
Effect of Temperature on Photosynthetic Pigment Degradation during Freeze–Thaw Process of Postharvest of Celery Leaves


 ,
, 
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Grouping Design and Treatment
2.3. Celery Sample Collection and Freezing Storage Extraction
2.4. Extraction and Determination of Photosynthetic Pigment Content
2.5. Data Analysis
3. Results
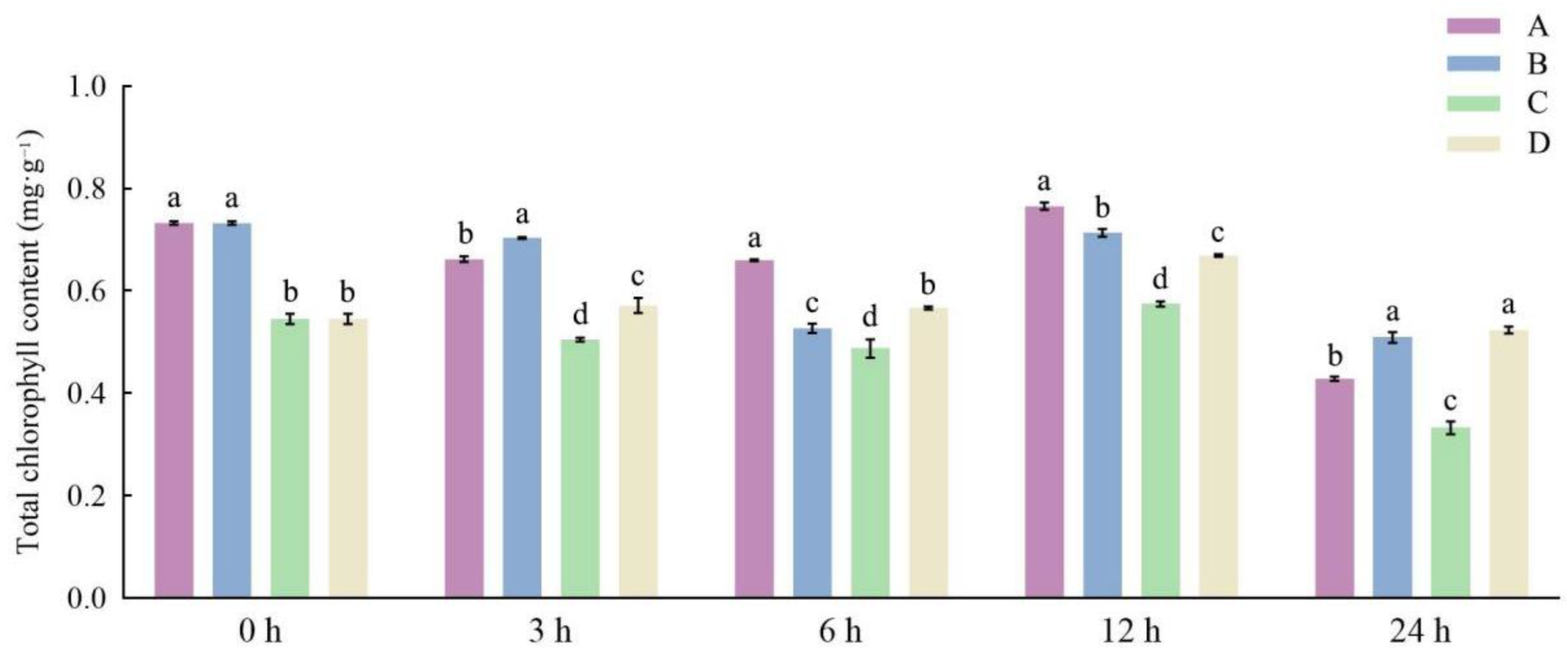
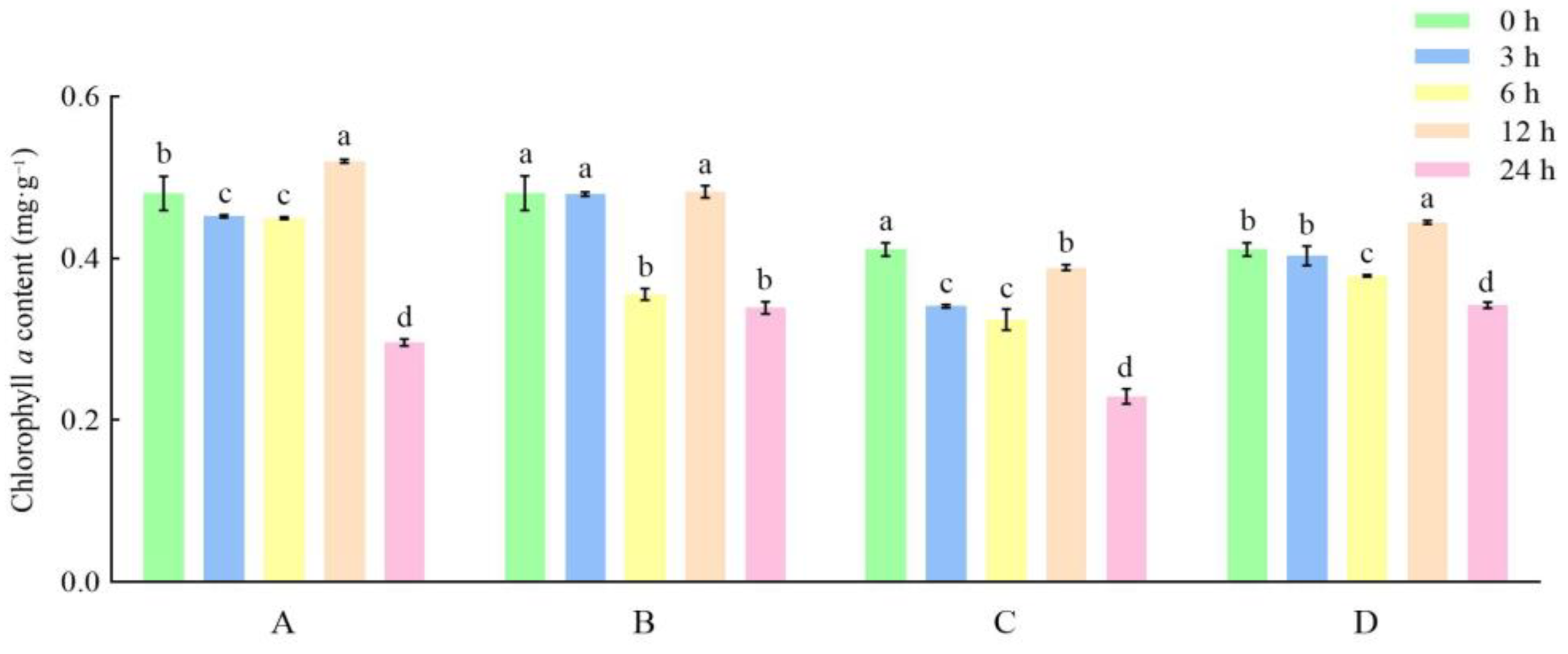
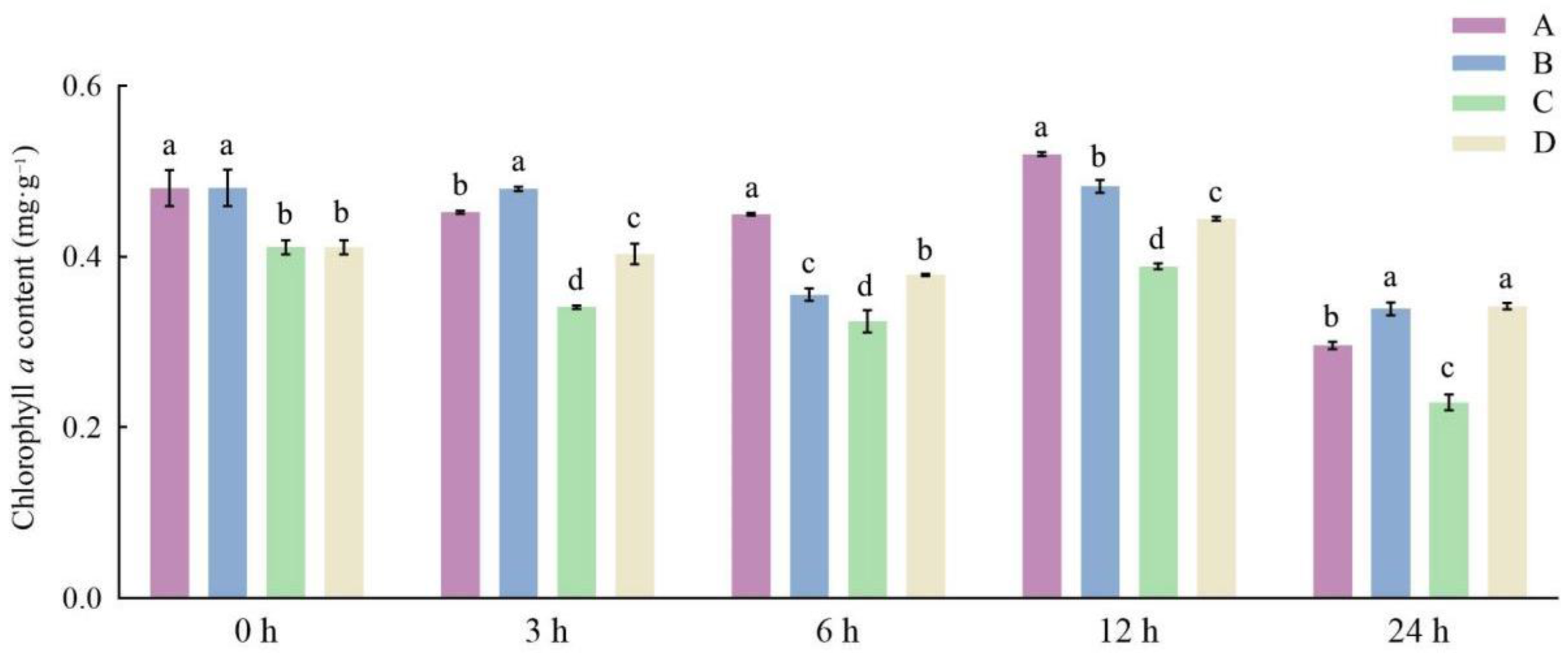
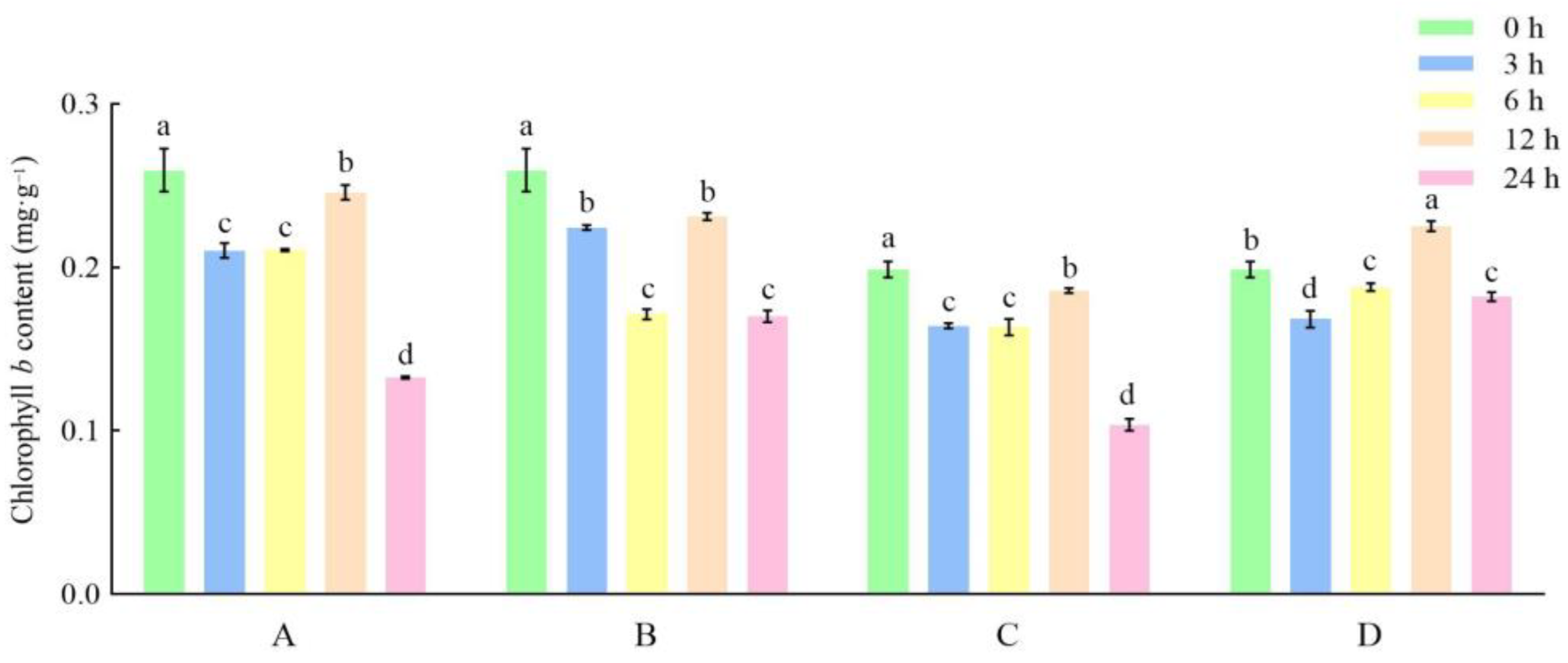
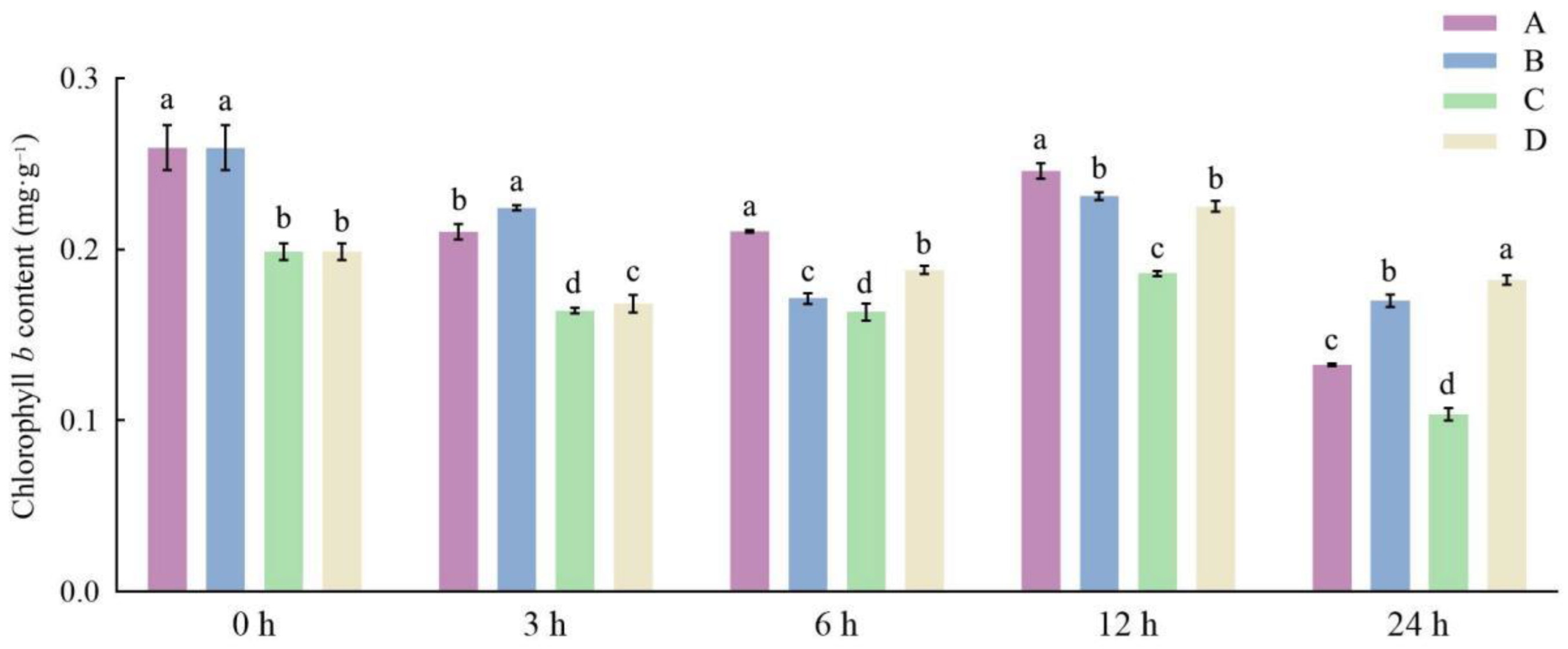
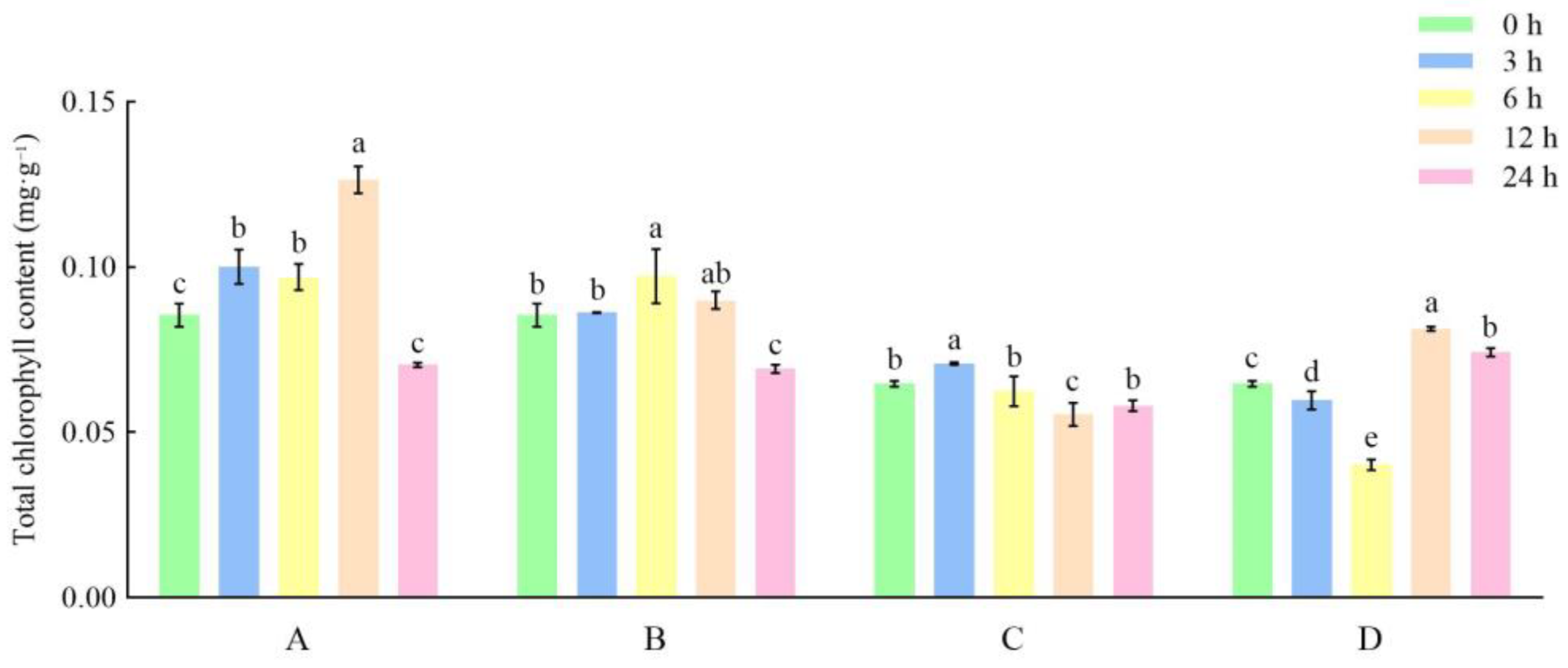
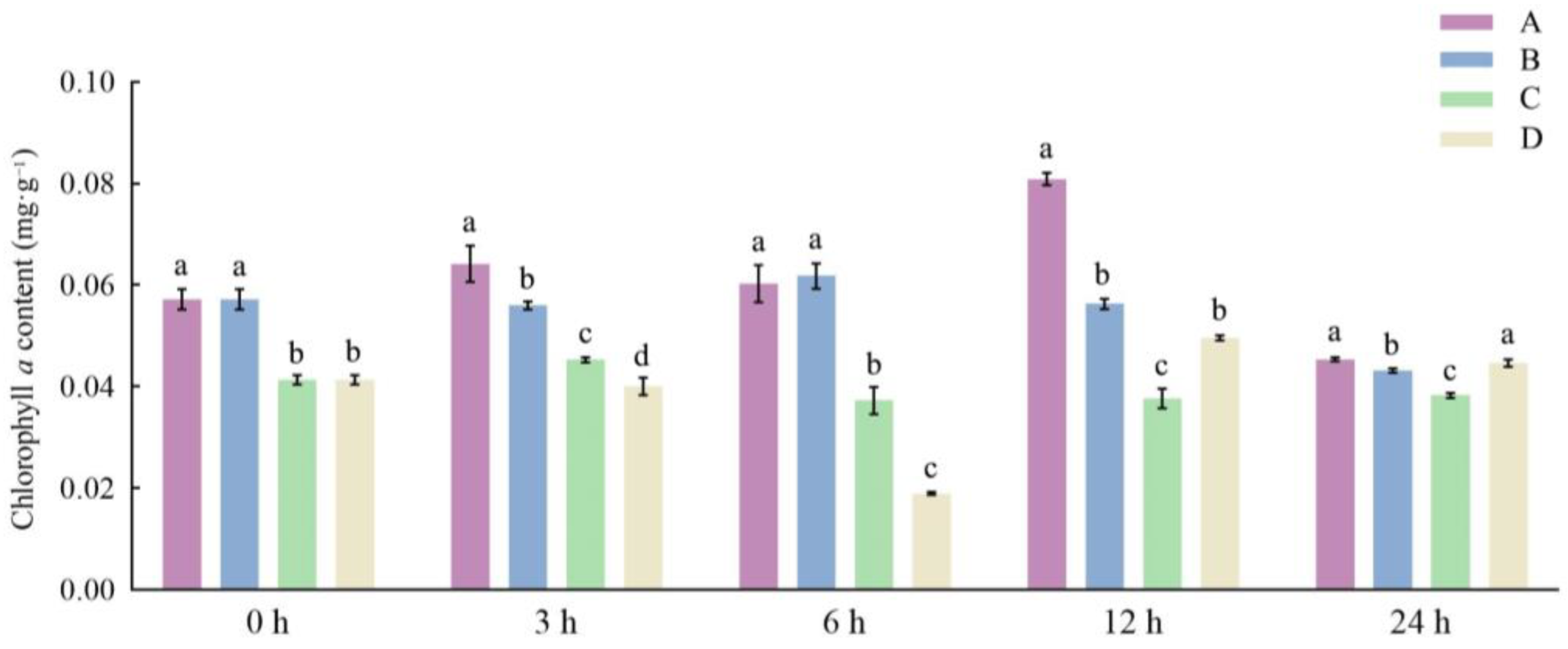
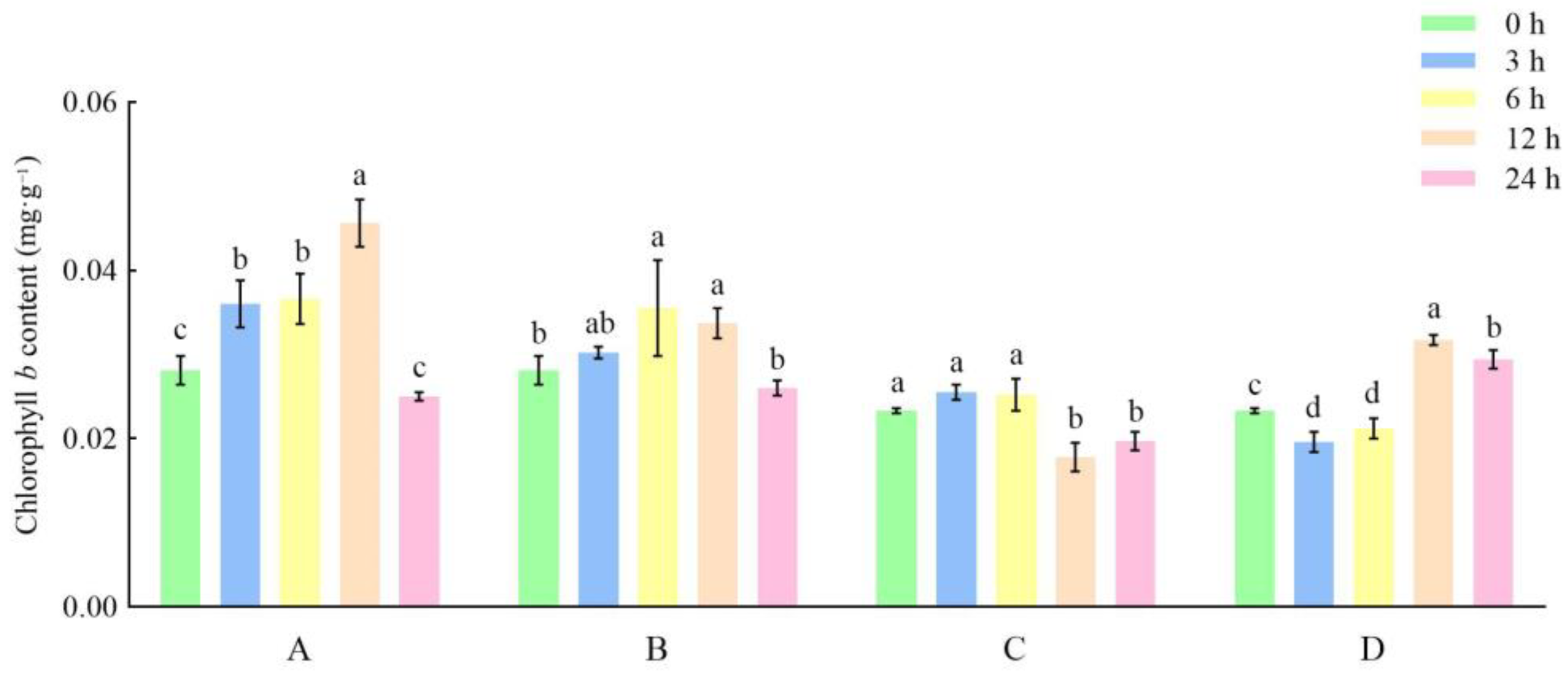
3.1. Effects of Different Storage and Thawing Temperatures on Chlorophyll Content of Celery Leaf Blades during Thawing
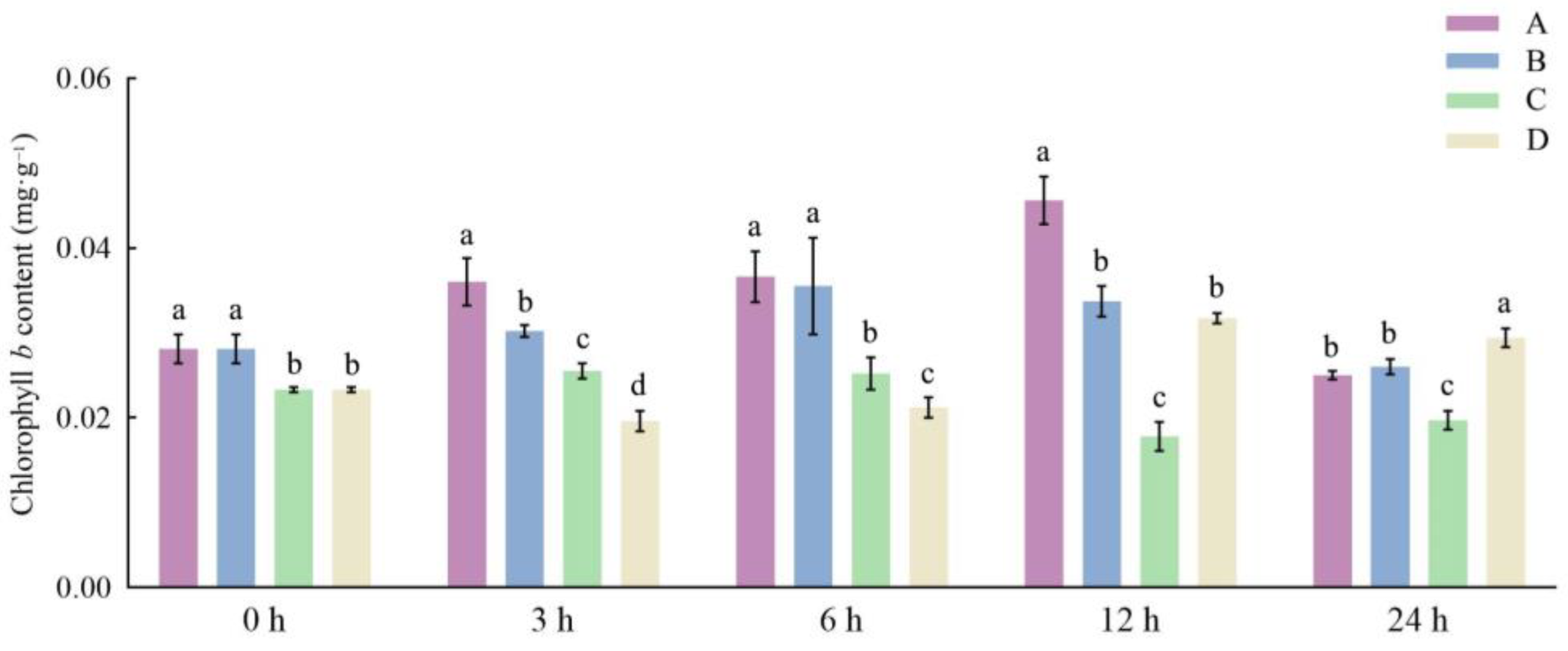
3.2. Effects of Different Storage and Thawing Temperatures on Chlorophyll Content of Celery Petioles during Thawing
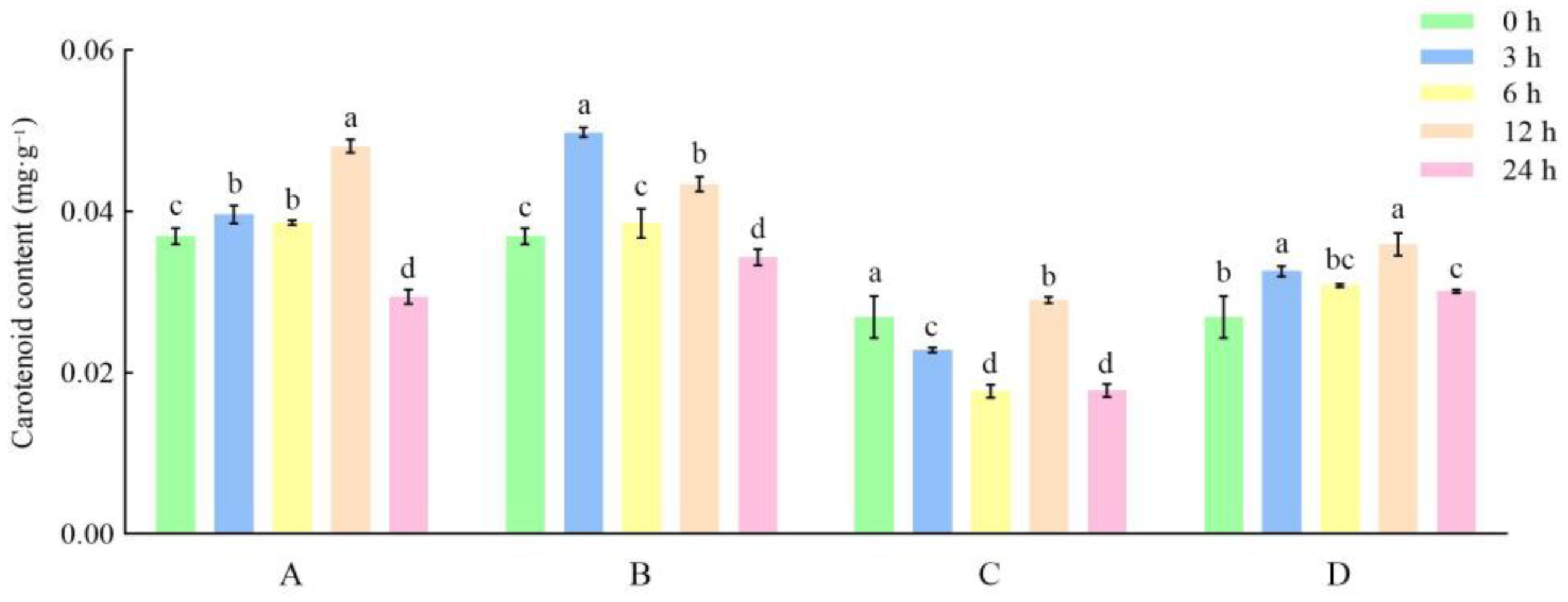
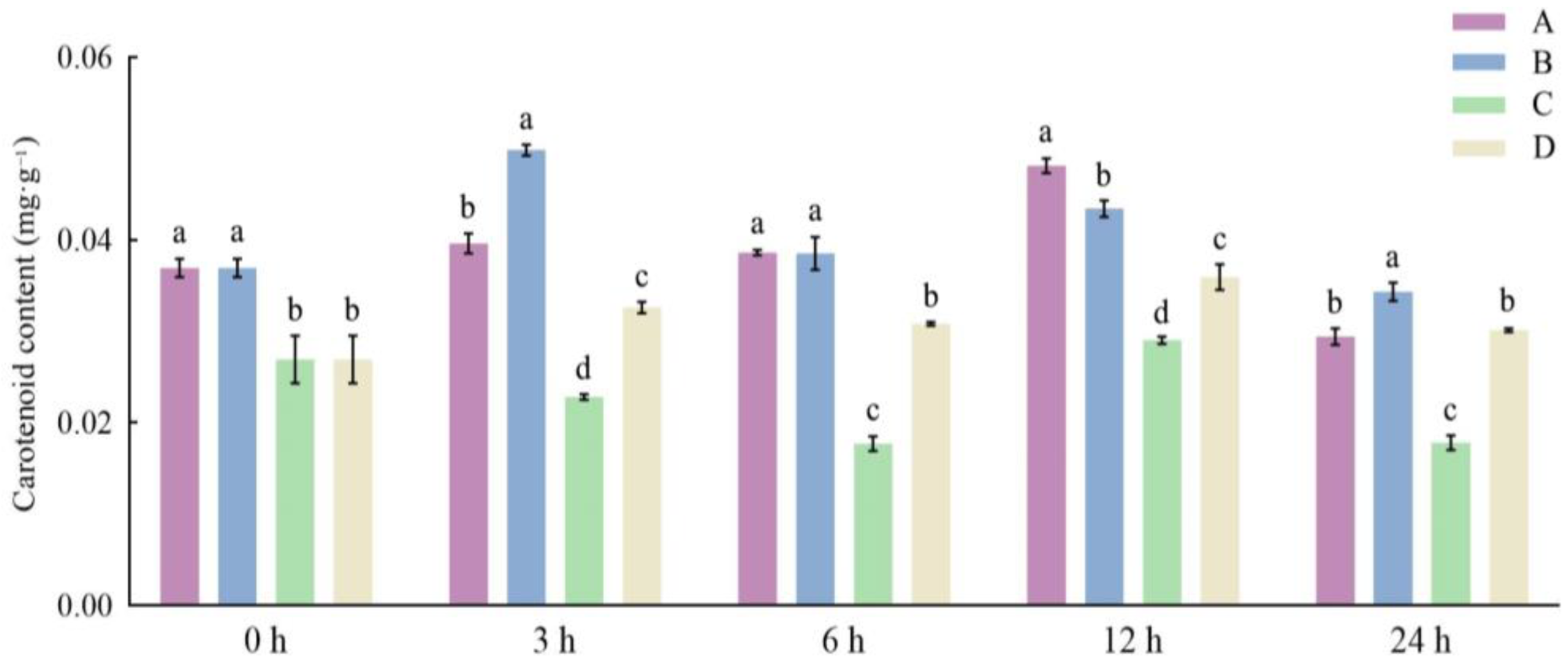
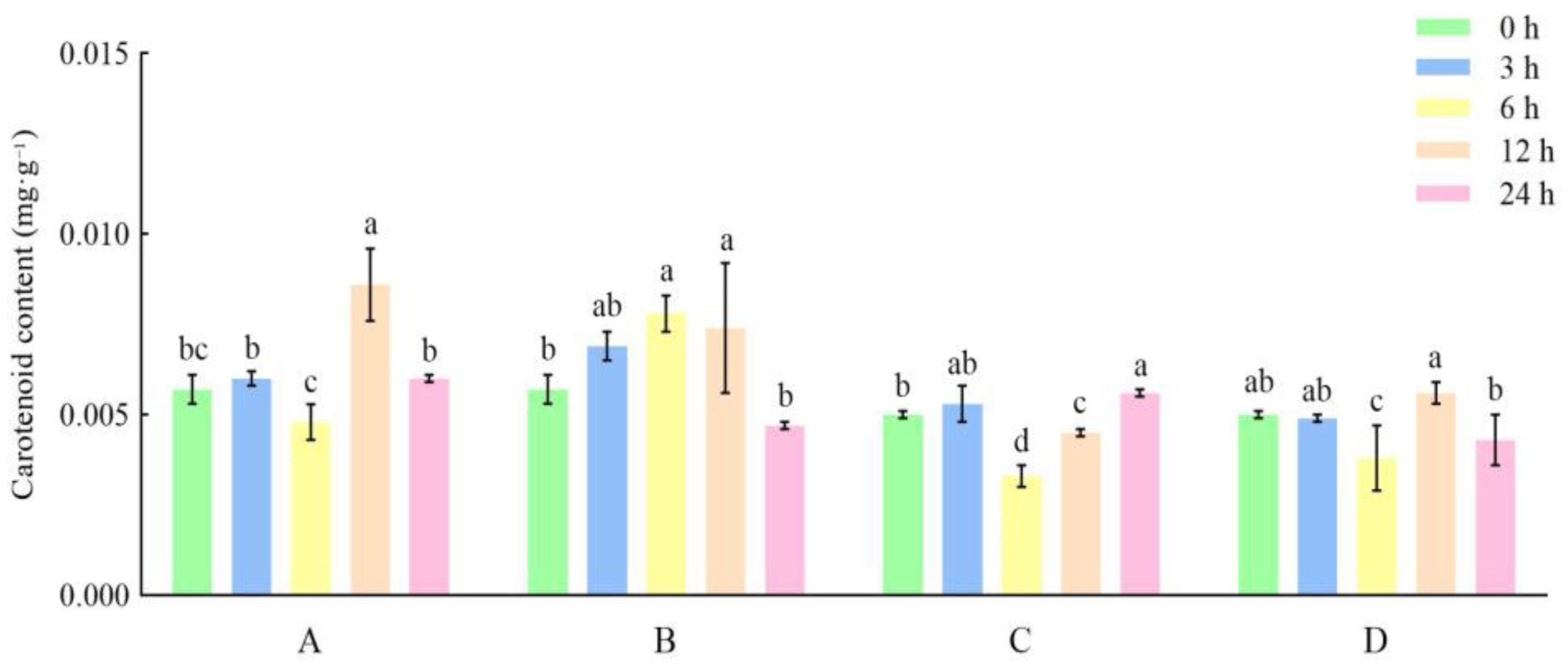
3.3. Effects of Different Storage and Thawing Temperatures on Carotenoid Content of Celery during Thawing
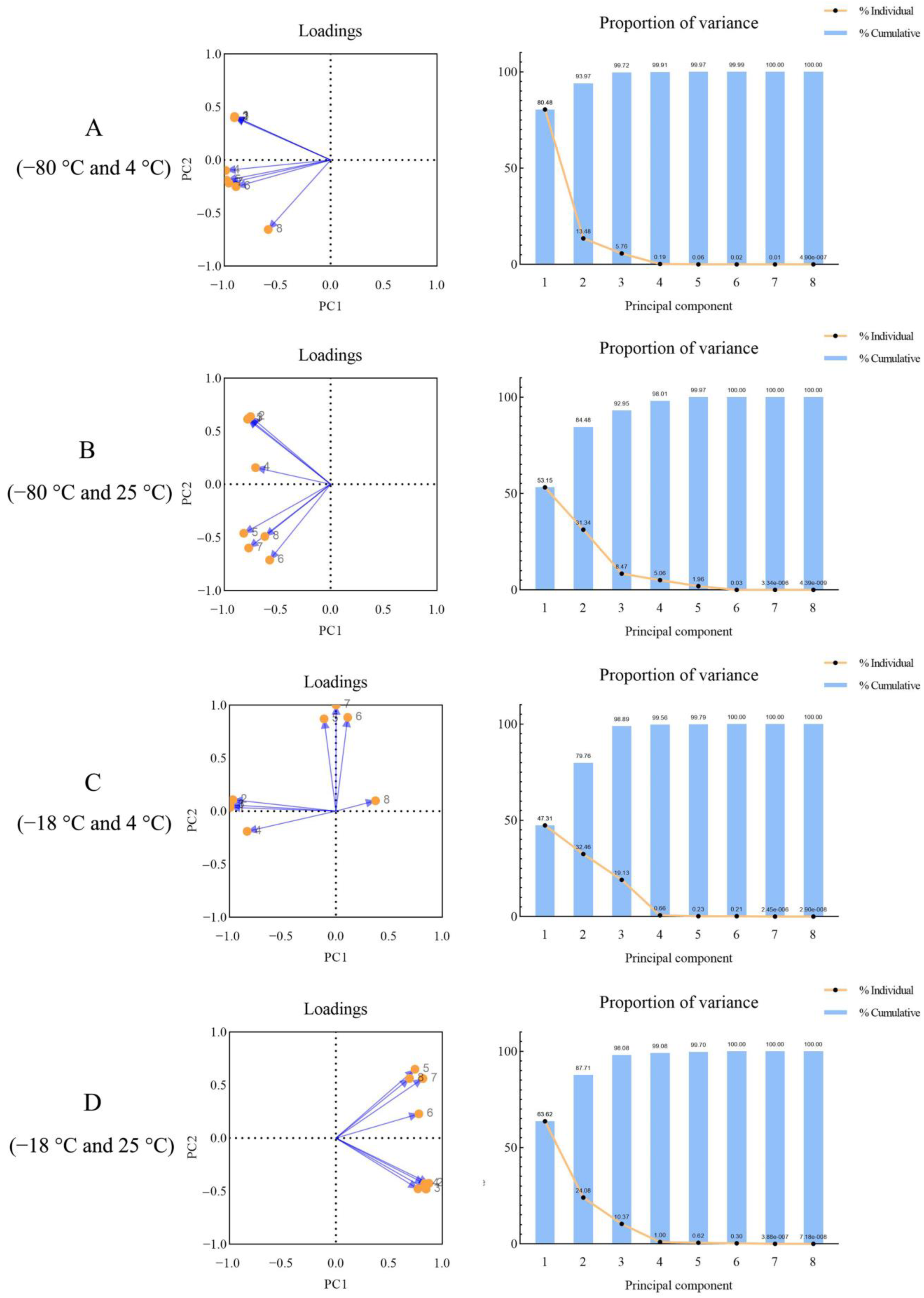
3.4. Principal Component Analysis of Celery Pigment Degradation with Different Treatment Temperature during Thawing
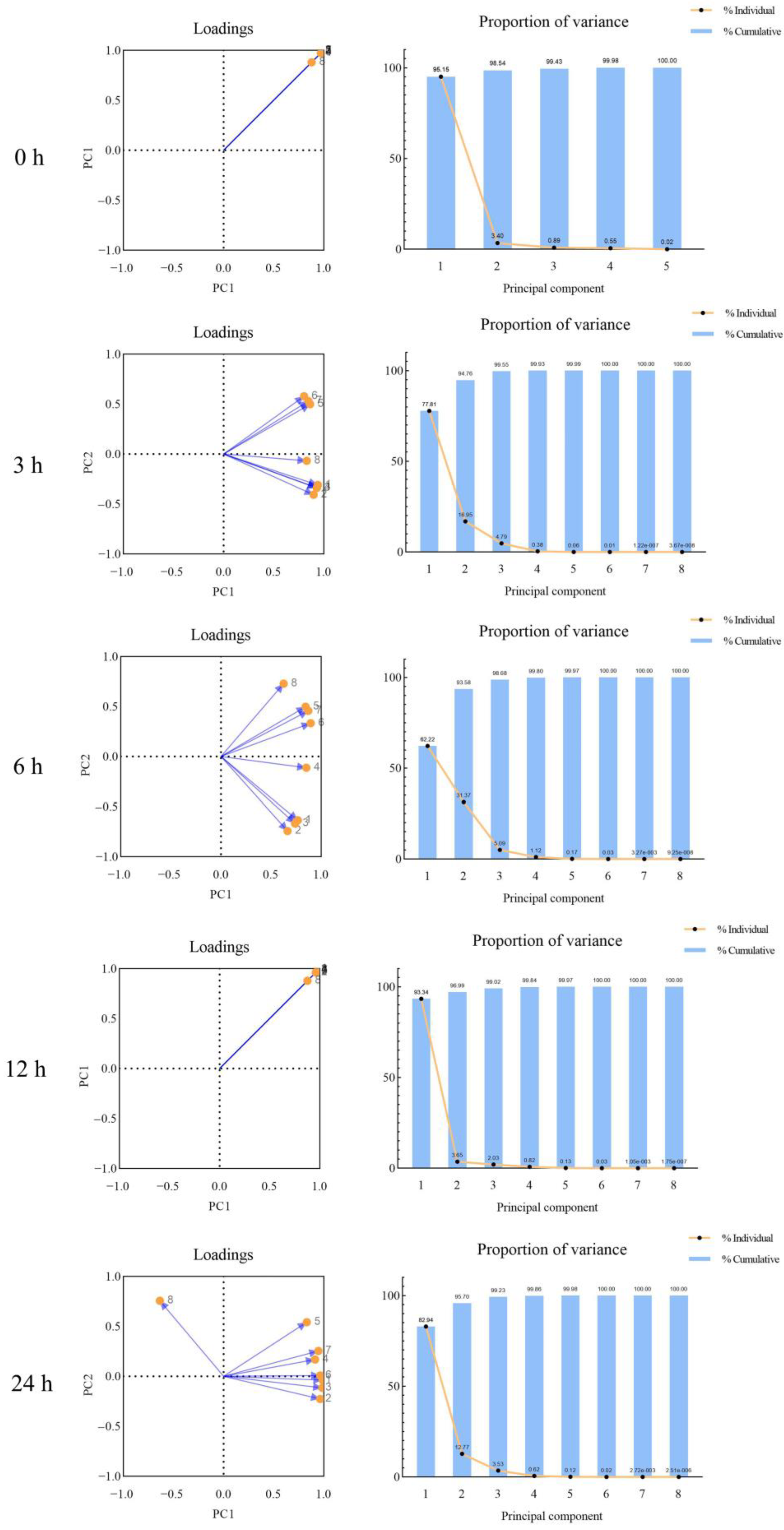
3.5. Principal Component Analysis of Celery Pigment Degradation with Different Thawing Time during Thawing
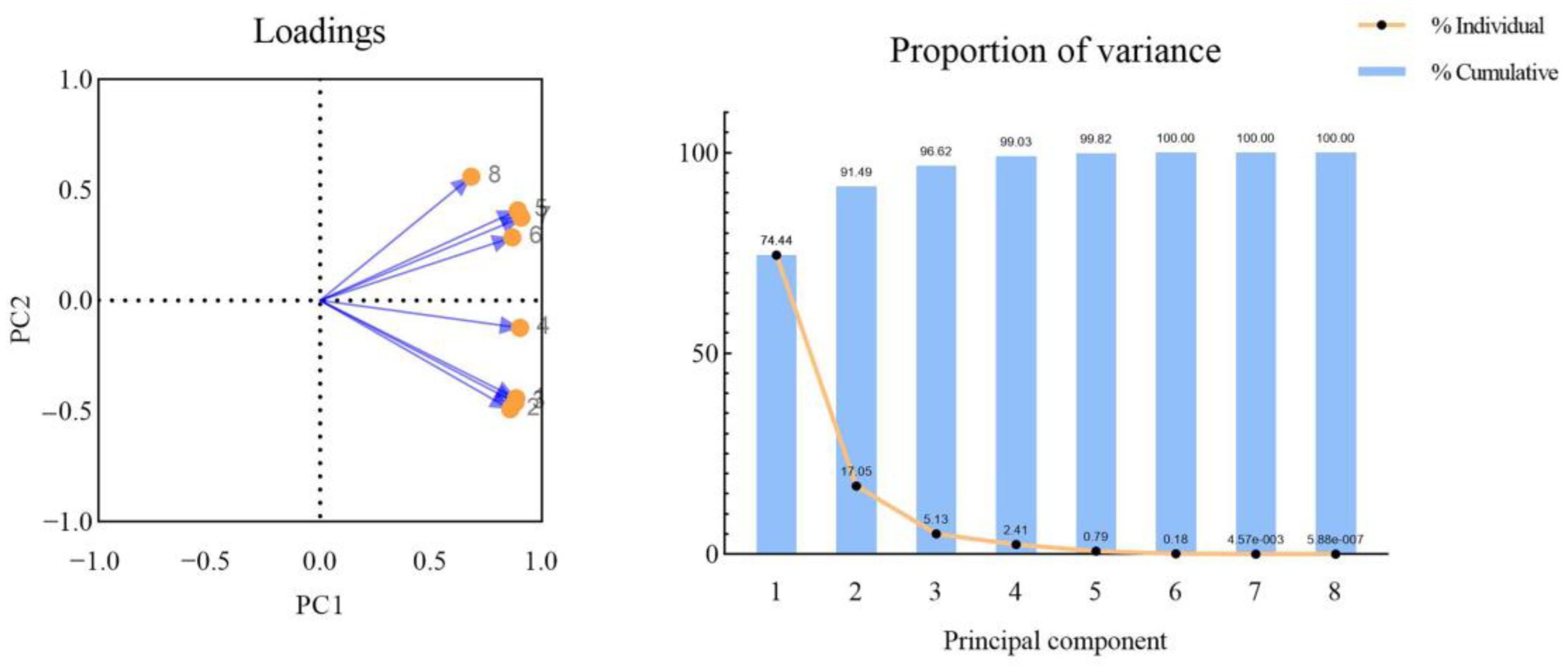
3.6. Principal Component Analysis of Celery Pigment Degradation of Different Leaves Parts during Thawing
4. Discussion
4.1. Relationship between Actual Photosynthetic Pigment Content and Light Absorption Content
4.2. Relationship between Temperature and Photosynthetic Pigment Degradation
4.3. Relationship between Thawing Time and Photosynthetic Pigment Degradation
4.4. Relationship between Plant Parts and Photosynthetic Pigment Degradation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maruyama, T.; Abbaskhan, A.; Choudhary, M.I.; Tsuda, Y.; Goda, Y.; Farille, M.; Reduron, J.P. Botanical origin of Indian celery seed (fruit). J. Nat. Med. 2009, 63, 248–253. [Google Scholar] [CrossRef]
