
The Growth and Tuber Yield of Potatoes (Solanum tuberosum L.) under Varying LED Light Spectrums in Controlled Greenhouse Conditions
 ,
,  , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Plant Growth Conditions and Light-Emitting Diode (LED) Settings and Nutrient Growth
2.3. Leaf Gas Exchange Measurement
2.4. Analysis of Photosynthetic Pigments
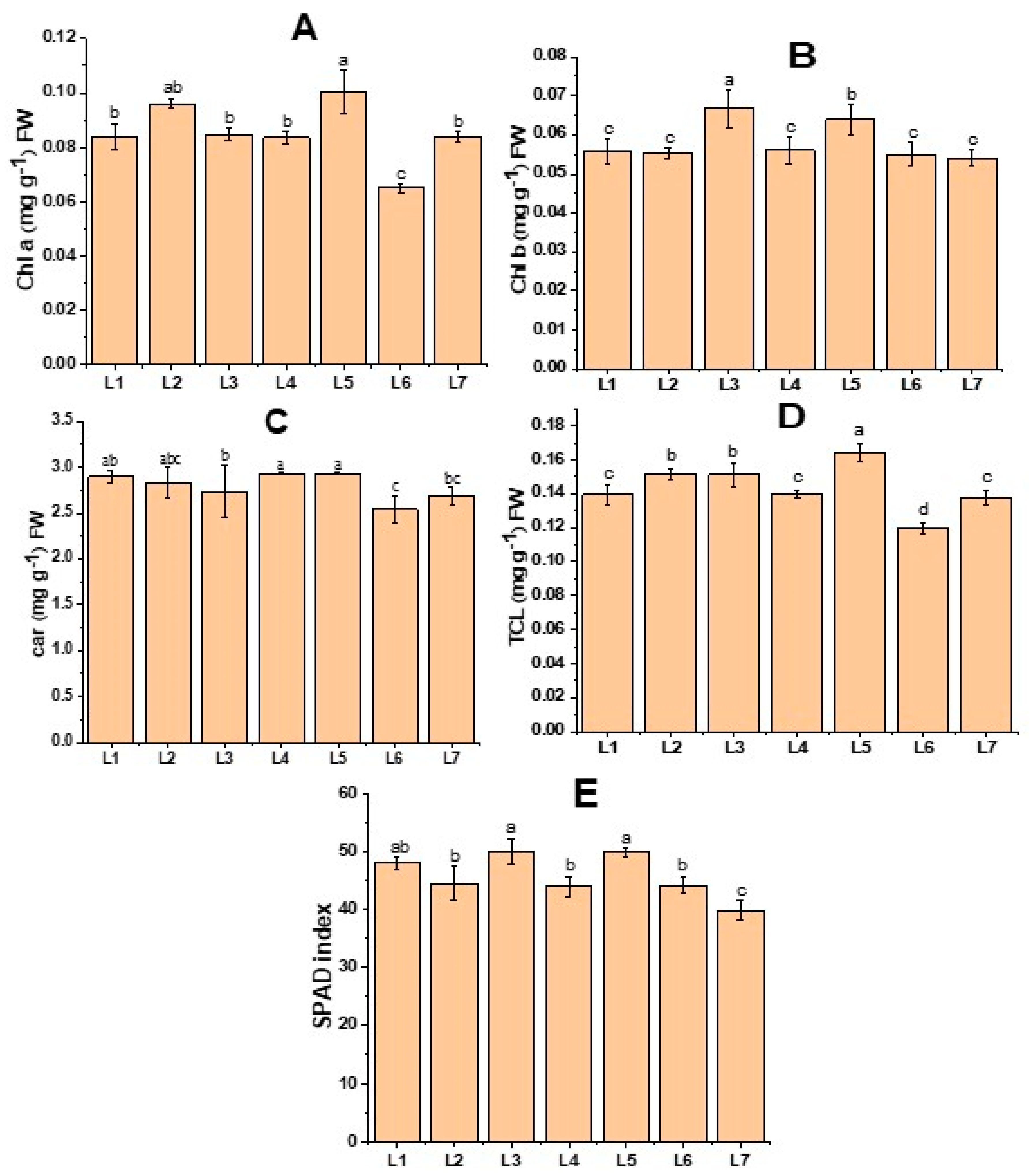
2.5. SPAD Index Analysis
2.6. GA3 Analysis Pretreatment Methods
2.7. Tuber Yield
2.8. Statistical Analysis
3. Results and Discussion
3.1. The Morphological Characteristics of Potato Plants Grown under LED Light
3.2. Effects of Different LED Light Treatments on Chlorophyll Content
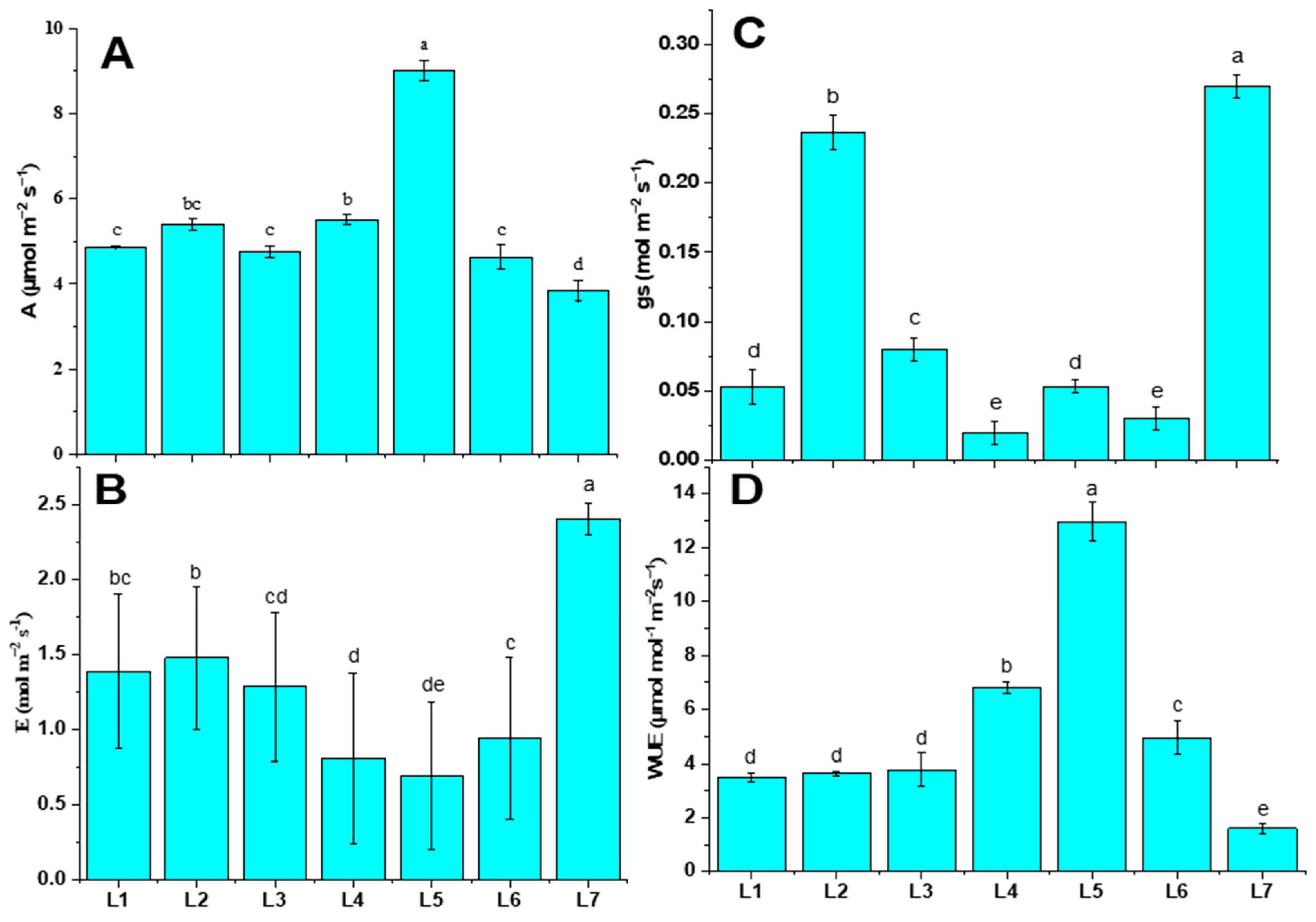
3.3. Photosynthetic Gas Exchange under Different LED Treatment
3.4. Potato Tuber Yield and Grading
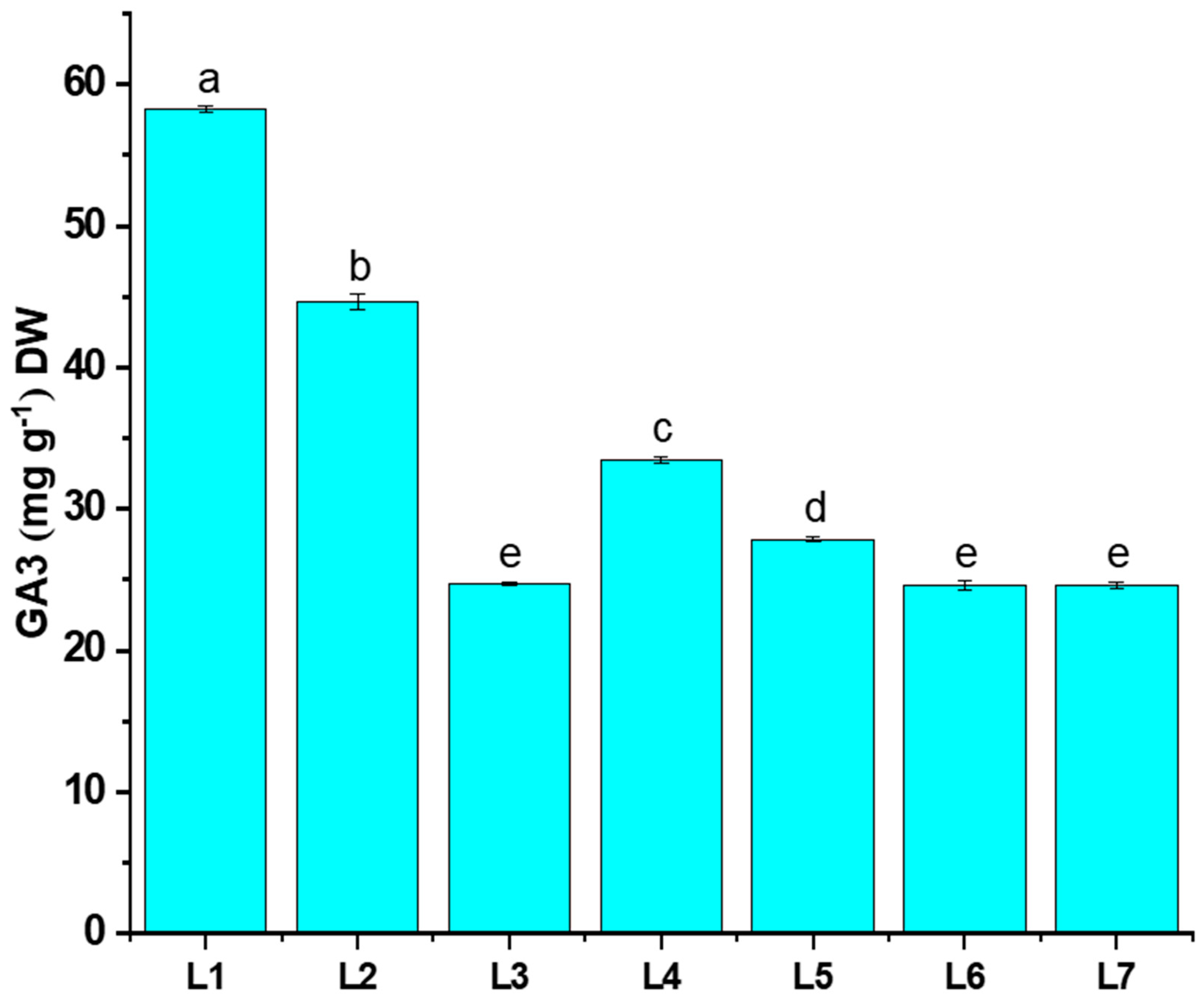
3.5. Gibberellic Acid Content Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Da Rocha, P.S.G.; de Oliveira, R.P.; Scivittaro, W.B. New light sources for in-vitro potato micropropagation. Biosci. J. 2015, 31, 1312–1318. [Google Scholar] [CrossRef]
- Olle, M.; Viršile, A. The effects of light-emitting diode lighting on greenhouse plant growth and quality. Agric. Food Sci. 2013, 22, 223–234. [Google Scholar] [CrossRef]
- Grishchenko, O.V.; Subbotin, E.P.; Gafitskaya, I.V.; Vereshchagina, Y.V.; Burkovskaya, E.V.; Khrolenko, Y.A.; Grigorchuk, V.P.; Nakonechnaya, O.V.; Bulgakov, V.P.; Kulchin, Y.N. Growth of micropropagated Solanum tuberosum L. plantlets under artificial solar spectrum and different mono-and polychromatic LED lights. Hortic. Plant J. 2022, 8, 205–214. [Google Scholar] [CrossRef]
- Thomas-Sharma, S.; Abdurahman, A.; Ali, S.; Andrade-Piedra, J.L.; Bao, S.; Charkowski, A.O.; Crook, D.; Kadian, M.; Kromann, P.; Struik, P.C.; et al. Seed degeneration in potato: The need for an integrated seed health strategy to mitigate the problem in developing countries. Plant Pathol. 2016, 65, 3–16. [Google Scholar] [CrossRef]
