Abstract
Citrus plants are prone to phosphorus (P) deficiency, especially in acidic soil, making them more dependent on root-associated endophytic fungi for growth and development. Beni-Madonna, a hybrid of Citrus nanko × C. amakusa, is a citrus known as “tangor” that is highly popular in China and other parts of the world due to its deep red color and jelly-like flesh. In this study, the inoculation response to two arbuscular mycorrhizal fungi (Diversispora versiformis, Dv; Funneliformis mosseae, Fm) and an endophytic fungus (Serendipita indica, Si) with regard to the growth, leaf gas exchange, light energy efficiency, P levels, acid phosphatase activity, and expression of the purple acid phosphatase (PAP) and phosphate transporter (PT) genes through a potted experiment using Beni-Madonna (tangor) citrus plants grafted on trifoliate orange (Poncirus trifoliata) was studied. Two years following inoculation, the root fungal colonization rates of inoculated plants were significantly increased, accompanied by an increase in plant height and stem diameter, with Si presenting the best effect. Fungal inoculations also significantly increased the leaf chlorophyll index, nitrogen-balance index, photosynthesis rate, stomatal conductance, transpiration rate, photosynthetic efficiency, and quantum yield of PSII, while it reduced photoinhibition and heat dissipation, with Si having the best effect on light energy efficiency. Si significantly increased leaf and root P levels, as well as root CsPAP1‒3, CsPT2, CsPT3, and CsPT7 expression and soil acid phosphatase activity; Fm in mycorrhizal fungi significantly increased leaf and root P levels, as well as root CsPT1 and CsPT7 expression and root/soil acid phosphatase activity. These observations suggested an advantage of Si over the other two AMF in terms of improved plant growth and P acquisition, while Fm displayed prominent effects on increased photosynthetic efficiency.
1. Introduction
Beni-Madonna (“Ehime Kashi No. 28”) (Citrus nanko × C. amakusa) is known as “tangor”, a citrus hybrid developed by crossing the “Nankou” and “Amakusa” cultivars [1]. Its crimson-colored fruits have slag, high levels of sugar, a sweet orange-like aroma, and ripen between late November and early December [2], with hesperidin, naringenin, and vanilloid as the main flavonoids [3]. In China, Beni-Madonna tangor, one of the most popular early-ripening citrus hybrid varieties, is mostly planted in infertile soils with a prevalent phosphorus (P) deficiency, which results in a significant decline in productivity and quality over time.
Phosphorus is known for its function in plant growth and development [4], as well as other plant metabolic activities such as energy conversion, biomolecule synthesis, photosynthesis, and respiration [5]. In acidic soils, Olsen-P is a primary soil fertility constraint due to its chemical precipitation and adsorption of inorganic P [6,7], reducing crop growth and yield [4]. Plants acquire P via roots and symbiotic fungi [8]. Purple acid phosphatase (PAP) is a metal phosphatase that catalyzes the hydrolysis of phospholipids to release phosphate (Pi), which is essential for plant P uptake and transport [9,10]. Phosphate transporter (PT) is a carrier of inorganic Pi that is directly absorbed from the soil and redistributed within the plants, playing an important role in plant P acquisition [11]. An analysis of the expression of PAP and PT genes is anticipated to provide important clues to understanding the underlying mechanism of P uptake by mycorrhizal plants.
Soil arbuscular mycorrhizal fungi (AMF) are able to create symbiotic associations with the roots of nearly 80% of terrestrial plants [12,13]. AMF have been isolated in large numbers from citrus plants [14]. Potted experiments have demonstrated that AMF inoculation improves citrus plant growth, promotes nutrient uptake, improves fruit quality, and increases stress tolerance [15,16,17,18]. Cozzolino et al. [19] found that AMF inoculation increased maize plant growth, yield, and P uptake, while it decreased soil Olsen-P levels. AMF colonization significantly promoted soil acid and alkaline phosphatase activities, as well as P uptake in mango roots, resulting in higher mango yield [20]. Ren et al. [21] discovered that inoculation with Glomus mosseae significantly increased P uptake in alfalfa, whereas inoculation with G. intraradices had little effect on P uptake in the host plant. AMF proliferation in vitro is not yet on a large scale, limiting its application in the field. Furthermore, it is unclear whether AMF colonization promotes P uptake in prevalent Beni-Madonna tangor plants.
