
Synthetic Seed Production and Slow Growth Storage of In Vitro Cultured Plants of Iris pallida Lam.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
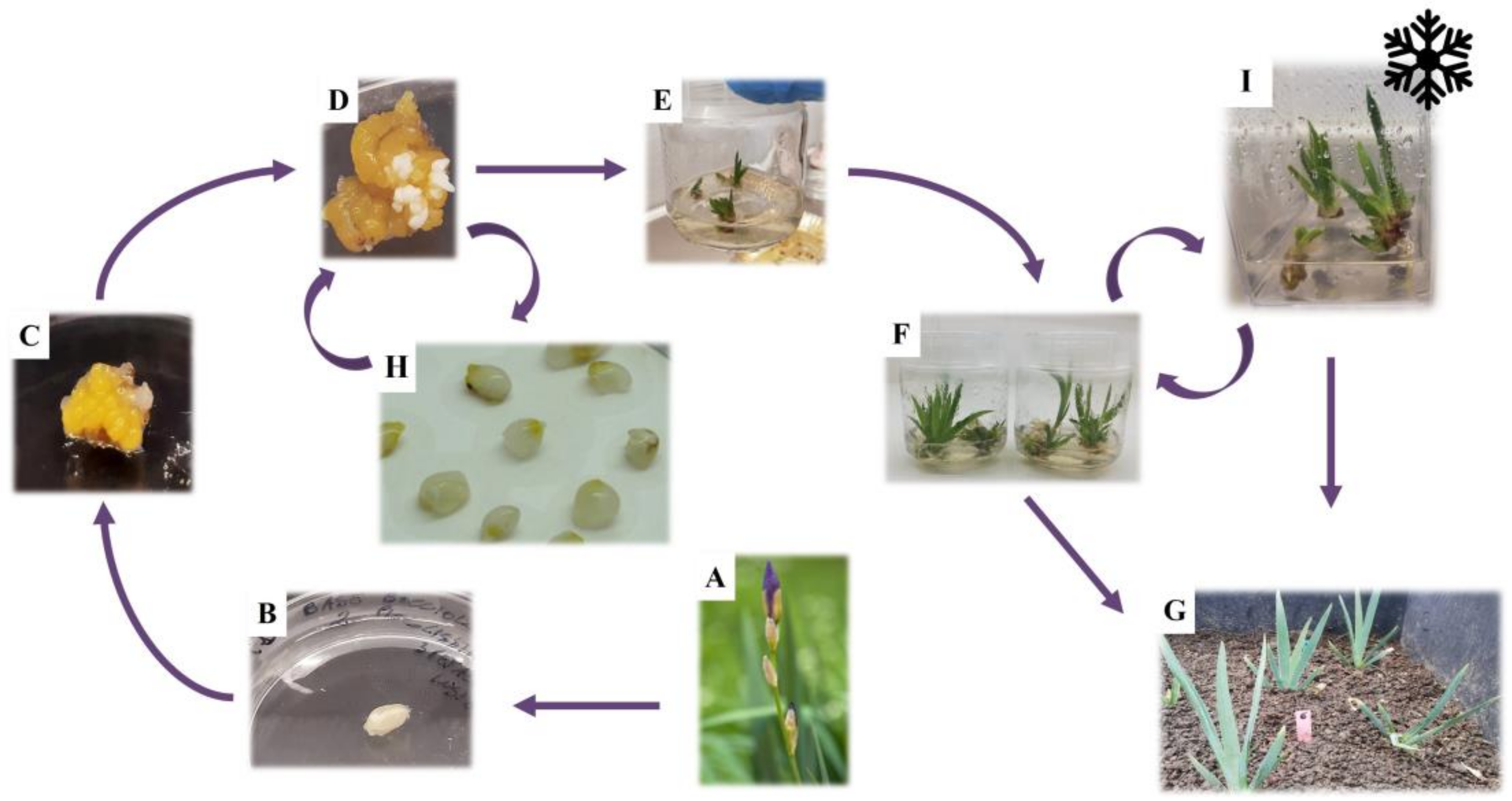
2.1. Induction of Somatic Embryos and Plantlet Regeneration
2.2. Slow Growth Storage (SGS)
Morphological and Biochemical Determinations
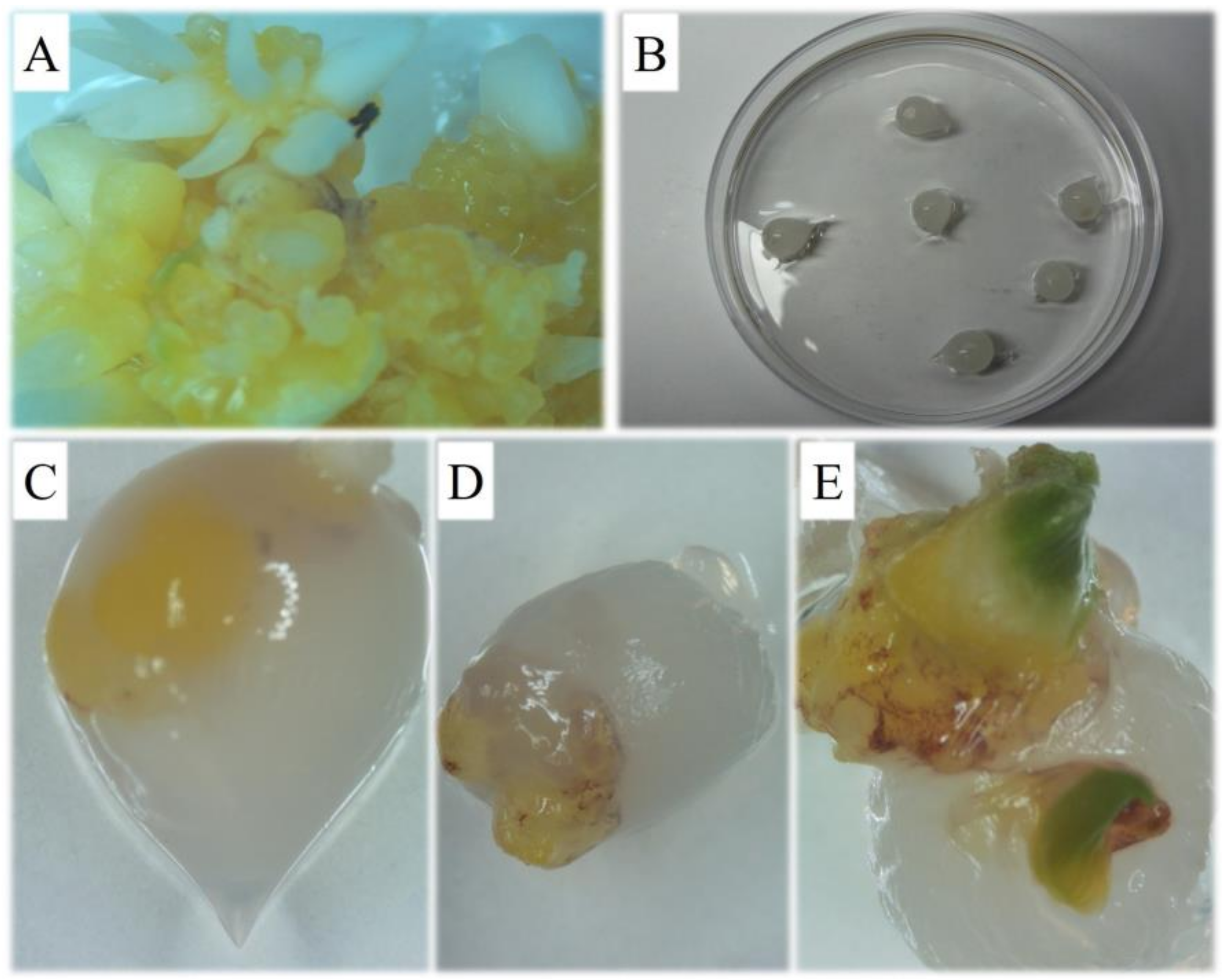
2.3. Synthetic Seeds Production and Storage
- (A)
- Fourteen days of storage at a cold temperature of 4 °C in a laboratory refrigerator;
- (B)
- Fourteen days of storage at a room temperature of 23 °C;
- (C)
- Twenty-eight days of storage at a cold temperature of 4 °C in a laboratory refrigerator;
- (D)
- Twenty-eight days of storage at a room temperature of 23 °C;
2.4. Statistical Analysis
3. Results
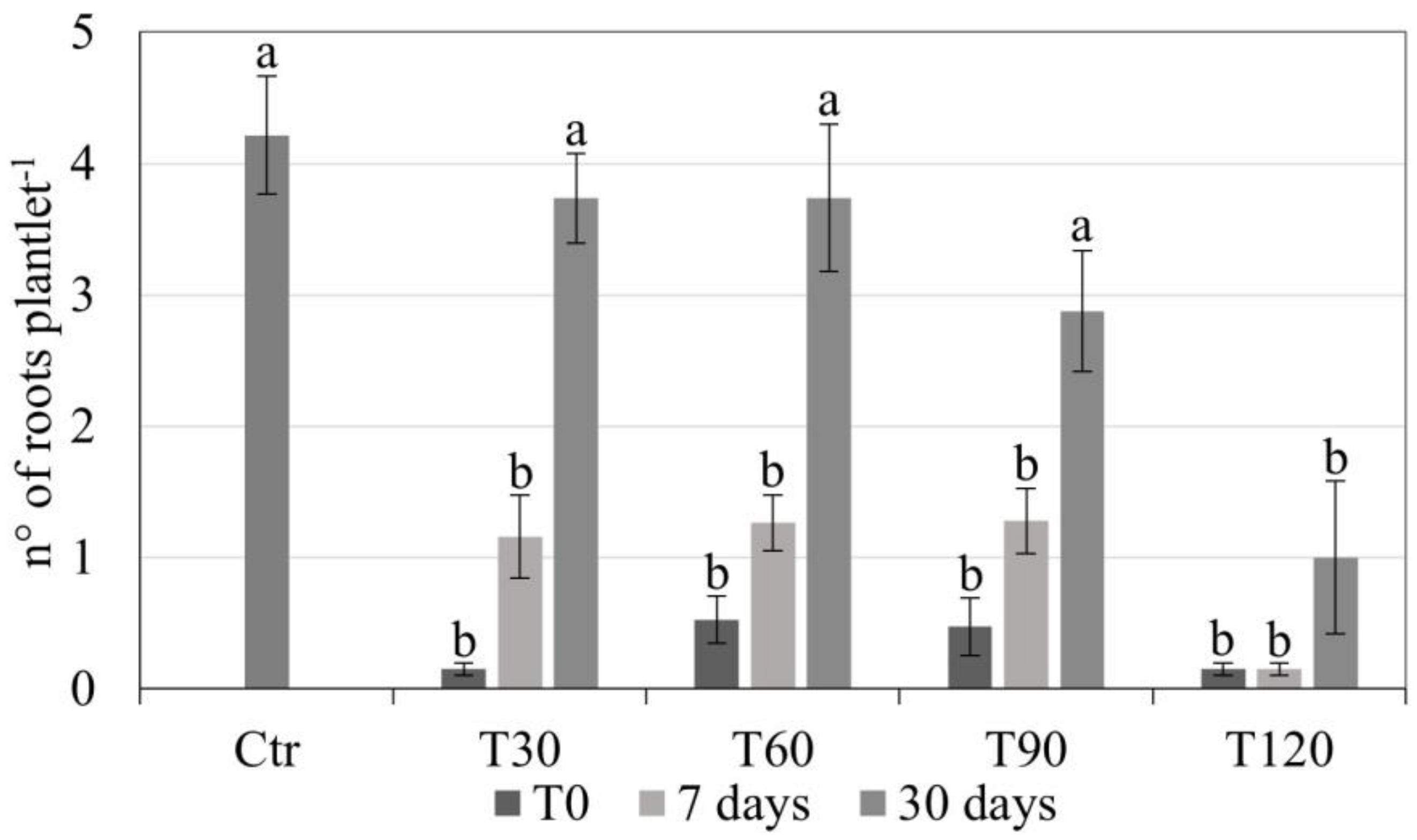
3.1. Slow Growth Storage (SGS)
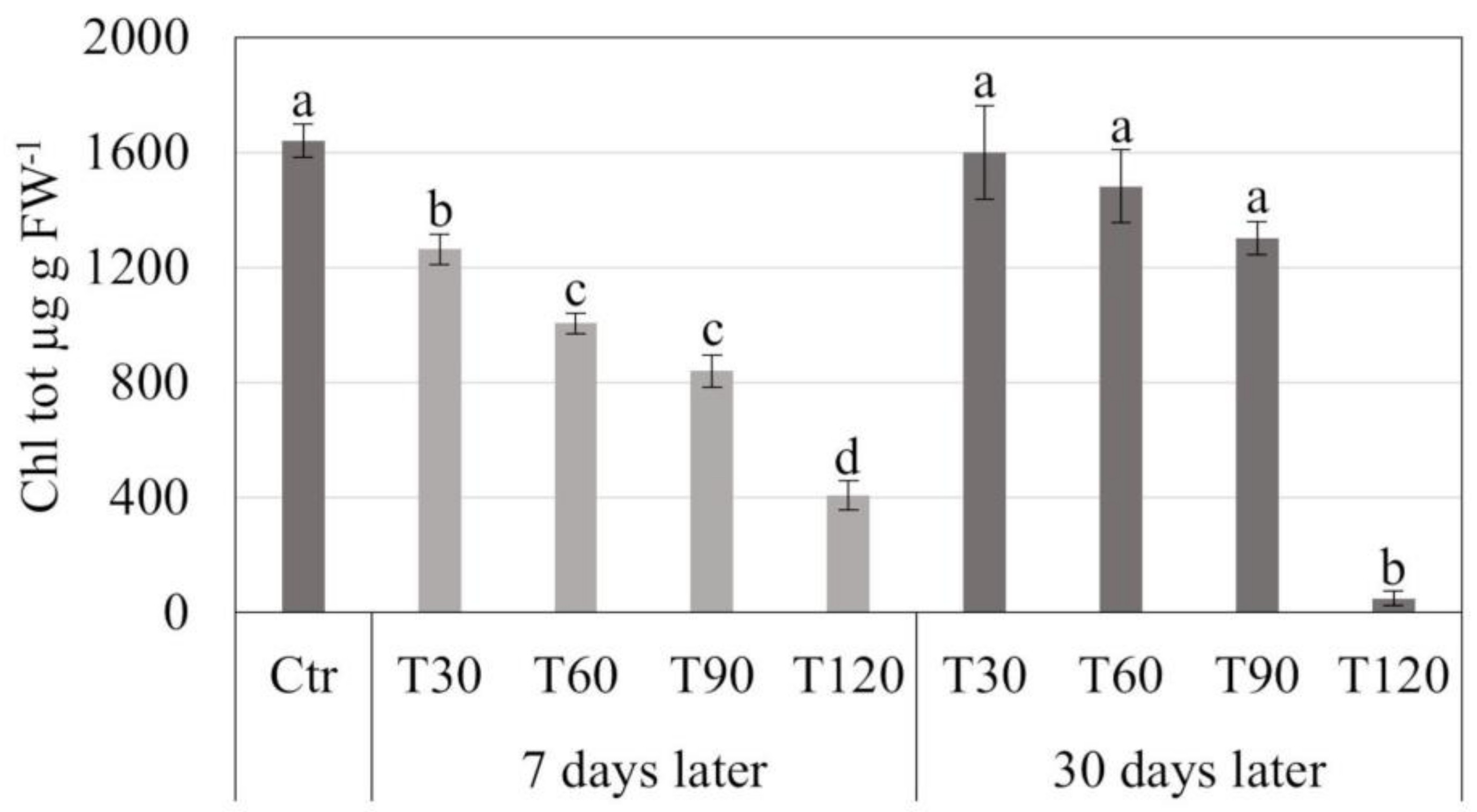
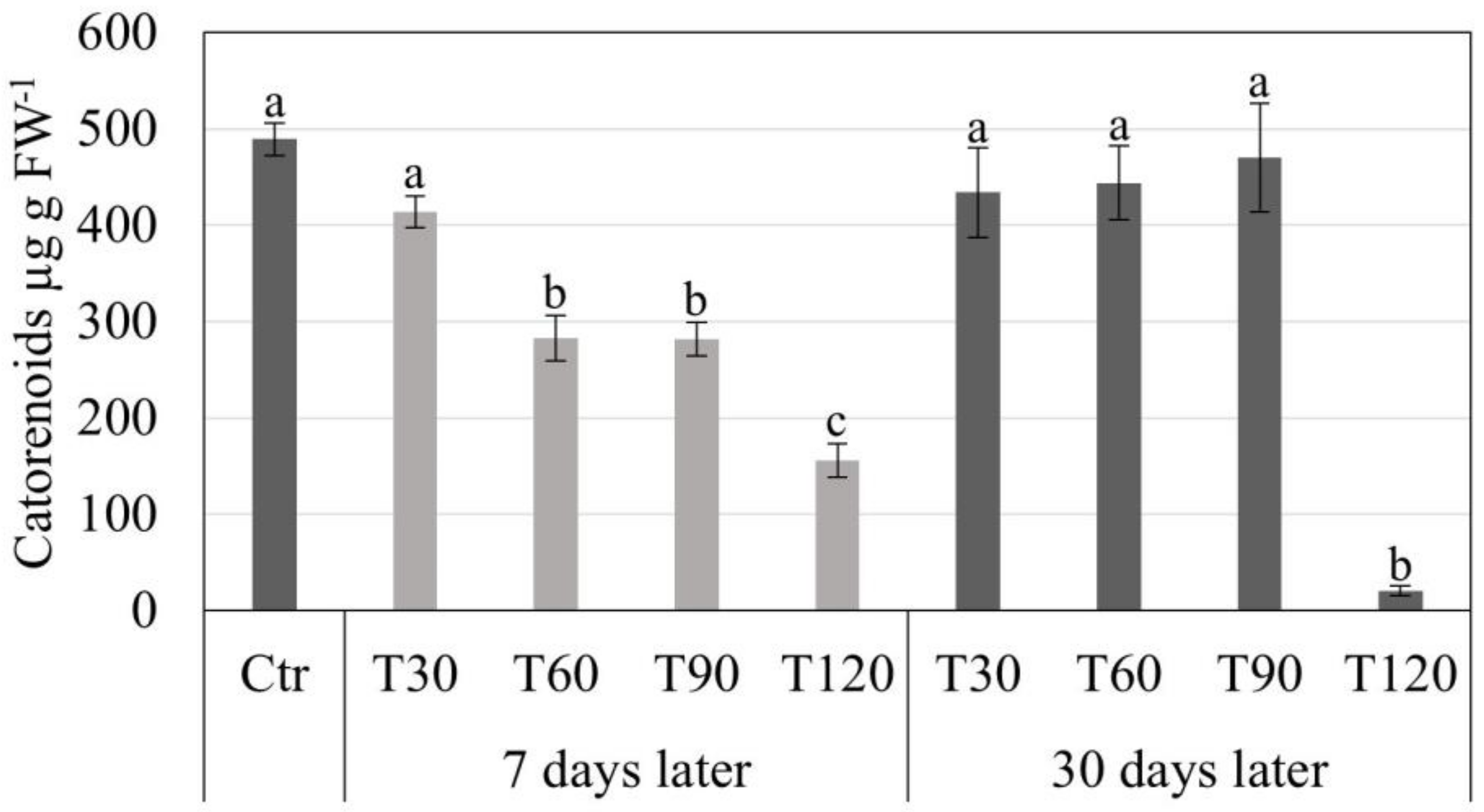
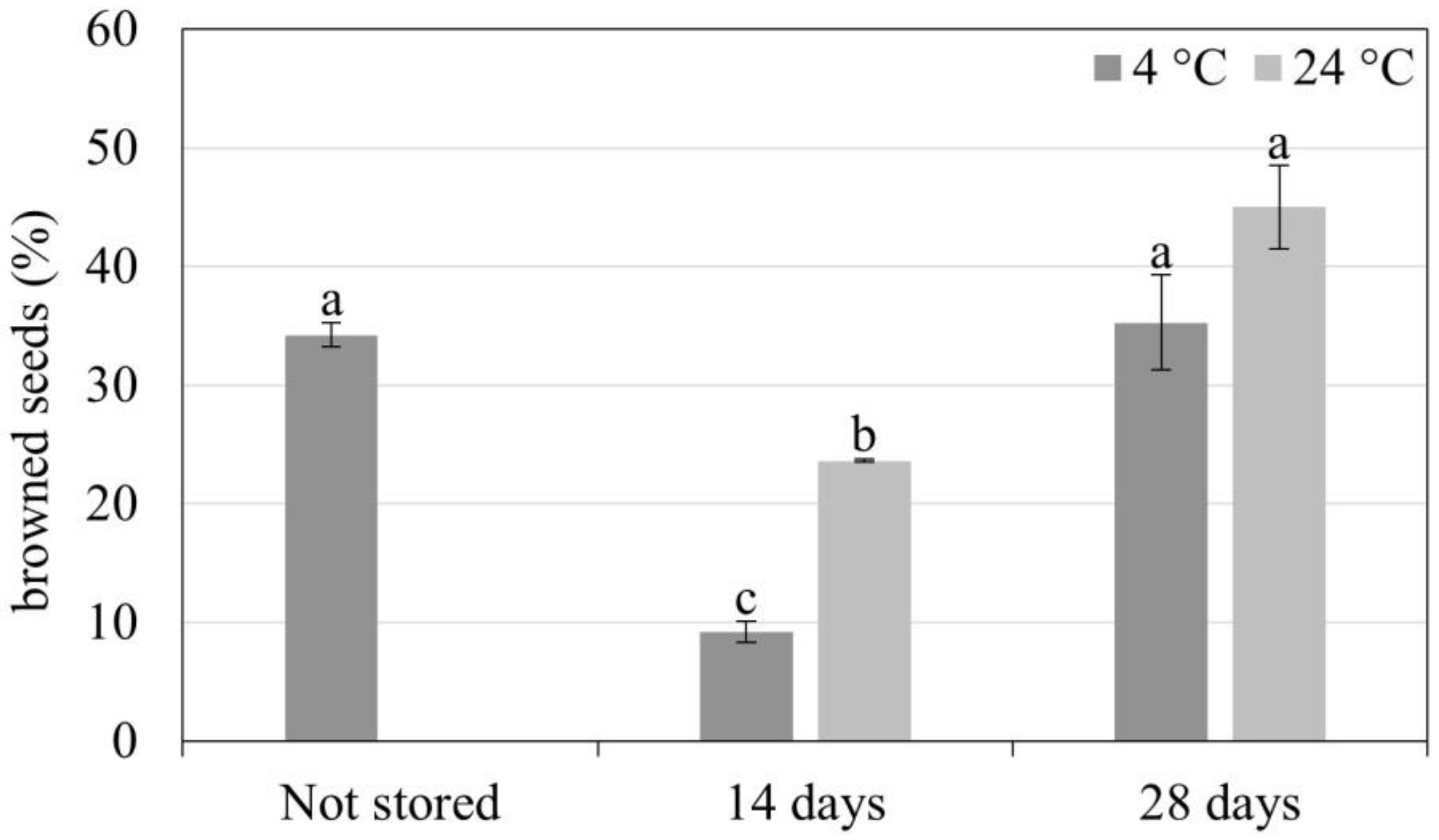
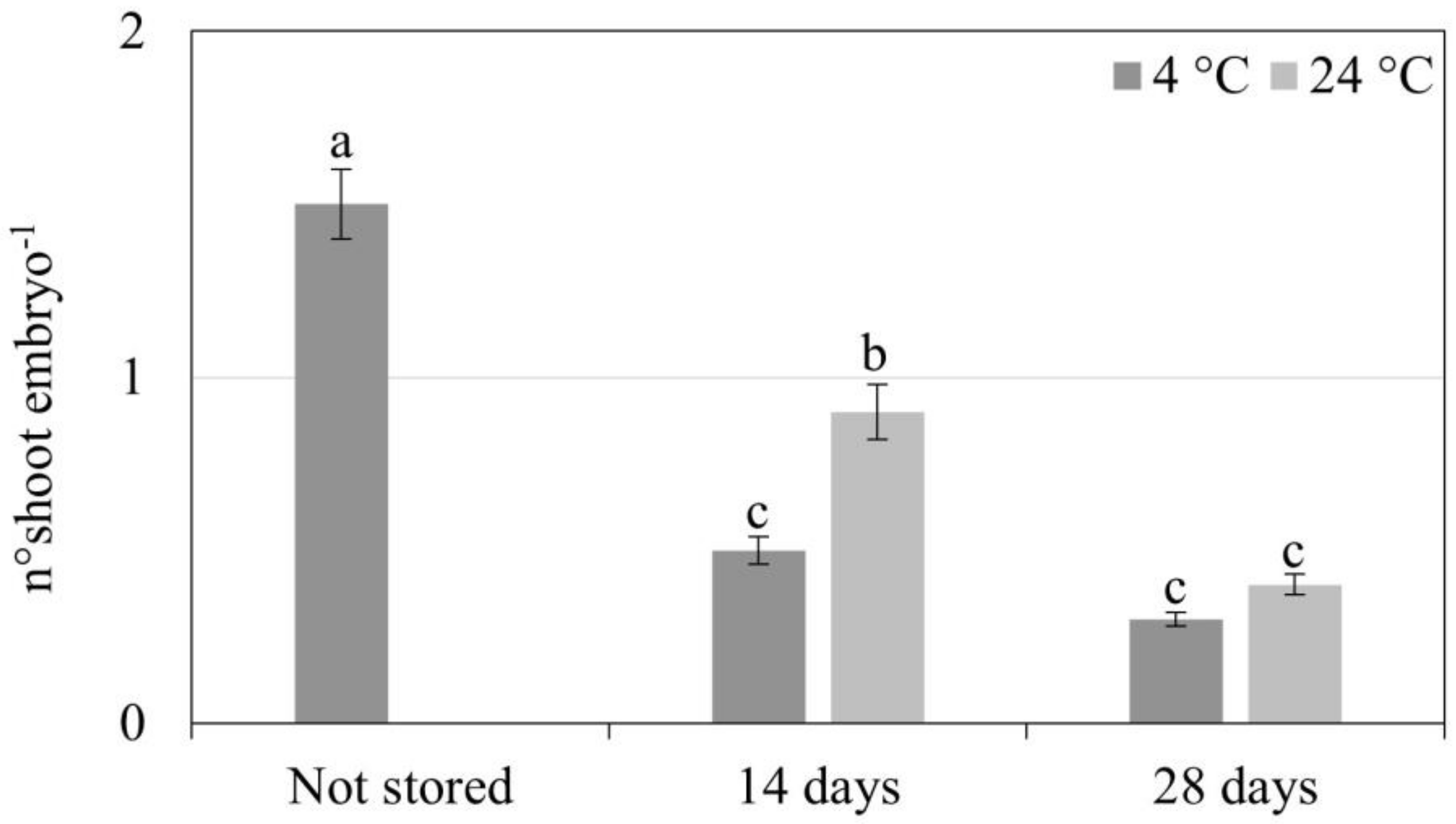
3.2. Synthetic Seed Production and Storage
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Bonneval, E.; David, O.R.P.; Doré, J.; Hojlo, A.S.; Inrig, W.; Muller, C.; de Swardt, D. Iris: L’iris en Parfumerie; Coédition Nez éditions/Laboratoire Monique Rémy: San Diego, CA, USA, 2020; p. 96. [Google Scholar]