- Rahman, M.H.; Azad, M.O.K.; Islam, M.J.; Rana, M.S.; Li, K.; Lim, Y.S. Production of Potato (Solanum tuberosum L.) Seed Tuber under Artificial LED Light Irradiation in Plant Factory. Plants 2021, 10, 297. [Google Scholar] [CrossRef]
- Hussain, A.; Qarshi, I.A.; Nazir, H.; Ullah, I. Plant tissue culture: Current status and opportunities. Recent Adv. Plant Vitr. Cult. 2012, 6, 1–28. [Google Scholar]
- Rongfu, G.; Hongming, Z. Advances of researches on photoregulation in plants. J. Beijing For. Univ. 2002, 24, 235–243. [Google Scholar]
- Islam, M.J.; Hassan, M.K.; Sarker, S.R.; Rahman, A.B.; Fakir, M.S.A. Light and temperature effects on sprout yield and its proximate composition and vitamin C content in lignosus and mung beans. J. Bangladesh Agric. Univ. 2017, 15, 248–254. [Google Scholar] [CrossRef]
- Wenke, L. Light environmental management for artificial protected horticulture. Agrotechnology 2012, 1, 101. [Google Scholar]
- Lin, K.-H.; Huang, M.-Y.; Huang, W.-D.; Hsu, M.-H.; Yang, Z.-W.; Yang, C.-M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Shimizu, H.; Saito, Y.; Nakashima, H.; Miyasaka, J.; Ohdoi, K. Light environment optimization for lettuce growth in plant factory. IFAC Proc. Vol. 2011, 44, 605–609. [Google Scholar] [CrossRef]
- Yorio, N.C.; Goins, G.D.; Kagie, H.R.; Wheeler, R.M.; Sager, J.C. Improving spinach, radish, and lettuce growth under red light-emitting diodes (LEDs) with blue light supplementation. HortScience 2001, 36, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef]
- Hanyu, H.; Shoji, K. Acceleration of growth in spinach by short-term exposure to red and blue light at the beginning and at the end of the daily dark period. In Proceedings of the IV International ISHS Symposium on Artificial Lighting 580, Québec City, QC, Canada, 7–9 November 2000; pp. 145–150. [Google Scholar]