- Li, J.W.; Feng, K.; Xu, Z.S.; Xiong, A.S. Transcriptome profiling of β-carotene biosynthesis genes and β-carotene accumulation in leaf blades and petioles of celery cv. Jinnanshiqin. Acta Biochim. Biophys. Sin. 2018, 51, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Feng, K.; Hou, X.L.; Jiang, Q.; Xu, Z.S.; Wang, G.L.; Liu, J.X.; Wang, F.; Xiong, A.S. The genome sequence of celery (Apium graveolens L.), an important leaf vegetable crop rich in apigenin in the Apiaceae family. Hortic. Res. 2020, 7, 223–253. [Google Scholar] [CrossRef]
- Li, M.Y.; Hou, X.L.; Wang, F.; Tan, G.F.; Xu, Z.S.; Xiong, A.S. Advances in the research of celery, an important Apiaceae vegetable crop. Crit. Rev. Biotechnol. 2018, 38, 172–183. [Google Scholar] [CrossRef]
- Wang, X.J.; Luo, Q.; Li, T.; Meng, P.H.; Pu, Y.T.; Liu, J.X.; Zhang, J.; Liu, H.; Tan, G.F.; Xiong, A.S. Origin, evolution, breeding, and omics of Apiaceae: A family of vegetables and medicinal plants. Hortic. Res. 2022, 9, uhac076. [Google Scholar] [CrossRef]
- Li, M.Y.; Li, J.; Xie, F.J.; Zhou, J.; Sun, Y.; Luo, Y.; Zhang, Y.; Chen, Q.; Wang, Y.; Lin, Y.X.; et al. Combined evaluation of agronomic and quality traits to explore heat germplasm in celery (Apium graveolens L.). Sci. Hortic. 2023, 317, 112039. [Google Scholar] [CrossRef]
- Ma, P.H.; Pong, L.; Yu, N.; Li, A.; Liu, P.; Wang, Q.; Sheng, J.P. Image-based nutrient estimation for Chinese dishes using deep learning. Food Res. 2021, 147, 110437. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Wang, F.; Jiang, Q.; Ma, J.; Xiong, A.S. Identification of SSRs and differentially expressed genes in two cultivars of celery (Apium graveolens L.) by deep transcriptome sequencing. Hortic. Res. 2014, 1, 10. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Cao, S.F.; Xie, K.Q.; Chi, Z.Y.; Wang, J.; Wang, H.f.; Wei, Y.Y.; Shao, X.F.; Zhang, C.D.; Xu, F.; et al. Melatonin delays yellowing of broccoli during storage by regulating chlorophyll catabolism and maintaining chloroplast ultrastructure. Postharvest Biol. Technol. 2021, 172, 111378. [Google Scholar] [CrossRef]
- Ilic, Z.S.; Sunic, L.J.; Milenkovic, L. Extended harvest time improves the shelf life of celeriac (Apium graveolens var. rapaceum) through postharvest treatment and storage conditions. Acta Hortic. 2016, 1142, 269–275. [Google Scholar]
- Jia, M.; Zhu, S.Q.; Wang, Y.H.; Liu, J.X.; Tan, S.S.; Liu, H.; Shu, S.; Tao, J.P.; Xiong, A.S. Morphological characteristics, anatomical structure, and dynamic change of ascorbic acid under different storage conditions of celery. Protoplasma 2023, 260, 21–33. [Google Scholar] [CrossRef]