- Khatib, S.; Faraloni, C.; Bouissane, L. Exploring the Use of Iris Species: Antioxidant Properties, Phytochemistry, Medicinal and Industrial Applications. Antioxidants 2022, 11, 526. [Google Scholar] [CrossRef] [PubMed]
- Jaenicke, L.; Marner, F.J. The irones and their origin. Pure Appl. Chem. 1990, 62, 1365–1368. [Google Scholar] [CrossRef]
- Bicchi, C.; Rubiolo, P. Analysis of constituents of iris rhizomes. Part I: High performance liquid chromatographic-particle beam-mass spectral analysis of iridals from Iris pallida rhizomes. Phytochem. Anal. 1993, 4, 171–177. [Google Scholar] [CrossRef]
- Ascough, G.D.; Erwin, J.E.; Van Staden, J. Micropropagation of Iridaceae—A review. Plant Cell Tissue Organ Cult. (PCTOC) 2009, 97, 1–19. [Google Scholar] [CrossRef]
- Lucchesini, M.; Bedini, L.; Florio, E.F.; Maggini, R.; Malorgio, F.; Pezzarossa, B.; Mensuali-Sodi, A. The improvement of Iris pallida propagation by somatic embryogenesis. Acta Hortic. 2017, 1155, 127–134. [Google Scholar] [CrossRef]
- Alzubi, H.; Yepes, L.M.; Fuchs, M. In vitro storage of micropropagated grapevine rootstocks at low temperature. In Vitro Cell. Dev. Biol.-Plant 2019, 55, 334–341. [Google Scholar] [CrossRef]
