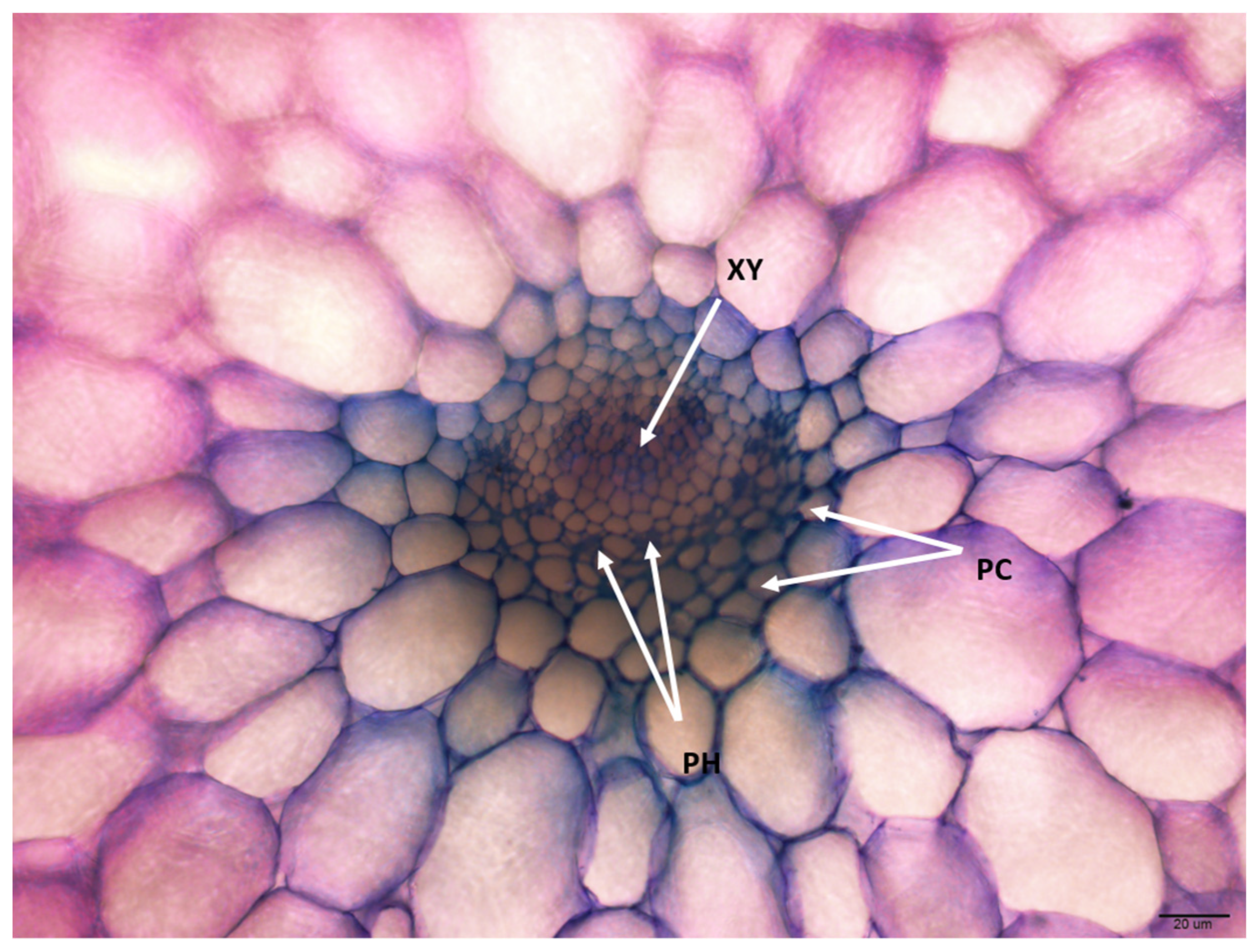
3.2.3. Vascular Bundles
3.3. Moisture Content
3.4. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- von Willert, D.J.; Eller, B.M.; Werger, M.J.A.; Brinckmann, E.; Ihlenfeldt, H.D. Life strategies of succulents in deserts. Vegetation 1990, 90, 131–143. [Google Scholar] [CrossRef]
- Eggli, U.; Nyffeler, R. Living under temporarily arid conditions succulence as an adaptive strategy. Bradleya 2009, 27, 13–36. [Google Scholar] [CrossRef]
- Van Woert, N.D.; Rowe, D.B.; Andresen, J.A.; Rugh, C.L.; Xiao, L. Watering regime and green roof substrate design affect Sedum plant growth. HortScience 2005, 40, 659–664. [Google Scholar] [CrossRef]
- Velasco, E.; Roth, M.; Norford, L.K.; Molina, L.T. Does urban vegetation enhance carbon sequestration? Landsc. Urban Plan. 2016, 148, 99–107. [Google Scholar] [CrossRef]
- Kim, I.H.; Huh, K.Y.; Huh, M.R. Cold tolerance assessment of Sedum species for shallow extensive green roof system. Korean J. Hortic. Sci. Technol. 2010, 28, 22–30. [Google Scholar]
- Francini, A.; Romano, D.; Toscano, S.; Ferrante, A. The contribution of ornamental plants to urban ecosystem services. Earth 2022, 3, 1258–1274. [Google Scholar] [CrossRef]
- Kluge, M.; Lange, O.L.; Eichmann, M.V.; Schmid, R. CAM in Tillandsia usneoides: Studies on the pathway of carbon and the dependency of CO2–exchange on light intensity, temperature and water content of the plant. Planta 1973, 112, 357–372. [Google Scholar] [CrossRef]
- Ogburn, R.M.; Edwards, E.J. The ecological water-use strategies of succulent plants. Adv. Bot. Res. 2010, 55, 179–225. [Google Scholar]
- Li, F.L.; Bao, W.K. Elevational trends in leaf size of Campylotropis polyantha in the arid Minjiang River Valley, SW China. J. Arid Environ. 2014, 108, 1–9. [Google Scholar] [CrossRef]
- Larbi, A.; Vázquez, S.; El-Jendoubi, H.; Msallem, M.; Abadia, J.; Abaia, A.; Morales, F. Canopy light heterogeneity drives leaf anatomical, ecophysiological, and photosynthetic changes in olive trees grown in a high-density plantation. Photosynth. Res. 2014, 123, 141–155. [Google Scholar] [CrossRef]
- Jones, L.A. Anatomical adaptations of four Crassula species to water availability. Biosci. Horiz. 2011, 4, 13–22. [Google Scholar] [CrossRef]
- Royer, D.L.; McElwain, J.C.; Adams, J.M.; Wilf, P. Sensitivity of leaf size and shape to climate within Acer rubrum and Quercus kelloggii. New Phytol. 2008, 179, 808–817. [Google Scholar] [CrossRef]
- Hu, Y.; Yang, L.; Gao, C.; Liao, D.S.; Long, L.; Jie, Q.; Wei, H.L.; Deng, Q.N.; Zhou, Y.C. A comparative study on the leaf anatomical structure of Camellia oleifera in a low-hot valley area in Guizhou Province, China. PLoS ONE 2021, 17, e0262509. [Google Scholar] [CrossRef]
- Wyka, T.P.; Bagniewska-Zadworna, A.; Kuczyńska, A.; Mikołajczak, K.; Ogrodowicz, P.; Żytkowiak, M.; Surma, M.; Adamski, T. Drought-induced anatomical modifications of barley (Hordeum vulgare L.) leaves: An allometric perspective. Environ. Exp. Bot. 2019, 166, 103798. [Google Scholar] [CrossRef]
- Taratima, W.; Ritmaha, T.; Jongrungklang, N.; Maneerattanarungroj, P.; Kunpratum, N. Effect of stress on the leaf anatomy of sugarcane cultivars with different drought tolerance (Saccharum officinarum, Poaceae). Rev. Biol. Trop. 2020, 68, 1159–1170. [Google Scholar] [CrossRef]