- Li, M.Y.; Wang, Y.; Wei, X.H.; Wang, Z.; Wang, C.; Du, X.M.; Lin, Y.X.; Zhang, Y.T.; Wang, Y.; He, W.; et al. Effects of pretreatment and freezing storage on the bioactive components and antioxidant activity of two kinds of celery after postharvest. Food Chem. X 2023, 18, 100655. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.; Li, T.; Wang, H.; Liu, Y.H.; Feng, K.; Duan, A.Q.; Liu, H.; Shu, S.; Xiong, A.S. CRISPR/Cas9-mediated precise targeted mutagenesis of phytoene desaturase in celery. Hortic. Res. 2022, 9, uhac162. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Liu, H.; Tao, J.P.; Zhang, J.Q.; Zhao, T.M.; Hou, X.L.; Xiong, A.S.; You, X. Low light intensity elongates period and defers peak time of photosynthesis: A computational approach to circadian-clock-controlled photosynthesis in tomato. Hortic. Res. 2023, 10, uhad077. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Ma, Y.H.; Huang, Y.; Li, Y.; Wang, G.L.; Jiang, Q.; Wang, F.; Xiong, A.S. Comparative proteomic analysis provides novel insights into chlorophyll biosynthesis in celery under temperature stress. Physiol. Plant 2017, 161, 468–485. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Jia, L.L.; Xing, G.M.; Tao, J.P.; Sun, S.; Tan, G.F.; Li, S.; Liu, J.X.; Duan, A.Q.; Wang, H.; et al. The accumulation of lutein and β-carotene and transcript profiling of genes related to carotenoids biosynthesis in yellow celery. Mol Biotechnol. 2021, 63, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.Y.; Zhang, Y.Z.; Tang, W.J.; Chen, J.X.; Ge, Y.H.; Li, J.R. Concentration-dependent impacts of exogenous methyl jasmonate (MeJA) on chlorophyll degradation of apple fruit during ripening. Postharvest Biol. Technol. 2023, 203, 112398. [Google Scholar] [CrossRef]
- Sharafi, E.; Dehestani, A.; Farmani, J.; Parizi, A.P. Bioinformatics evaluation of plant chlorophyllase, the key enzyme in chlorophyll degradation. Appl. Food Biotechnol. 2017, 4, 167–178. [Google Scholar]
- Yin, L.; Liu, J.X.; Tao, J.P.; Xing, G.M.; Tan, G.F.; Li, S.; Duan, A.Q.; Ding, X.; Xu, Z.S.; Xiong, A.S. The gene encoding lycopene epsilon cyclase of celery enhanced lutein and β-carotene contents and confers increased salt tolerance in Arabidopsis. Plant Physiol. Biochem. 2020, 157, 339–347. [Google Scholar] [CrossRef]