- Hatzilazarou, S.; Kostas, S.; Joachim, M.; Economou, A. Regeneration of Viburnum dentatum L. from alginate-encapsulated shoot explants after short-term cold storage and assessment of genetic stability using ISSR analysis. Agronomy 2020, 10, 1660. [Google Scholar] [CrossRef]
- Manokari, M.; Latha, R.; Privadharshini, S.; Jogam, P.; Shekhawat, M.S. Short-term cold storage of encapsulated somatic embryos and retrieval of plantlets in grey orchid (Vanda tessellata (Roxb.) Hook. ex G. Don). Plant Cell Tissue Organ Cult. (PCTOC) 2021, 144, 171–183. [Google Scholar] [CrossRef]
- Martinez, M.T.; Arrillaga, I.; Sales, E.; Perez-Oliver, M.A.; Gonzalez-Mas, M.D.C.; Corredoira, E. Micropropagation, characterization, and conservation of Phytophthora cinnamomi-tolerant holm oak mature trees. Forests 2021, 12, 1634. [Google Scholar] [CrossRef]
- Qahtan, A.A.; Abdel-Salam, E.M.; Alatar, A.A.; Wang, Q.C.; Faisal, M. An Introduction to Synthetic Seeds: Production, Techniques, and Applications. In Synthetic Seeds; Faisal, M., Alatar, A., Eds.; Springer: Cham, Switzerland, 2019; pp. 1–20. [Google Scholar]
- Maqsood, M.; Khusrau, M.; Mujib, A.; Kaloo, Z.A. Synthetic seed technology in some ornamental and medicinal plants: An overview. In Propagation and Genetic Manipulation of Plants; Springer: Singapore, 2021; pp. 19–31. [Google Scholar]