- Griffiths, H.; Males, J. Succulent plants. Curr. Biol. 2017, 27, 890–896. [Google Scholar] [CrossRef]
- Nobel, P.S. Environmental Biology of Agaves and Cacti; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Kluge, M.; Ting, I.P. Crassulacean Acid Metabolism: Analysis of an Ecological Adaptation; Springer: Berlin, Germany, 1978; pp. 5–44. [Google Scholar]
- Nobel, P.S. Environmental influences on CO2 uptake by agaves, CAM plants with high productivities. Econ. Bot. 1990, 44, 488–502. [Google Scholar] [CrossRef]
- Grace, O.M. Succulent plant diversity as natural capital. Plant People Planet 2019, 1, 336–345. [Google Scholar] [CrossRef]
- Manokari, M.; Cokulraj, M.; Badhepuri, M.K.; Dey, A.; Faisal, M.; Alatar, A.A.; Singh, R.K.; Shekhawat, M.S. Microstructural and histochemical modifications in leaves at successive stages of in vitro development of the terrestrial orchid Spathoglottis plicata Blume. Hortic. Environ. Biotechnol. 2023, 64, 497–510. [Google Scholar] [CrossRef]
- Cabahug-Braza, R.A.; Tran, M.K.T.H.; Lim, K.B.; Hwang, Y.J. Phenotypic evaluation and nuclear DNA content analysis of oryzalin-induced Echeveria mutant cultivars. Hortic. Sci. Technol. 2023, 41, 315–328. [Google Scholar] [CrossRef]
- Khan, M.N.E.A.; Hassan, J.; Biswas, M.S.; Khan, H.I.; Sultana, H.; Suborna, M.N.; Rajib, M.M.R.; Akter, J.; Gomasta, J.; Anik, A.A.M. Morphological and anatomical characterization of colchicine-induced polyploids in watermelon. Hortic. Environ. Biotechnol. 2023, 64, 461–474. [Google Scholar] [CrossRef]
- Yeung, E. A beginner’s guide to the study of plant structure. In Tested Studies for Laboratory Teaching; Karche, S.J., Ed.; Purdue University: West Lafayette, IN, USA, 1998; pp. 125–141. [Google Scholar]
- Mohammed, F.; Arishiya, T.F.; Mohamed, S. Microtomes and microtome knives—A review and proposed classification. Ann. Dent. 2012, 19, 43–50. [Google Scholar]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Company Inc.: London, UK, 1940; p. 530. [Google Scholar]
- Ribeiro, V.C.; Leitão, C.A.E. Utilisation of toluidine blue O pH 4.0 and histochemical inferences in plant sections obtained by free-hand. Protoplasma 2020, 257, 993–1008. [Google Scholar] [CrossRef]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- Thierry, R.; Marie-Christine, R.; Marie’Le, M.; Richard, B.; Anca, M. Modified technique of Toluidine blue staining in rapid on-site evaluation. Diagn. Cytopathol. 2011, 40, 847–848. [Google Scholar] [CrossRef]
- Parker, A.J.; Haskins, E.F.; Deyrup-Olsen, I. Toluidine blue: A simple, effective stain for plant tissues. Am. Biol. Teach. 1982, 44, 487–489. [Google Scholar] [CrossRef]
- Koller, A.L.; Rost, T.L. Structural analysis of water-storage tissue in leaves of Sansevieria (Agavaceae). Bot. Gaz. 1988, 149, 260–274. [Google Scholar] [CrossRef]
- Fradera-Soler, M.; Rudall, P.J.; Prychid, C.J.; Grace, O.M. Evolutionary success in arid habitats: Morpho-anatomy of succulent leaves of Crassula species from southern Africa. J. Arid Environ. 2021, 185, 104319. [Google Scholar] [CrossRef]
- Nyffeler, R.; Eggli, U. An up-to-date familial and suprafamilial classification of succulent plants. Bradleya 2010, 28, 125–144. [Google Scholar] [CrossRef]
- Uhl, C.H. Polyploidy, dysploidy, and chromosome pairing in Echeveria (Crassulaceae) and its hybrid. Am. J. Bot. 1992, 79, 556–566. [Google Scholar] [CrossRef]