Serendipita indica (Si) is an endophytic fungus isolated from the rhizosphere of desert plants in India that can complete its life cycle in artificial media without plant roots and exhibits mycorrhizal fungus-like characteristics in facilitating the acquisition of P by host plants [5,22]. According to Ngwene et al. [23], Si is involved in the solubilization of Pi and the onward transfer of Pi to the host plant. Kushwaha and Kumar [24] identified an acid phosphatase gene in Si that could dissolve organic P. However, it is unclear whether Si establishes a symbiotic relationship with tangor citrus plants, leading to increased physiological responses. The colonization with Si has been demonstrated to promote plant physiological activities and P levels in trifoliate orange seedlings, but it is weaker than AMF [22]. Li et al. [25], however, reported that Si had no effect on the soluble solids and titratable acid content in fruits of the Lane Late navel orange. Diversispora versiformis (Dv) inoculation distinctly increased the amount of fruits and soluble solids–titratable acids ratio in Lane Late navel oranges. This suggests that AMF and Si have varying impacts on host plants. Whether such colonization responses are seen in tangor citrus, however, remain unknown until now.
This study aimed to analyze how two AMF strains and an endophytic fungus (Si) affected the growth, leaf gas exchange, chlorophyll, nitrogen-balance index, chlorophyll fluorescence parameters, P levels, acid phosphatase activity, and the expression of PT and PAP genes in tangor citrus plants.
2. Materials and Methods
2.1. Preparation of Mycorrhizal Inoculums
Two AMF species, Diversispora versiformis (Dv) and Funneliformis mosseae (former Glomus mosseae) (Fm), were obtained from the Institute of Root Biology, Yangtze University (Jingzhou, China) and propagated for three months using Trifolium repens L. in pots. The AMF-colonized roots and growth substrates were collected as the AMF inoculum, containing 18–20 spores/g. Si was donated by the College of Life Science, Yangtze University (Jingzhou, China) and propagated in solid potato–dextrose medium. After the mycelium had grown to cover the medium, it was transferred into an Erlenmeyer flask and 500 mL of distilled water was added to make a spore suspension at 2.72 × 109 colony forming units/mL.
2.2. Plant Culture and Experimental Design
Tangor plants budding on trifoliate orange (2-year-old) in spring were provided by the Fruit and Tea Research Institute, Hubei Academy of Agricultural Sciences (Wuhan, China). In the second year, the plants (with an average root mycorrhizal colonization rate of 5.04 ± 1.26%) were grown in 15 L plastic pots. Each pot received an autoclaved mixture (13.0 kg) composed of soil:sand:commercial substrate (69% charcoal, 25% vermiculite, and 6% perlite) in a volume ratio of 8:2:1. Fungal inoculation was performed at the time of plant transplanting in May 2021, with Dv and Fm inoculated at 800 g/pot and Si inoculated at 1.0 L spore suspension/pot. The no-inoculation group (Control) received an equal amount of autoclaved mycorrhizal fungi and equal amounts of the autoclaved spore suspension of Si.
Following fungal inoculation, the plants were kept indoors for 15 days to acclimatize before being transplanted to a plastic greenhouse on Yangtze University’s West Campus. The plastic greenhouse did not have any equipment for environmental control. A layer of shade netting was applied only on the top of the plastic greenhouse during July‒September, reducing the natural light by approx. 40%. No fertilizer was applied during the experiment.
The experiment included four inoculation treatments: (i) inoculation with Dv; (ii) inoculation with Fm; (iii) inoculation with Si; and (iv) an uninoculated Control in a completely randomized setup, with each treatment replicated five times.
2.3. Variable Determinations
After two years of fungal treatment, the stem diameter and plant height of each plant were determined in June 2023. The roots of the potted plants were collected, and 12 root segments of 1.5 cm length were chosen from each plant and the root fungi stained with 0.05% trypan blue, as described by Yang et al. [22]. The fungal colonization of roots was observed using a biomicroscope (NE610, Ningbo Yongxin Optics Co., Ltd., Ningbo, China), and the root fungal colonization rate was calculated as the percentage of colonized root segments to the detected root segments.