- Garg, R.; Maheshwari, S. Synthetic seed technology, application and future trends. EPH-Int. J. Agric. Environ. Res. 2023, 9, 1–10. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Engelmann, F. Use of biotechnologies for the conservation of plant biodiversity. In Vitro Cell. Dev. Biol. Plant 2011, 47, 5–16. [Google Scholar] [CrossRef]
- Benelli, C.; Ozudogru, E.A.; Lambardi, M.; Dradi, G. In vitro conservation of ornamental plants by slow growth storage. In Proceedings of the VII International Symposium on In Vitro Culture and Horticultural Breeding, Ghent, Belgium, 18–22 September 2011; Volume 961, pp. 89–93. [Google Scholar]
- Pezzarossa, B.; Bretzel, F.; Malorgio, F.; Borghesi, E.; Maggini, R.; Scaramuzzi, S. La coltivazione del Giaggiolo in Toscana: Dalla marginalità alla valorizzazione. Colt. Protette 2016, 2016, 7–8. [Google Scholar]
- Fürtauer, L.; Weiszmann, J.; Weckwerth, W.; Nägele, T. Dynamics of Plant Metabolism during Cold Acclimation. Int. J. Mol. Sci. 2019, 20, 5411. [Google Scholar] [CrossRef]
- Lambardi, M.; Ozudogru, E.A.; Jain, S.M. (Eds.) Protocols for Micropropagation of Selected Economically-Important Horticultural Plants; Humana Press: Hertfordshire, UK, 2013; Volume 994, p. 490. [Google Scholar]
- Benson, E.E.; Harding, K.; Debouck, D.; Dumet, D.; Escobar, R.; Mafla, G.; Panis, B.; Panta, A.; Tay, D.; Houwe Van den, I.; et al. Refinement and Standardization of Storage Procedures for Clonal