- Cabahug, R.A.M.; Nam, S.Y.; Lim, K.B.; Jeon, J.K.; Hwang, Y.J. Propagation techniques for ornamental succulents. Flower Res. J. 2018, 26, 90–101. [Google Scholar] [CrossRef]
- Borys, M.W.; Leszczyńska-Borys, H. The genus Echeveria as a potential new floral crop. Acta Hortic. 2013, 1000, 91–96. [Google Scholar] [CrossRef]
- Males, J. Secrets of succulence. J. Exp. Bot. 2017, 68, 2121–2134. [Google Scholar] [CrossRef]
- Razzaghmanesh, M.; Beechman, S.; Kazemi, F. The growth and survival of plants in urban green roofs in a dry climate. Sci. Total Environ. 2014, 476, 288–297. [Google Scholar] [CrossRef]
- Sandoval-Zapotitla, E.; Martínez-Quezada, D.M.; Reyes-Santiago, J.; Islas-Luna, M.; Rosas, U. Leaf morpho-anatomical diversity in Echeveria aff. gigantea (Crassulaceae). Bot. Sci. 2019, 97, 218–235. [Google Scholar] [CrossRef]
- Matimati, I.; Musil, C.F.; Raitt, L.; February, E. Non-rainfall moisture interception by dwarf succulents and their relative abundance in an inland arid South African ecosystem. Ecohydrology 2013, 6, 818–825. [Google Scholar] [CrossRef]
- Teskey, R.O. Canopy Processes. In Tree Physiology (Encyclopedia of Forest Sciences); Elsevier Ltd.: Amsterdam, The Netherlands, 2004; pp. 1622–1628. [Google Scholar]
- Francis, B.; Gilman, R.T. Light intensity affects leaf morphology in a wild population of Adenostyles alliariae (Asteraceae). Ital. Bot. 2019, 8, 35–45. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Leigh, A.; Boyce, C.K.; Jones, C.S.; Niklas, K.J.; Royer, D.L.; Tsukaya, H. The evolution and functional significance of leaf shape in the angiosperms. Funct. Plant Biol. 2011, 38, 535–552. [Google Scholar] [CrossRef]
- Rost, T.L. Vascular pattern and hydathodes in leaves of Crassula argentea (Crassulaceae). Bot. Gaz. 1969, 130, 267–270. [Google Scholar] [CrossRef]
- Pilbeam, J. The Genus Echeveria; The British Cactus & Succulent Society: Great Malvern, UK, 2008. [Google Scholar]
- Ash, A.; Ellis, B.; Hickey, L.J.; Johnson, K.; Wilf, P.; Wing, S. Manual of Leaf Architecture; Smithsonian Institution: Washington, DC, USA, 1999; pp. 12–67. [Google Scholar]
- Neinhuis, C.; Edelmann, H.G. Methanol as a rapid fixative for the investigation of plant surfaces by SEM. J. Microsc. 1996, 184, 14–16. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Salisbury, E.J. On the causes and ecological significance of stomata frequency with special reference to the woodland flora. Philos. Trans. R. Soc. Lond. 1927, 216, 1–65. [Google Scholar]
- Lee, J.H.; Nam, S.Y. Analysis of growth and leaf color changes of Sedum album cv. Athoum according to the spectral power distribution of several white LEDs. Flower Res. J. 2022, 30, 184–193. [Google Scholar] [CrossRef]
- Wright, I.J.; Dong, N.; Maire, V.; Prentice, I.C.; Westoby, M.; Díaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A.; et al. Global climatic drivers of leaf size. Science 2017, 357, 917–921. [Google Scholar] [CrossRef] [PubMed]
- Long, L.M.; Patel, H.P.; Cory, W.C.; Stapleton, A.E. The maize epicuticular wax layer provides UV protection. Funct. Plant Biol. 2003, 30, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Chernetskyy, M.; Weryszko-Chmielewska, E. Structure of Kalanchoë pumila Bak. leaves (Crassulaceae DC). Acta Agrobot. 2008, 61, 11–24. [Google Scholar] [CrossRef]