- Seabrook, J.E.A. Light effects on the growth and morphogenesis of potato (Solanum tuberosum) in vitro: A review. Am. J. Potato Res. 2005, 82, 353–367. [Google Scholar] [CrossRef]
- Paradiso, R.; Arena, C.; Rouphael, Y.; d’Aquino, L.; Makris, K.; Vitaglione, P.; De Pascale, S. Growth, photosynthetic activity and tuber quality of two potato cultivars in controlled environment as affected by light source. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2019, 153, 725–735. [Google Scholar] [CrossRef]
- Rahman, M.H.; Islam, M.J.; Azad, M.O.K.; Rana, M.S.; Ryu, B.R.; Lim, Y.-S. LED Light pre-treatment improves pre-basic seed potato (Solanum tuberosum L. cv. Golden King) production in the aeroponic system. Agronomy 2021, 11, 1627. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. ISBN 0076-6879. [Google Scholar]
- Han, T.; Vaganov, V.; Cao, S.; Li, Q.; Ling, L.; Cheng, X.; Peng, L.; Zhang, C.; Yakovlev, A.N.; Zhong, Y. Improving “color rendering” of LED lighting for the growth of lettuce. Sci. Rep. 2017, 7, 45944. [Google Scholar] [CrossRef]
- Schellenberger Costa, B.; Jungandreas, A.; Jakob, T.; Weisheit, W.; Mittag, M.; Wilhelm, C. Blue light is essential for high light acclimation and photoprotection in the diatom Phaeodactylum tricornutum. J. Exp. Bot. 2013, 64, 483–493. [Google Scholar] [CrossRef]
- Amoozgar, A.; Mohammadi, A.; Sabzalian, M.R. Impact of light-emitting diode irradiation on photosynthesis, phytochemical composition and mineral element content of lettuce cv. Grizzly. Photosynthetica 2017, 55, 85–95. [Google Scholar] [CrossRef]
- Ma, X.; Wang, Y.; Liu, M.; Xu, J.; Xu, Z. Effects of green and red lights on the growth and morphogenesis of potato (Solanum tuberosum L.) plantlets in vitro. Sci. Hortic. 2015, 190, 104–109. [Google Scholar] [CrossRef]