Leaf gas-exchange parameters such as net photosynthetic rate (Pn), intercellular CO2 concentration (Ci), stomatal conductance (Gs), and transpiration rate (Tr) were measured from the top third leaf of the current year’s spring shoots, using a Li-6400 photosynthesizer (Li-COR Inc., Lincoln, NE, USA) on a sunny day at 9:00 am in June 2023. The conditions for the Li-6400 determination were as follows: temperature of 25‒28 °C, light intensity of 820 μmo/m2/s, CO2 concentration of 400 μmol/mol in the leaf compartment, and flow rate of 500 μmol/s. Other parameters such as the leaf chlorophyll index (Chl) and the nitrogen-balance index (Nbi) were measured from the second leaf of the spring shoots with the help of a portable plant polyphenol-chlorophyll meter (Dualex, Force-A, Orsay, France).
The top third leaf of the spring shoots was selected to measure the leaf chlorophyll fluorescence parameters, including Fv/Fm_Lss (maximum efficiency of PSII), QY_max (maximum mass-quantum yield), QY_Lss (minimum quantum yield), and NPQ_Lss (nonphotochemical quenching) using a high-throughput plant fluorescence phenotype-monitoring platform (RAP-FLUO, Wuhan Gufeng Photonics Co., Ltd., Wuhan, China).
The 0.3 g of dried leaf and root samples were digested in H2SO4-H2O2 for the determination of leaf and root P concentrations through a modified molybdenum–antimony anti-reagent colorimetric method at 880 nm with a standard consisting of a 5 μg/mL P solution [26].
Soil acid phosphatase activity was assayed according to the ρ-nitrophenylphosphate method, as described by Zhao and Jiang [27]. Acid phosphatase was extracted with acetate buffer (pH 5.0) and the absorbance was measured at 570 nm. Likewise, root acid phosphatase activity was determined as per the colorimetric method outlined by Tang et al. [28].
The Olsen-P level in soil was determined using an intelligent soil nutrient detector (HM-TYA, Shandong Hengmei Electronic Technology Co., Ltd., Weifang, China) as described in the user’s manual.
Liquid nitrogen was used to grind 50 mg of fresh leaf and root samples into powder. Total RNA from leaves and roots was extracted using the FastPure Plant Total RNA Isolation Kit (Vazyme Biotechnology Co., Ltd., Nanjing, China) and reverse-transcribed into cDNA with the HiSoript I 1st Strand cDNA Synthesis Kit (+gDNA) (Vazyme Biotechnology Co., Ltd.). Specific primers (Supplementary Material Table S1) for three PAP genes and five PT genes were designed based on Genbank sequences (http://www.ncbi.nlm.nih.gov/genbank/, accessed on 31 July 2023). qRT-PCR was used to detect the expression of selected genes. The gene expression was calculated using the 2−∆∆Ct method [29], normalized to the uninoculated Control, with GADPH (glyceraldehyde-3-phosphate dehydrogenase) as the housekeeping gene.
2.4. Data Analysis
All experimental data were analyzed by the one-way analysis of variance with the help of the SPSS 26.0 software (SPSS Inc., Chicago, IL, USA). Percentage data, such as the root fungal colonization rate, were transformed by arcsine. The Duncan’s multiple-range test was employed to compare the significance among treatments at p < 0.05. The Pearson correlation coefficient between fungal colonization and P-associated variables was conducted using the SPSS 26.0 software (SPSS Inc., Chicago, USA).