- Raffo, A.; Sinesio, F.; Moneta, E.; Nardo, N.; Peparaio, M.; Paoletti, F. Internal quality of fresh and cold stored celery petioles described by sensory profile, chemical and instrumental measurements. Eur. Food Res. Technol. 2006, 222, 590–599. [Google Scholar] [CrossRef]
- Ding, X.; Liu, J.X.; Li, T.; Duan, A.Q.; Yin, L.; Wang, H.; Jia, L.L.; Liu, Y.H.; Liu, H.; Tao, J.P.; et al. AgZDS, a gene encoding ζ-carotene desaturase, increases lutein and β-carotene contents in transgenic Arabidopsis and celery. Plant Sci. 2021, 312, 111043. [Google Scholar] [CrossRef]
- Kręcisz, M.; Kolniak-Ostek, J.; Łyczko, J.; Stępień, B. Evaluation of bioactive compounds, volatile compounds, drying process kinetics and selected physical properties of vacuum impregnation celery dried by different methods. Food Chem. 2023, 413, 135490. [Google Scholar] [CrossRef]
- Lee, S.M.; Lee, J.Y.; Cho, Y.J.; Kim, M.S.; Kim, Y.S. Determination of volatiles and carotenoid degradation compounds in red pepper fermented by Lactobacillus parabuchneri. J. Food Sci. 2018, 83, 2083–2091. [Google Scholar] [CrossRef]
- Zhu, Z.P.; Yu, J.X.; Qiao, X.H.; Yu, Z.F.; Xiong, A.S.; Sun, M. Hydrogen sulfide delays yellowing and softening, inhibits nutrient loss in postharvest celery. Sci. Hortic. 2023, 315, 111991. [Google Scholar] [CrossRef]
- Kim, M.; Lee, J.; Choe, E. Pigment changes in fried dough containing spinach powder during storage in the dark. J. Food Sci. 2003, 68, 1923–1927. [Google Scholar] [CrossRef]
- Chen, N.X.; Li, J.B.; Meng, C.L.; Fan, S.X. Research progress in preservation of postharvest leafy vegetables. Adv. Mater. Res. 2013, 2512, 749. [Google Scholar] [CrossRef]
- Sun, M.; Yang, T.; Qiao, X.H.; Zhao, P.; Zhu, Z.P.; Wang, G.L.; Xu, L.L.; Xiong, A.S. Nitric oxide regulates the lignification and carotenoid biosynthesis of postharvest carrot (Daucus carota L.). Postharvest Biol. Technol. 2024, 207, 112593. [Google Scholar] [CrossRef]
- Chen, Q.; Li, J.L.; Yang, H.; Qian, J.Q. A dynamic shelf-life prediction method considering actual uncertainty: Application to fresh fruits in long-term cold storage. J. Food Eng. 2023, 349, 111471. [Google Scholar] [CrossRef]
- Xu, Y.X.; Chen, X.N.; Xu, L.; Du, B. The research on modified atmosphere packaging preservation of fresh-cut iceberg lettuce. Adv. Gr. Commun. Packag. Technol. Mater. 2016, 369, 549–559. [Google Scholar]