Crops. Global Public Goods Phase 2: Part 1. Project Landscape and General Status of Clonal Crop In Vitro Conservation Technologies; System-Wide Genetic Resources Programme (SGRP): Rome, Italy, 2011; p. 86. ISBN 978-92-9043-905-9. [Google Scholar]
- Ruta, C.; Lambardi, M.; Ozudogru, E.A. Biobanking of vegetable genetic resources by in vitro conservation and cryopreservation. Biodivers. Conserv. 2020, 29, 3495–3532. [Google Scholar] [CrossRef]
- Cha-um, S.; Kirdmanee, C. Minimal growth in vitro culture for preservation of plant species. Fruit Veg. Cereal Sci. Biotechnol. 2007, 1, 13–25. [Google Scholar]
- Lucchesini, M.; Mensuali-Sodi, A.; Massai, R.; Gucci, R. Development of Autotrophy and Tolerance to Acclimatization of Myrtus Communis Transplants Cultured In Vitro under Different Aeration. Biol. Plant. 2001, 44, 167–174. [Google Scholar] [CrossRef]
- Lucchesini, M.; Pacifici, S.; Maggini, R.; Pardossi, A.; Mensuali, A. A novel microfloating culture system for the in vitro rooting of Echinacea angustifolia D.C.: Photosynthetic performance and production of caffeic acid derivatives. Plant Cell Tissue Organ Cult. (PCTOC) 2019, 136, 123–132. [Google Scholar] [CrossRef]
- Kiferle, C.; Lucchesini, M.; Maggini, R.; Pardossi, A.; Mensuali-Sodi, A. In vitro culture of sweet basil: Gas exchanges, growth, and rosmarinic acid production. Biol. Plant. 2014, 58, 601–610. [Google Scholar] [CrossRef]