- Denton, M.F. SEM Analysis of leaf epicuticular waxes of Sedum Section Gormania (Crassulaceae). Brittonia 1994, 46, 296–308. [Google Scholar] [CrossRef]
- Miroslavov, E.A. Structure and Function of Leaf Epidermis of Angiosperms Plants; Nauka: St. Petersburg, Russia, 1974; pp. 12–54. [Google Scholar]
- Niklas, K.J. Evolution of plant shape: Design constraints. Trends Ecol. Evol. 1986, 1, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Schulze, E.D.; Beck, E.; Muller-Hohenstein, K. Plant Ecology; Springer: Berlin, Germany, 2005; pp. 61–79. [Google Scholar]
- Tomlinson, P.B. Commelinales-Zingiberales. In Anatomy of the Monocotyledons; Metcalfe, C.R., Ed.; Oxford Press: Sydney, Australia, 1969; pp. 81–92. [Google Scholar]
- Kaul, R.B. The role of the multiple epidermis in foliar succulence in Peperomia (Piperaceae). Bot. Gaz. 1977, 138, 213–218. [Google Scholar] [CrossRef]
- De Micco, V.; Aronne, G. Morpho–anatomical traits for plant adaptation to drought. In Plant Responses to Drought Stress; Springer: Berlin, Germany, 2012; pp. 37–61. [Google Scholar]
- Dickison, W. Integrative Plant Anatomy; Academic Press: San Diego, CA, USA, 2000; p. 533. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledones, 2nd ed.; Clarendon Press: Oxford, UK, 1979; pp. 456–473. [Google Scholar]
- Stace, C.A. The taxonomic importance of the leaf surface. In Current Concepts in Plant Taxonomy; Heywood, V.H., Moore, D.M., Eds.; Academic Press: London, UK, 1984; pp. 67–93. [Google Scholar]
- Sapala, A.; Runions, A.; Routier-Kierzkowska, A.-L.; Gupta, M.D.; Hong, L.; Hofhuis, H.; Verger, S.; Mosca, G.; Li, C.-B.; Hay, A.; et al. Why plants make puzzle cells, and how their shape emerges. eLife 2018, 7, e32794. [Google Scholar] [CrossRef]
- Sapala, A.; Runions, A.; Smith, R.S. Mechanics, geometry and genetics of epidermal cell shape regulation: Different pieces of the same puzzle. Curr. Opin. Plant Biol. 2019, 47, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gang, Z.; Liu, B.; Rohwer, J.G.; Ferguson, D.K.; Yang, Y. Leaf epidermal micromorphology defining the clades in Cinnamomum (Lauraceae). PhytoKeys 2021, 4, 125–148. [Google Scholar] [CrossRef]
- Plaza, L.; Fernandez, I.; Juan, R.; Pastor, J.; Pujadas, A. Micromorphological studies on seeds of Orobanche species from the Iberian Peninsula and the Balearic Islands, and their systematic significance. Ann. Bot. 2004, 94, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Moraes, T.M.; Barros, C.F.; Silva Neto, S.J.; Gomes, V.M.; Da Cunha, M. Leaf blade anatomy and ultrastructure of six Simira species (Rubiaceae) from the Atlantic rain forest, Brazil. Biocell 2009, 33, 155–165. [Google Scholar] [CrossRef]
- Riederer, M.; Muller, C. Biology of the Plant Cuticle; Blackwell Publishing Ltd.: Oxford, UK, 2006; pp. 12–56. [Google Scholar]
- Stace, C.A. Cuticular studies as an aid to plant taxonomy. Bull. Br. Mus. Bot. 1965, 4, 3–78. [Google Scholar]
- Song, J.H.; Yang, S.G.; Choi, G.Y. Taxonomic implications of leaf micromorphology using microscopic analysis: A tool for identification and authentication of Korean Puperales. Plants 2020, 9, 566. [Google Scholar] [CrossRef]
- Mott, K.A.; Michaelson, O. Amphistomy as an adaptation to high light intensity in Ambrosia cordifolia (Compositae). Am. J. Bot. 1991, 78, 76–79. [Google Scholar] [CrossRef]