- Guyer, L.; Salinger, K.; Krugel, U.; Hortensteiner, S. Catalytic and structural properties of pheophytinase, the phytol esterase involved in chlorophyll breakdown. J. Exp. Bot. 2018, 69, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.W.; Xin, F.Y.; Xu, H.J.L.; Chu, Y.Y.; Du, Y.L.; Tian, H.Q.; Zhu, B.Z. Chitosan inhibits postharvest berry abscission of ‘Kyoho’ table grapes by affecting the structure of abscission zone, cell wall degrading enzymes and SO2 permeation. Postharvest Biol. Technol. 2021, 176, 111507. [Google Scholar] [CrossRef]
- Saba, A.; Moneta, E.; Peparaio, M.; Sinesio, F.; Vassallo, M.; PaolettiSaba, F. Towards a multi-dimensional concept of vegetable freshness from the consumer’s perspective. Food Qual. Pref. 2018, 66, 1–12. [Google Scholar] [CrossRef]
- Ningrum, A.; Minh, N.N.; Schreiner, M. Carotenoids and norisoprenoids as carotenoid degradation products in pandan leaves (Pandanus amaryllifolius Roxb.). Int. J. Food Prop. 2015, 18, 1905–1914. [Google Scholar] [CrossRef]
- Behzadi, A.; Holmberg, S.; Duwig, C.; Haghighat, F.; Ooka, R.; Sadrizadeh, S. Smart design and control of thermal energy storage in low temperature heating and high-temperature cooling systems: A comprehensive review. Renew. Sustain. Energy Rev. 2022, 166, 116625. [Google Scholar] [CrossRef]
- Biehler, E.; Mayer, F.; Hoffmann, L.; Krause, E.; Bohn, T. Comparison of 3 spectrophotometric methods for carotenoid determination in frequently consumed fruits and vegetables. J. Food Sci. 2010, 75, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Dermesonluoglu, E.; Katsaros, G.; Tsevdou, M.; Giannakourou, M.; Taoukis, P. Kinetic study of quality indices and shelf life modelling of frozen spinach under dynamic conditions of the cold chain. J. Food Eng. 2015, 148, 13–23. [Google Scholar] [CrossRef]
- Luo, S.; Luo, T.; Peng, P.; Li, Y.P.; Li, X.G. Disturbance of chlorophyll biosynthesis at Mg branch affects the chloroplast ROS homeostasis and Ca2+ signaling in Pisum sativum. Plant Cell 2016, 127, 729–737. [Google Scholar] [CrossRef]
- Shimoda, Y.; Ito, S.; Tanaka, A. Arabidopsis STAY-GREEN, Mendel’s green cotyledon gene, encodes magnesium—Dechelatase. Plant Cell 2016, 28, 2147–2160. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, N.D.; Dhuique-Mayer, C.; Cisse, M.; Dornier, M. Identification and thermal degradation kinetics of chlorophyll pigments and ascorbic acid from ditax nectar (Detarium senegalense J.F. Gmel). J. Agric. Food Chem. 2011, 59, 12018–12027. [Google Scholar] [CrossRef]