- Polivanova, O.B.; Bedarev, V.A. Hyperhydricity in Plant Tissue Culture. Plants 2022, 11, 3313. [Google Scholar] [CrossRef]
- Danso, K.E.; Ford-Lloyd, B.V. Encapsulation of nodal cuttings and shoot tips for storage and exchange of cassava germplasm. Plant Cell Rep. 2003, 21, 718–725. [Google Scholar] [CrossRef]
- Hung, C.D.; Trueman, S.J. Encapsulation technology for short-term preservation and germplasm distribution of the African mahogany Khaya senegalensis. Plant Cell Tissue Organ Cult. (PCTOC) 2011, 107, 397–405. [Google Scholar] [CrossRef]
- Standardi, A.; Micheli, M. Encapsulation of in vitro-derived explants: An innovative tool for nurseries. In Protocols for Micropropagation of Selected Economically-Important Horticultural Plants; Humana Press: Totowa, NJ, USA, 2013; pp. 397–418. [Google Scholar]
- Devi, P.; Rathor, S.; Sharma, P.; Sen, J.; Kaur, H.; Singh, J. Development of novel gastroretentive salbutamol sulfate-loaded sodium alginate-pectin bubble beads prepared by co-axial needle air-injection method and in vivo clinical evaluation by ultrasound studies. Eur. J. Pharm. Sci. 2018, 122, 359–373. [Google Scholar] [CrossRef]