- Jordan, G.J.; Carpenter, R.J.; Brodribb, T.J. Using fossil leaves as evidence for open vegetation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 395, 168–175. [Google Scholar] [CrossRef]
- Willmer, C.; Fricker, M. Stomata; Springer: Dordrecht, The Netherlands, 1996; pp. 73–76. [Google Scholar]
- Pérez, V.; Arévalo, A.; Villanueva-Almanza, L.; Ezcurra, E. Variation in leaf xeromorphism in the desert palm genus Washingtonia (Arecaceae). J. Arid Environ. 2021, 186, 104412. [Google Scholar] [CrossRef]
- Parkhurst, D.F. The adaptive significance of stomatal occurence on one or both surfaces of leaves. J. Ecol. 1978, 66, 367–383. [Google Scholar] [CrossRef]
- Prabhakar, M. Structure, delimitation, nomenclature and classification of stomata. Acta Bot. Sin. 2004, 46, 242–252. [Google Scholar]
- Nunes, T.D.G.; Zhang, D.; Raissig, M.T. Form, development and function of grass stomata. Plant J. 2020, 101, 780–799. [Google Scholar] [CrossRef]
- Munemasa, S.; Hauser, F.; Park, J.; Waadt, R.; Brandt, B.; Schroeder, J.I. Mechanisms of abscisic acid-mediated control of stomatal aperture. Curr. Opin. Plant Biol. 2015, 28, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Mori, I.C.; Munemasa, S. Diverse stomatal signaling and the signal integration mechanism. Annu. Rev. Plant Biol. 2015, 66, 369–392. [Google Scholar] [CrossRef] [PubMed]
- Sunberg, W.; Marshall, D. A comparison of stomatal distribution and length in succulent and non-succulent desert plants. Phytomorphology 1986, 36, 53–66. [Google Scholar]
- Wall, S.; Vialet-Chabrand, S.; Davey, P.; Van Rie, J.; Galle, A.; Cockram, J.; Lawson, T. Stomata on the abaxial and adaxial leaf surfaces contribute differently to leaf gas exchange and photosynthesis in wheat. New Phytol. 2022, 235, 1743–1756. [Google Scholar] [CrossRef]
- Buttery, B.R.; Tan, C.S.; Buzzell, R.I.; Gaynor, J.D.; MacTavish, D.C. Stomatal numbers of soybean and response to water stress. Plant Soil 1993, 149, 283–288. [Google Scholar] [CrossRef]
- Ogburn, R.M.; Edwards, E.J. Repeated origin of three-dimensional leaf venation releases constraints on the evolution of succulence in plants. Curr. Biol. 2013, 23, 722–726. [Google Scholar] [CrossRef]
- Caballero-Ruano, A.; Jiménez-Parrondo, M.S. A contribution to the leaf anatomy studies of Canarian Crasuláceas. Vieraea 1978, 7, 115–132. [Google Scholar]
- Melo-de-Pinna, G.F.A.; Hernades-Lopes, J.; Ogura, A.S.; Santos, L.K.; Silva, D.C.; Haevermans, T. Growth patterns and different arrangements of vascular tissues in succulent leaves. Int. J. Plant Sci. 2016, 177, 643–660. [Google Scholar] [CrossRef]
- Ardelean, M.; Stanescu, I.; Cachita-Cosma, D. Sedum telephium spp. Maximum (L.) Krock. histo-anatomical aspects on the vegetative organs. J. Plant Dev. 2009, 55, 75–80. [Google Scholar]
- Ihlenfeldt, H.-D. Life forms and survival strategies in succulents. Rep. Ger. Bot. Soc. 1985, 98, 409–423. [Google Scholar]
- Ievinsh, G. Water content of plant tissues: So simple that almost forgotten? Plants 2023, 12, 1238. [Google Scholar] [CrossRef] [PubMed]
- Bousselot, J.M.; Klett, J.E.; Koski, R.D. Moisture content of extensive green roof substrate and growth response of 15 temperate plant species during dry down. HortScience 2011, 46, 518–522. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Leaf Characteristics | Other Distinct Features | ||||||
|---|---|---|---|---|---|---|---|---|