- Jang, W.; Lee, C.; Suh, H.J.; Lee, J. β-Carotene and β-apo-8′-carotenal contents in processed foods in Korea. Food Sci. Biotechnol. 2023, 32, 1501–1513. [Google Scholar] [CrossRef]
- Mehmood, A.; Zeb, A. Effects of different cooking techniques on the carotenoids composition, phenolic contents, and antioxidant activity of spinach leaves. J. Food Meas. Charact. 2023, 17, 4760–4774. [Google Scholar] [CrossRef]
- Su, S.Q.; Zhou, Y.M.; Qin, J.G.; Yao, W.Z.; Ma, Z.H. Optimization of the Method for Chlorophyll Extraction in Aquatic Plants. J. Freshw. Ecol. 2010, 25, 531–538. [Google Scholar] [CrossRef]
- Bashir, T.; Haq, S.A.U.; Masoom, S.; Ibdah, M.; Husaini, A.M. Quality trait improvement in horticultural crops: OMICS and modern biotechnological approaches. Mol. Biol. Rep. 2023, 50, 87298742. [Google Scholar] [CrossRef] [PubMed]
- Boroda, A.V.; Aizdaicher, N.A.; Odintsova, N.A. The influence of ultra-low temperatures on marine microalgal cells. J. Appl. Phycol. 2014, 26, 387–397. [Google Scholar] [CrossRef]
- Yao, J.H.; Chen, W.J.; Fan, K. Novel efficient physical technologies for enhancing freeze drying of fruits and vegetables: A review. Foods 2023, 12, 4321. [Google Scholar] [CrossRef] [PubMed]
- Funamoto, Y.; Yamauchi, N.; Masayoshi, S. Involvement of peroxidase in chlorophyll degradation in stored broccoli (Brassica oleracea L.) and inhibition of the activity by heat treatment. Postharvest Biol. Technol. 2003, 28, 39–46. [Google Scholar] [CrossRef]
- Pathare, P.B.; Rahman, M.S. Nondestructive Quality Assessment Techniques for Fresh Fruits and Vegetables; Springer: Singapore, 2022; pp. 163–188. [Google Scholar]
- Hasan, M.U.; Singh, Z.; Shah, H.M.S.; Kaur, J.; Woodward, A.; Afrifa-Yamoah, E.; Malik, A.U. Oxalic acid: A blooming organic acid for postharvest quality preservation of fresh fruit and vegetables. Postharvest Biol. Technol. 2023, 206, 112574. [Google Scholar] [CrossRef]
- Cai, J.H.; Luo, F.; Zhao, Y.B.; Zhou, Q.; Wei, B.D.; Zhou, J.; Ji, S.J. 24-Epibrassinolide treatment regulates broccoli yellowing during shelf life. Postharvest Biol. Technol. 2019, 154, 87–95. [Google Scholar] [CrossRef]
- Brown, W.; Ryser, E.; Gorman, L.; Steinmaus, S.; Vorst, K. Transit temperatures experienced by fresh-cut leafy greens during cross-country shipment. Food Control 2016, 61, 146–155. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, G.H.; Wen, W.W.; Ma, X.Q.; Xu, B.; Huang, B.R. Functional characterization and hormonal regulation of the pheophytinase gene LpPPH controlling leaf senescence in perennial ryegrass. J. Exp. Bot. 2016, 67, 935–945. [Google Scholar] [CrossRef]