- Saadat, S.; Majd, A.; Naseri, L.; Iranbakhsh, A.; Jafari, M. Optimization of somatic embryogenesis, synthetic seed production, and evaluation of genetic fidelity in Teucrium polium L. In Vitro Cell. Dev. Biol.-Plant 2023, 59, 483–496. [Google Scholar] [CrossRef]
- Ahmed, M.R.; Anis, M.; Al-Etta, H.A. Encapsulation technology for short-term storage and germplasm exchange of Vitex trifolia L. Rendiconti Lincei 2015, 26, 133–139. [Google Scholar] [CrossRef]
- Ara, H.; Jaiswal, U.; Jaiswal, V.S. Synthetic seed: Prospects and limitations. Curr. Sci. 2000, 78, 1438–1444. [Google Scholar]
- Khilwani, B.; Kaur, A.; Ranjan, R.; Kumar, A. Direct somatic embryogenesis and encapsulation of somatic embryos for in vitro conservation of Bacopa monnieri (L.) Wettst. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 127, 433–442. [Google Scholar] [CrossRef]
- Shilpha, J.; Pandian, S.; Largia, M.J.V.; Sohn, S.I.; Ramesh, M. Short-term storage of Solanum trilobatum L. synthetic seeds and evaluation of genetic homogeneity using SCoT markers. Plant Biotechnol. Rep. 2021, 15, 651–661. [Google Scholar] [CrossRef]
- Maqsood, M.; Mujib, A.; Khusrau, M. Preparation and low temperature short-term storage for synthetic seeds of Caladium bicolor. Not. Sci. Biol. 2015, 7, 90–95. [Google Scholar] [CrossRef]