| Shape z | Margin y | Apex x | Edge Pattern w | Epicuticular Wax (EW) | EW Amount v | Trichomes | Branching | |
| E. ‘Benbadis’ | 1 | 1 | 1 | + | + | 1 | − | − |
| E. ‘Brave’ | 2 | 1 | 2 | + | + | 1 | − | − |
| E. colorata E. Walther | 2 | 1 | 1 | + | + | 1 | − | − |
| E. ‘Cubic Frost’ | 1 | 1 | 2 | + | + | 2 | − | − |
| E. ‘Dark Ice’ | 2 | 1 | 1 | + | + | 1 | − | − |
| E. ‘Doterang’ | 1 | 1 | 1 | + | + | 2 | − | − |
| E. ‘Fire Pillar’ | 1 | 1 | 1 | + | + | 1 | − | − |
| E. ‘Glam Pink’ | 1 | 1 | 4 | + | + | 1 | − | − |
| E. ‘Loy’ | 1 | 1 | 1 | - | + | 1 | − | − |
| E. ‘Milk Rose’ | 1 | 1 | 4 | + | + | 2 | − | − |
| E. ‘Peerless’ | 2 | 1 | 2 | + | + | 2 | − | − |
| E. ‘Snow Bunny’ | 1 | 1 | 2 | + | + | 3 | − | − |
| E. ‘Silhouette’ | 1 | 1 | 2 | + | + | 1 | − | − |
| E. ‘Tippy’ | 2 | 1 | 1 | + | + | 1 | − | − |
| E. ‘Viyant’ | 2 | 1 | 3 | + | + | 3 | − | − |
| Cultivar | Plant Parameters (mm) | Leaf Parameters (mm) | ||||
|---|---|---|---|---|---|---|
| Height | Diameter | Length | Width | Thickness | No. of Leaves | |
| E. ‘Benbadis’ | 37.54 ±2.40 z | 55.4 ± 1.61 | 26.53 ± 1.18 | 17.38 ± 0.84 | 4.65 ± 0.29 | 46.33 ± 2.33 |
| E. ‘Brave’ | 25.60 ± 1.44 | 62.33 ± 2.22 | 28.40 ± 1.00 | 15.31 ± 1.03 | 7.21 ± 0.25 | 19.67 ± 0.33 |
| E. colorata E. Walther | 39.11 ± 1.35 | 82.36 ± 3.51 | 37.91 ± 0.58 | 19.11 ± 0.60 | 6.11 ± 0.39 | 31.00 ± 0.58 |
| E. ‘Cubic Frost’ | 35.40 ± 2.75 | 72.00 ± 1.04 | 33.80 ± 0.94 | 24.04 ± 0.46 | 4.85 ± 0.34 | 28.00 ± 1.15 |
| E. ‘Dark Ice’ | 38.62 ± 1.02 | 70.95 ± 0.77 | 32.58 ± 2.29 | 17.88 ± 0.75 | 4.98 ± 0.16 | 22.67 ± 0.88 |
| E. ‘Doterang’ | 33.00 ± 4.22 | 66. 73 ± 2.01 | 30.88 ± 3.33 | 15.81 ± 0.84 | 5.81 ± 0.45 | 24.67 ± 0.88 |
| E. ‘Fire Pillar’ | 37. 10 ± 0.64 | 63.01 ± 0.34 | 30.25 ± 1.88 | 21.16 ± 1.27 | 6.01 ± 0.13 | 25.33 ± 2.33 |
| E. ‘Glam Pink’ | 46.98 ± 0.53 | 77.69 ± 4.49 | 39.24 ± 0.99 | 18.09 ± 1.28 | 6.24 ± 0.13 | 20.33 ± 3.84 |
| E. ‘Loy’ | 49.10 ± 1.31 | 83.07 ± 0.54 | 39.49 ± 1.61 | 21.84 ± 0.13 | 6.67 ± 0.09 | 42.33 ± 1.76 |
| E. ‘Milk Rose’ | 41. 14 ± 2.70 | 93.72 ± 4.45 | 49.15 ± 2.24 | 26.19 ± 0.65 | 8.21 ± 0.66 | 27.33 ± 0.88 |
| E. ‘Peerless’ | 28.60 ± 0.85 | 75.56 ± 2.27 | 35.47 ± 1.40 | 21.47 ± 1.31 | 7.34 ± 0.28 | 27.00 ± 1.00 |
| E. ‘Silhouette’ | 40.02 ± 3.08 | 75.65 ± 6.53 | 30.97 ± 1.67 | 20.65 ± 1.65 | 5.34 ± 0.07 | 37.33 ± 2.18 |
| E. ‘Snow Bunny’ | 30.85 ± 0.61 | 46.59 ± 2.50 | 22.25 ± 0.66 | 15.63 ± 0.33 | 6.52 ± 0.27 | 17.33 ± 1.21 |
| E. ‘Tippy’ | 39. 34 ± 2.20 | 29.28 ± 1.31 | 29.28 ± 1.31 | 14.57 ± 0.02 | 5.12 ± 0.51 | 39.67 ± 1.76 |
| E. ‘Viyant’ | 27.01 ± 1.90 | 26.50 ± 3.95 | 26.50 ± 3.95 | 13.42 ± 0.45 | 6.05 ± 0.11 | 35.33 ± 3.53 |
| Cultivar | Epidermis | Hypodermis | ||
|---|---|---|---|---|
| Width | Length | Width | Length | |
| E. ‘Benbadis’ | 17.11 ± 0.67 x | 52.50 ± 3.23 | 72.65 ± 1.30 | 64.84 ± 4.78 |
| E. ‘Brave’ | 27.81 ± 1.04 | 67.20 ± 2.94 | 133.20 ± 1.86 | 114.14 ± 10.20 |
| E. colorata E. Walther | 38.44 ± 0.85 | 119.18 ± 8.91 | 95.80 ± 2.50 | 92.99 ± 2.19 |
| E. ‘Cubic Frost’ | 23.67 ± 1.70 | 47.96 ± 3.00 | 49.16 ± 1.3 | 47.46 ± 2.21 |
| E. ‘Dark Ice’ | 26.87 ± 1.31 | 82.62 ± 3.42 | 74.59 ± 3.22 | 120.10 ± 6.16 |
| E. ‘Doterang’ | 38.49 ± 2.52 | 63.58 ± 3.92 | 73.11 ± 1.64 | 93.46 ± 3.66 |
| E. ‘Glam Pink’ | 31.81 ± 0.97 | 64.51 ± 4.02 | 131.85 ± 3.37 | 98.56 ± 2.15 |
| E. ‘Fire Pillar’ | 19.82 ± 0.19 | 62.14 ± 6.43 | 81.16 ± 1.11 | 83.20 ± 0.80 |