- Büchert, A.M.; Civello, P.M.; Gustavo, A. Chlorophyllase versus pheophytinase as candidates for chlorophyll dephytilation during senescence of broccoli. J. Plant Physiol. 2010, 168, 337–343. [Google Scholar] [CrossRef]
- Cuadra-Crespo, P.; Amor, F.M. Effects of postharvest treatments on fruit quality of sweet pepper at low temperature. J. Sci. Food Agric. 2010, 90, 2716–2722. [Google Scholar] [CrossRef]
- Dhemre, K.J.; Shete, B.M.; Kotecha, M.P. Performance of packaging on shelf life and quality of fenugreek at different storage conditions in kharif season. Adv. Res. 2017, 10, 35266. [Google Scholar] [CrossRef]
- Marie, L.G.; Luca, V.; Valentina, G.; Lazazzara, V.; Zanella, A.; Robatscher, P.; Scampicchio, M.; Oberhuber, M. Chlorophyll breakdown during fruit ripening: Qualitative analysis of phyllobilins in the peel of apples (Malus domestica Borkh.) cv. ‘Gala’ during different shelf life stages. Food Res. Int. 2022, 162, 112061. [Google Scholar]
- Wang, N.; Kong, X.M.; Luo, M.L.; Sun, Y.Y.; Liu, Z.Y.; Feng, H.; Ji, S.J. SGR mutation in pak choi prolongs its shelf life by retarding chlorophyll degradation and maintaining membrane function. Postharvest Biol. Technol. 2022, 191, 112011. [Google Scholar] [CrossRef]
- Shi, J.Y.; Gao, L.P.; Zuo, J.H.; Wang, Q.; Fan, L.L. Exogenous sodium nitroprusside treatment of broccoli florets extends shelf life, enhances antioxidant enzyme activity, and inhibits chlorophyll-degradation. Postharvest Biol. Technol. 2016, 116, 98–104. [Google Scholar] [CrossRef]
- Lee, D.S.; Robertson, G. Interactive influence of decision criteria, packaging film, storage temperature and humidity on shelf life of packaged dried vegetables. Food Packag. Shelf Life 2021, 28, 100674. [Google Scholar] [CrossRef]
- Lara, A.; Dalene, B.D.; Magdalena, M.; Marieta, V.D.R.; Elizabeth, J. Impact of steam treatment on shelf-life stability of a xanthone-rich green herbal tea (Cyclopia maculata Andrews Kies)—Identifying quality changes during storage. J. Sci Food Agric. 2019, 99, 1334–1341. [Google Scholar]
- Nduwumuremyi, A.; Melis, R.; Shanahan, P.; Asiimwe, T. Introgression of antioxidant activity into cassava (Manihot esculenta C.): An effective technique for extending fresh storage roots shelf life. Plant Breed. 2016, 135, 1–8. [Google Scholar] [CrossRef]