- Ikhlaq, M.; Hafiz, I.A.; Micheli, M.; Ahmad, T.; Abbasi, N.A.; Standardi, A. In vitro storage of synthetic seeds: Effect of different storage conditions and intervals on their conversion ability. Afr. J. Biotechnol. 2010, 9, 5712–5721. [Google Scholar]
- Micheli, M.; Hafiz, I.A.; Bazzurri, N.; Standardi, A. Methodological development for the synthetic seeds production of Moraiolo. Olive Bioteq. 2006, 1, 155–158. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meucci, A.; Ghelardi, C.; Chietera, G.; Mensuali, A. Synthetic Seed Production and Slow Growth Storage of In Vitro Cultured Plants of Iris pallida Lam. Horticulturae 2024, 10, 272. https://doi.org/10.3390/horticulturae10030272
Meucci A, Ghelardi C, Chietera G, Mensuali A. Synthetic Seed Production and Slow Growth Storage of In Vitro Cultured Plants of Iris pallida Lam. Horticulturae. 2024; 10(3):272. https://doi.org/10.3390/horticulturae10030272
Chicago/Turabian StyleMeucci, Annalisa, Cristina Ghelardi, Giorgiana Chietera, and Anna Mensuali. 2024. "Synthetic Seed Production and Slow Growth Storage of In Vitro Cultured Plants of Iris pallida Lam." Horticulturae 10, no. 3: 272. https://doi.org/10.3390/horticulturae10030272
APA StyleMeucci, A., Ghelardi, C., Chietera, G., & Mensuali, A. (2024). Synthetic Seed Production and Slow Growth Storage of In Vitro Cultured Plants of Iris pallida Lam. Horticulturae, 10(3), 272. https://doi.org/10.3390/horticulturae10030272