| E. ‘Loy’ | 26.71 ± 3.19 | 73.59 ± 4.18 | 129.00 ± 22.74 | 112.68 ± 10.30 |
| E. ‘Milk Rose’ | 24.51 ± 1.72 | 88.07 ± 5.80 | 70.65 ± 2.89 | 75.10 ± 2.32 |
| E. ‘Peerless’ | 29.67 ± 1.33 | 62.07 ± 0.56 | 112.05 ± 6.93 | 87.83 ± 0.74 |
| E. ‘Snow Bunny’ | 40.18 ± 1.37 | 62.25 ± 1.64 | 74.57 ± 1.21 | 91.78 ± 11.33 |
| E. ‘Silhouette’ | 25.60 ± 1.52 | 53.96 ± 4.68 | 65.28 ± 5.22 | 77.17 ± 6.93 |
| E. ‘Tippy’ | 23.99 ± 1.05 | 53.03 ± 0.81 | 44.85 ± 2.26 | 54.27 ± 1.79 |
| E. ‘Viyant’ | 16.65 ± 1.69 | 45.60 ± 2.74 | 68.54 ± 3.12 | 56.45 ± 3.64 |
| F-test z | ** | ** | ** | ** |
| Cultivar | Epidermal Cell Shape | Periclinal Wall | Anticlinal Cell Wall |
|---|---|---|---|
| E. ‘Benbadis’ | Polygonal | Non-reticulate | Sinuous |
| E. ‘Brave’ | Polygonal | Non-reticulate | Sinuous |
| E. colorata E. Walther | Polygonal | Non-reticulate | Straight |
| E. ‘Cubic frost’ | Polygonal | Reticulate | Nearly Straight |
| E. ‘Dark Ice’ | Irregular | Reticulate | Sinuous |
| E. ‘Doterang’ | Polygonal | Non-reticulate | Straight |
| E. ‘Fire Pillar’ | Polygonal | Reticulate | Sinuous |
| E. ‘Glam Pink’ | Polygonal | Non-reticulate | Straight |
| E. ‘Loy’ | Polygonal | Non-reticulate | Straight |
| E. ‘Milk Rose’ | Polygonal | Non-reticulate | Straight |
| E. ‘Peerless’ | Polygonal | Non-reticulate | Straight |
| E. ‘Silhouette’ | Polygonal | Non-reticulate | Straight |
| E. ‘Snow Bunny’ | Polygonal | Non-reticulate | Straight |
| E. ‘Tippy’ | Irregular | Reticulate | Sinuous |
| E. ‘Viyant’ | Polygonal | Non-reticulate | Straight |
| Cultivar | Stomata Size (µm) | Stomata Density (mm−2) | Stomata Index (%) | ||
|---|---|---|---|---|---|
| Adaxial | Abaxial | Adaxial | Abaxial | ||
| E. ‘Benbadis’ | 21.23 ± 0.04 z | 18.21 ± 0.12 | 17.33 ± 2.85 | 9.00 ± 0.58 | 11.66 |
| E. ‘Brave’ | 17.06 ± 0.04 | 18.64 ± 0.07 | 14.50 ± 0.50 | 11.50 ± 0.50 | 18.26 |
| E. colorata E. Walther | 18.74 ± 0.77 | 18.75 ± 0.21 | 20.33 ± 4.06 | 10.00 ± 1.15 | 22.96 |
| E. ‘Cubic Frost’ | 21.60 ± 1.20 | 21.50 ± 1.20 | 14.67 ± 6.69 | 7.33 ± 2.85 | 10.39 |
| E. ‘Dark Ice’ | 17.31 ± 1.50 | 17.41 ± 1.75 | 18.25 ± 3.47 | 14.25 ± 2.95 | 18.75 |
| E. ‘Doterang’ | 18.20 ± 0.26 | 17.31 ± 0.22 | 33.67 ± 9.39 | 17.00 ± 1.53 | 20.87 |
| E. ‘Fire Pillar’ | 21.61 ± 0.12 | 21.46 ± 0.13 | 11.67 ± 0.88 | 8.33 ± 0.33 | 12.73 |
| E. ‘Glam Pink’ | 17.70 ± 0.07 | 17.56 ± 0.09 | 14.67 ± 0.33 | 14.33 ± 0.88 | 11.11 |
| E. ‘Loy’ | 19.30 ± 0.03 | 19.25 ± 0.11 | 24.33 ± 1.67 | 13.00 ± 2.89 | 16.29 |
| E. ‘Milk Rose’ | 17.51 ± 0.21 | 15.45 ± 0.15 | 15.67 ± 1.20 | 12.67 ± 1.33 | 14.03 |
| E. ‘Peerless’ | 17.37 ± 0.06 | 18.53 ± 0.09 | 29.67 ± 1.20 | 13.00 ± 0.58 | 24.25 |
| E. ‘Silhouette’ | 24.51 ± 0.17 | 24.39 ± 0.05 | 12.00 ± 2.08 | 12.67 ± 5.90 | 13.48 |
| E. ‘Snow Bunny’ | 16.93 ± 0.07 | 16.70 ± 0.20 | 13.00 ± 1.15 | 8.33 ± 0.88 | 17.89 |
| E. ‘Tippy’ | 26.56 ± 0.20 | 19.45 ± 0.23 | 30.67 ± 2.33 | 13.00 ± 1.53 | 18.04 |
| E. ‘Viyant’ | 17.67 ± 0.25 | 17.20 ± 0.15 | 12.33 ± 1.76 | 6.33 ± 0.33 | 10.00 |
| Cultivar | Moisture Content Parameters | ||
|---|---|---|---|
| Fresh Weight (FW) (g) | Dry Weight (DW) (g) | Moisture Content (%) | |
| E. ‘Benbadis’ | 43.21 ± 1.00 z | 1.53 ± 0.19 | 96.59 ± 0.17 |
| E. ‘Brave’ | 34.86 ± 1.75 | 1.36 ± 0.18 | 96.96 ± 0.12 |
| E. colorata E. Walther | 51.52 ± 0.16 | 2.49 ± 0.26 | 95.57 ± 0.15 |