- Jiang, C.-D.; Jiang, G.-M.; Wang, X.; Li, L.-H.; Biswas, D.K.; Li, Y.-G. Increased photosynthetic activities and thermostability of photosystem II with leaf development of elm seedlings (Ulmus pumila) probed by the fast fluorescence rise OJIP. Environ. Exp. Bot. 2006, 58, 261–268. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose–responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, K.; Gong, X.; Wang, H.; Gao, Y.; Wang, X.; Zeng, Z.; Hu, Y. Effects of different LEDs light spectrum on the growth, leaf anatomy, and chloroplast ultrastructure of potato plantlets in vitro and minituber production after transplanting in the greenhouse. J. Integr. Agric. 2020, 19, 108–119. [Google Scholar] [CrossRef]
- HU, Y. Effects of red and blue LEDs on in vitro growth and microtuberization of potato single-node cuttings. Front. Agric. Sci. Eng. 2018, 5, 197–205. [Google Scholar]
- Islam, M.J.; Ryu, B.R.; Rahman, M.H.; Rana, M.S.; Cheong, E.J.; Wang, M.-H.; Lim, J.-D.; Hossain, M.A.; Lim, Y.-S. Cannabinoid accumulation in hemp depends on ROS generation and interlinked with morpho-physiological acclimation and plasticity under indoor LED environment. Front. Plant Sci. 2022, 13, 984410. [Google Scholar] [CrossRef] [PubMed]
- Stober, K.; Lee, K.; Yamada, M.; Pattison, M. Energy Savings Potential of SSL in Horticultural Applications; EERE Publication and Product Library: Washington, DC, USA, 2017. [Google Scholar]
- Kusuma, P.; Pattison, P.M.; Bugbee, B. From physics to fixtures to food: Current and potential LED efficacy. Hortic. Res. 2020, 7, 56. [Google Scholar] [CrossRef]
- Kalaitzoglou, P.; Van Ieperen, W.; Harbinson, J.; van der Meer, M.; Martinakos, S.; Weerheim, K.; Nicole, C.; Marcelis, L.F.M. Effects of continuous or end-of-day far-red light on tomato plant growth, morphology, light absorption, and fruit production. Front. Plant Sci. 2019, 10, 322. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; van Iersel, M.W. Photosynthetic Physiology of Blue, Green, and Red Light: Light Intensity Effects and Underlying Mechanisms. Front. Plant Sci. 2021, 12, 328. [Google Scholar] [CrossRef]