- Lara, I.; García, P.; Vendrell, M. Modifications in cell wall composition after cold storage of calcium-treated strawberry (Fragaria × ananassa Duch.) fruit. Postharvest Biol. Technol. 2004, 34, 331–339. [Google Scholar] [CrossRef]
- Li, T.; Bai, D.Q.; Tian, L.; Li, P.; Liu, Y.H.; Jiang, Y. Effects of stimulators on lutein and chlorophyll biosyntheses in the green alga chlorella pyrenoidosa under heterotrophic conditions. Adv. Appl. Biotechnol. 2015, 1, 28–30. [Google Scholar]
- Wu, X.F.; Zhang, M.; Adhikari, B.; Sun, J.C. Recent advances for rapid freezing and thawing methods of foods. Crit. Rev. Food Sci. Nutr. 2017, 57, 3620–3631. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Freezing Temperature | Thawing Temperature | Measure Time and Group | ||||
|---|---|---|---|---|---|---|
| 0 h | 3 h | 6 h | 12 h | 24 h | ||
| −80 °C | 4 °C | A1 | A2 | A3 | A4 | A5 |
| 25 °C | B1 | B2 | B3 | B4 | B5 | |
| −18 °C | 4 °C | C1 | C2 | C3 | C4 | C5 |
| 25 °C | D1 | D2 | D3 | D4 | D5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Wang, L.-X.; Li, M.-Y.; Tan, G.-F.; Liu, Y.-H.; Liu, P.-Z.; Li, Y.-P.; Liu, H.; Zhuang, J.; Tao, J.-P.; et al. Effect of Temperature on Photosynthetic Pigment Degradation during Freeze–Thaw Process of Postharvest of Celery Leaves. Horticulturae 2024, 10, 267. https://doi.org/10.3390/horticulturae10030267
Chen C, Wang L-X, Li M-Y, Tan G-F, Liu Y-H, Liu P-Z, Li Y-P, Liu H, Zhuang J, Tao J-P, et al. Effect of Temperature on Photosynthetic Pigment Degradation during Freeze–Thaw Process of Postharvest of Celery Leaves. Horticulturae. 2024; 10(3):267. https://doi.org/10.3390/horticulturae10030267
Chicago/Turabian StyleChen, Chen, Li-Xiang Wang, Meng-Yao Li, Guo-Fei Tan, Yan-Hua Liu, Pei-Zhuo Liu, Ya-Peng Li, Hui Liu, Jing Zhuang, Jian-Ping Tao, and et al. 2024. "Effect of Temperature on Photosynthetic Pigment Degradation during Freeze–Thaw Process of Postharvest of Celery Leaves" Horticulturae 10, no. 3: 267. https://doi.org/10.3390/horticulturae10030267
APA StyleChen, C., Wang, L.-X., Li, M.-Y., Tan, G.-F., Liu, Y.-H., Liu, P.-Z., Li, Y.-P., Liu, H., Zhuang, J., Tao, J.-P., & Xiong, A.-S. (2024). Effect of Temperature on Photosynthetic Pigment Degradation during Freeze–Thaw Process of Postharvest of Celery Leaves. Horticulturae, 10(3), 267. https://doi.org/10.3390/horticulturae10030267