| E. ‘Cubic Frost’ | 30.00 ± 0.53 | 2.15 ± 0.06 | 92.59 ± 0.29 |
| E. ‘Dark Ice’ | 37.70 ± 0.50 | 1.54 ± 0.08 | 95.54 ± 0.07 |
| E. ‘Doterang’ | 56.36 ± 1.27 | 2.23 ± 0.17 | 95.79 ± 0.06 |
| E. ‘Fire Pillar’ | 66.75 ± 1.24 | 2.60 ± 0.12 | 95.60 ± 0.51 |
| E. ‘Glam Pink’ | 80.63 ± 3.60 | 4.23 ± 0.31 | 94.74 ± 0.13 |
| E. ‘Loy’ | 76.84 ± 8.49 | 4.55 ± 0.75 | 93.69 ± 0.11 |
| E. ‘Milk Rose’ | 115.13 ± 1.78 | 3.40 ± 0.48 | 97.11 ± 0.29 |
| E. ‘Peerless’ | 62.90 ± 3.25 | 3.08 ± 0.16 | 95.09 ± 0.25 |
| E. ‘Silhouette’ | 54.69 ± 0.31 | 2.16 ± 0.11 | 95.77 ± 0.11 |
| E. ‘Snow Bunny’ | 14.40 ± 0.71 | 0.75 ± 0.07 | 95.51 ± 0.22 |
| E. ‘Tippy’ | 70.24 ± 5.19 | 2.61 ± 0.03 | 96.75 ± 0.04 |
| E. ‘Viyant’ | 39.34 ± 0.65 | 1.70 ± 0.15 | 95.22 ± 0.14 |
| Parameters | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Leaf thickness | 1.00 | 0.013 | 0.152 | 0.060 | 0.187 | 0.306 * | 0.536 ** | 0.322 * | 0.205 | −0.287 | 0.117 | −0.042 | 0.366 * | 0.324 * | −0.0.36 |
| 2. Stomata density adaxial | 1.00 | 0.587 ** | 0.482 ** | 0.339 * | 0.221 | 0.312 * | 0.134 | 0.296 * | 0.176 | −0.353 * | −0.181 | 0.321 * | 0.153 | 0.204 | |
| 3. Stomata density abaxial | 1.00 | 0.328 * | 0.376 * | 0.232 | 0.502 ** | 0.149 | 0.402 ** | 0.073 | −0.059 | −0.294 * | 0.363 * | 0.299 * | 0.062 | ||
| 4. Epidermal cell adaxial | 1.00 | 0.759 ** | −0.323 * | −0.064 | −0.166 | −0.160 | 0.357 * | −0.220 | −0.227 | 0.281 | 0.207 | 0.084 | |||
| 5. Epidermal cell abaxial | 1.00 | −0.078 | −0.077 | 0.005 | 0.080 | 0.368 * | −0.241 | −0.161 | 0.307 * | 0.202 | −0.011 | ||||
| 6. Epidermis width | 1.00 | 0.474 ** | 0.362 * | 0.573 ** | −0.332* | −0.245 | −0.178 | 0.148 | 0.056 | 0.110 | |||||
| 7. Epidermis length | 1.00 | 0.527 ** | 0.0655 ** | −0.210 | −0.061 | −0.383 ** | 0.291 | 0.279 | 0.033 | ||||||
| 8. Hypodermis width | 1.00 | 0.729 ** | −0.415 ** | −0.009 | −0.407 ** | 0.386 ** | 0.348 * | −0.075 | |||||||
| 9. Hypodermis length | 1.00 | −0.188 | −0.263 | −0.437 ** | 0.243 | 0.154 | 0.056 | ||||||||
| 10. Leaf shape | 1.00 | −0.171 | 0.022 | −0.193 | −0.255 | 0.092 | |||||||||
| 11. Leaf apex | 1.00 | 0.243 | 0.069 | 0.151 | −0.141 | ||||||||||
| 12. Wax amount | 1.00 | −0.219 | −0.188 | −0.106 | |||||||||||
| 13. Fresh weight | 1.00 | 0.834 ** | 0.024 | ||||||||||||
| 14. Dry weight | 1.00 | −0.469 ** | |||||||||||||
| 15. Moisture content | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha Tran, M.K.T.; Cabahug-Braza, R.A.M.; Hwang, Y.-J. Echeveria Leaf Morpho-Anatomical Analysis and Its Implications for Environmental Stress Conditions. Horticulturae 2024, 10, 308. https://doi.org/10.3390/horticulturae10040308
Ha Tran MKT, Cabahug-Braza RAM, Hwang Y-J. Echeveria Leaf Morpho-Anatomical Analysis and Its Implications for Environmental Stress Conditions. Horticulturae. 2024; 10(4):308. https://doi.org/10.3390/horticulturae10040308
Chicago/Turabian StyleHa Tran, My Khanh Thi, Raisa Aone M. Cabahug-Braza, and Yoon-Jung Hwang. 2024. "Echeveria Leaf Morpho-Anatomical Analysis and Its Implications for Environmental Stress Conditions" Horticulturae 10, no. 4: 308. https://doi.org/10.3390/horticulturae10040308
APA StyleHa Tran, M. K. T., Cabahug-Braza, R. A. M., & Hwang, Y.-J. (2024). Echeveria Leaf Morpho-Anatomical Analysis and Its Implications for Environmental Stress Conditions. Horticulturae, 10(4), 308. https://doi.org/10.3390/horticulturae10040308