- Nishio, J.N. Why are higher plants green? Evolution of the higher plant photosynthetic pigment complement. Plant. Cell Environ. 2000, 23, 539–548. [Google Scholar] [CrossRef]
- Aksenova, N.P.; Konstantinova, T.N.; Chailakhyan, M.K. Morphogenetic effect of blue and red light in the illumination of underground and above-ground potato organs in in vitro cultivation. Dokl. Akad. Nauk SSSR 1989, 305, 508–512. [Google Scholar]
- Matsuo, S.; Nanya, K.; Imanishi, S.; Honda, I.; Goto, E. Effects of blue and red lights on gibberellin metabolism in tomato seedlings. Hortic. J. 2019, 88, 76–82. [Google Scholar] [CrossRef]
- Eviatar-Ribak, T.; Shalit-Kaneh, A.; Chappell-Maor, L.; Amsellem, Z.; Eshed, Y.; Lifschitz, E. A cytokinin-activating enzyme promotes tuber formation in tomato. Curr. Biol. 2013, 23, 1057–1064. [Google Scholar] [CrossRef]
- Poudel, P.R.; Kataoka, I.; Mochioka, R. Effect of red-and blue-light-emitting diodes on growth and morphogenesis of grapes. Plant Cell Tissue Organ Cult. 2008, 92, 147–153. [Google Scholar] [CrossRef]
- Nhut, D.T.; Takamura, T.; Watanabe, H.; Tanaka, M. Efficiency of a novel culture system by using light-emitting diode (LED) on in vitro and subsequent growth of micropropagated banana plantlets. In Proceedings of the I International Symposium on Acclimatization and Establishment of Micropropagated Plants 616, Macedonia, Greece, 19–22 September 2001; pp. 121–127. [Google Scholar]
- Nhut, D.T.; Takamura, T.; Watanabe, H.; Okamoto, K.; Tanaka, M. Responses of strawberry plantlets cultured in vitro under superbright red and blue light-emitting diodes (LEDs). Plant Cell Tissue Organ Cult. 2003, 73, 43–52. [Google Scholar] [CrossRef]
- Kara, A.N.; Kotov, A.A.; Bukhov, N.G. Specific distribution of gibberellins, cytokinins, indole-3-acetic acid, and abscisic acid in radish plants closely correlates with photomorphogenetic responses to blue or red light. J. Plant Physiol. 1997, 151, 51–59. [Google Scholar] [CrossRef]
- Asghari-Zakaria, R.; Maleki-Zanjani, B.; Sedghi, E. Effect of in vitro chitosan application on growth and minituber yield of Solanum tuberosum L. Plant Soil Environ. 2009, 55, 252–256. [Google Scholar] [CrossRef]
- Su, N.; Wu, Q.; Cui, J. Effects of monochromatic LED light quality on endogenous hormone content in leaves and hypocotyl of Cucumis sativus L. Chin. J. Appl. Environ. Biol 2014, 20, 45–49. [Google Scholar]
- YU, L.; SONG, C.; SUN, L.; LI, L.; XU, Z.; TANG, C. Effects of light-emitting diodes on tissue culture plantlets and seedlings of rice (Oryza sativa L.). J. Integr. Agric. 2020, 19, 1743–1754. [Google Scholar] [CrossRef]
- Fukuda, N.; Ajima, C.; Yukawa, T.; Olsen, J.E. Antagonistic action of blue and red light on shoot elongation in petunia depends on gibberellin, but the effects on flowering are not generally linked to gibberellin. Environ. Exp. Bot. 2016, 121, 102–111. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Spectrum | Spectrum Combination | Intensity | Code Name |
|---|---|---|---|
| Red + blue * | 70 + 30 | 300 ** | L1 |
| Red + blue + white | 70 + 20 + 10 | 300 | L2 |
| Red + blue + green | 70 + 20 + 10 | 300 | L3 |
| Red + blue + far-red | 70 + 20 + 10 | 300 | L4 |
| Red + blue + green + white | 60 + 20 + 10 + 10 | 300 | L5 |
| Red + blue + far-red + white | 60 + 20 + 10 + 10 | 300 | L6 |
| Natural light | 250–300 | L7 |
| Treatments | Stem Length | Stem Diameter | Leaf Number | Leaf Length | Leaf Width | Branch Number | Plant Fresh Weight | Plant Dry Weight |
|---|---|---|---|---|---|---|---|---|
| L1 | 27.67 ± 2.05 d | 5.28 ± 0.08 bc | 11.6 ± 1.22 b | 13 ± 0.88 bc | 11.3 ± 0.49 b | 2.3 ± 0.39 b | 21 ± 0.82 de | 1.06 ± 0.03 bc |
| L2 | 37.6 ± 2.08 a | 5.24 ± 0.24 bc | 10. 7 ± 1.24 c | 13.16 ± 0.84 b | 9.7 ± 0.47 d | 2.6 ± 0.41 a | 36.3 ± 1.22 a | 2.68 ± 0.07 a |
| L3 | 27.6 ± 2.06 d | 6.12 ± 0.1 a | 13 ± 0.81 a | 13 ± 0.8 bc | 12.3 ± 0.41 a | 2 ± 0.44 c | 22.6 ± 2.05 d | 1.12 ± 0.02 bc |
| L4 | 33.3 ± 0.94 bc | 5.33 ± 0.06 bc | 8.7 ± 1.63 e | 14 ± 0.82 ab | 11 ± 0.78 b | 2.3 ± 0.47 b | 25.33 ± 0.47 cd | 1.14 ± 0.02 bc |
| L5 | 35.66 ± 1.69 b | 5.6 ± 0.33 b | 9.34 ± 1.2 d | 14.6 ± 1.21 a | 10.3 ± 0.44 c | 1.6 ± 0.41 d | 27.35 ± 0.94 c | 1.17 ± 0.01 bc |
| L6 | 23.3 ± 1.22 e | 5.24 ± 0.21 bc | 9 ± 0.8 de | 13 ± 0.81 bc | 12.33 ± 0.47 a | 1.3 ± 0.4 e | 30.38 ± 1.24 b | 1.32 ± 0.02 b |
| L7 | 32.6 ± 1.24 c | 4.31 ± 0.14 c | 9.33 ± 0.4 d | 11 ± 0.79 c | 10 ± 0.81 c | 1.3 ± 0.47 e | 19.6 ± 1.69 e | 1.08 ± 0.02 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.H.; Islam, M.J.; Mumu, U.H.; Ryu, B.-R.; Lim, J.-D.; Azad, M.O.K.; Cheong, E.J.; Lim, Y.-S. The Growth and Tuber Yield of Potatoes (Solanum tuberosum L.) under Varying LED Light Spectrums in Controlled Greenhouse Conditions. Horticulturae 2024, 10, 254. https://doi.org/10.3390/horticulturae10030254
Rahman MH, Islam MJ, Mumu UH, Ryu B-R, Lim J-D, Azad MOK, Cheong EJ, Lim Y-S. The Growth and Tuber Yield of Potatoes (Solanum tuberosum L.) under Varying LED Light Spectrums in Controlled Greenhouse Conditions. Horticulturae. 2024; 10(3):254. https://doi.org/10.3390/horticulturae10030254
Chicago/Turabian StyleRahman, Md Hafizur, Md. Jahirul Islam, Umma Habiba Mumu, Byeong-Ryeol Ryu, Jung-Dae Lim, Md Obyedul Kalam Azad, Eun Ju Cheong, and Young-Seok Lim. 2024. "The Growth and Tuber Yield of Potatoes (Solanum tuberosum L.) under Varying LED Light Spectrums in Controlled Greenhouse Conditions" Horticulturae 10, no. 3: 254. https://doi.org/10.3390/horticulturae10030254
APA StyleRahman, M. H., Islam, M. J., Mumu, U. H., Ryu, B.-R., Lim, J.-D., Azad, M. O. K., Cheong, E. J., & Lim, Y.-S. (2024). The Growth and Tuber Yield of Potatoes (Solanum tuberosum L.) under Varying LED Light Spectrums in Controlled Greenhouse Conditions. Horticulturae, 10(3), 254. https://doi.org/10.3390/horticulturae10030254