3. Results
3.1. Response to Root Fungal Colonization Rate
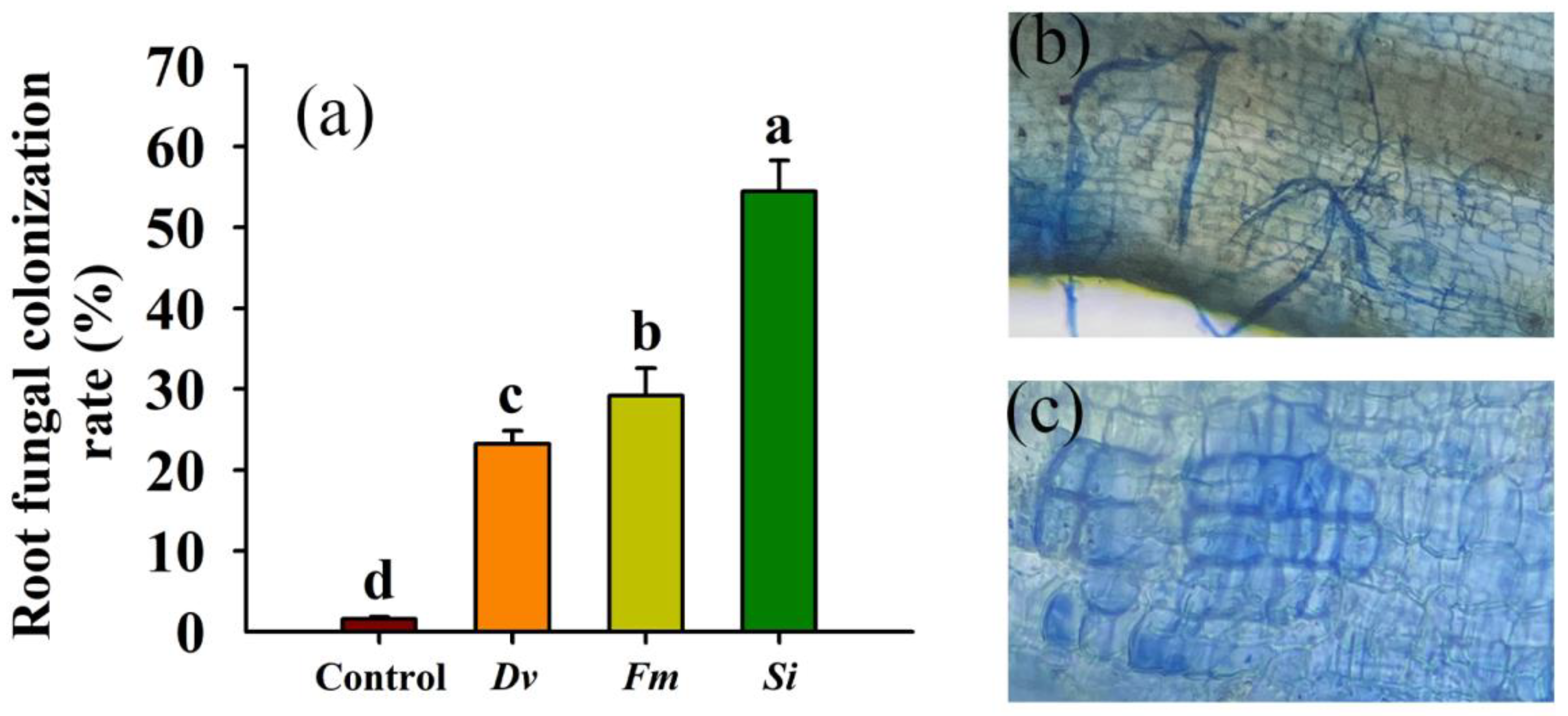
Fungal colonization in the roots of tangor plants inoculated without the tested endophytic fungi was observed as 8.07 ± 1.24% (Figure 1a), which then increased to 23.23 ± 1.6% to 54.46 ± 3.85% after inoculation with the tested endophytic fungi, with a maximum colonization rate of 54.46% in Si-inoculated plants (Figure 1b,c).
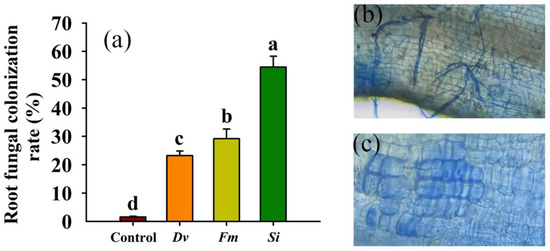
Figure 1.
Changes in the fungal colonization rate (a), mycorrhizal colonization (b), and Serendipita indica colonization (c) in roots of Beni-Madonna tangor (Citrus nanko × C. amakusa) plants. Different letters on the bars (means ± SD, n = 4) indicate significant differences at the 0.05 level. Abbreviations: Control, inoculation without any tested fungus; Dv, Diversipora versiformis; Fm, Funneliformis mosseae; Si, Serendipita indica.
3.2. Plant Growth Response
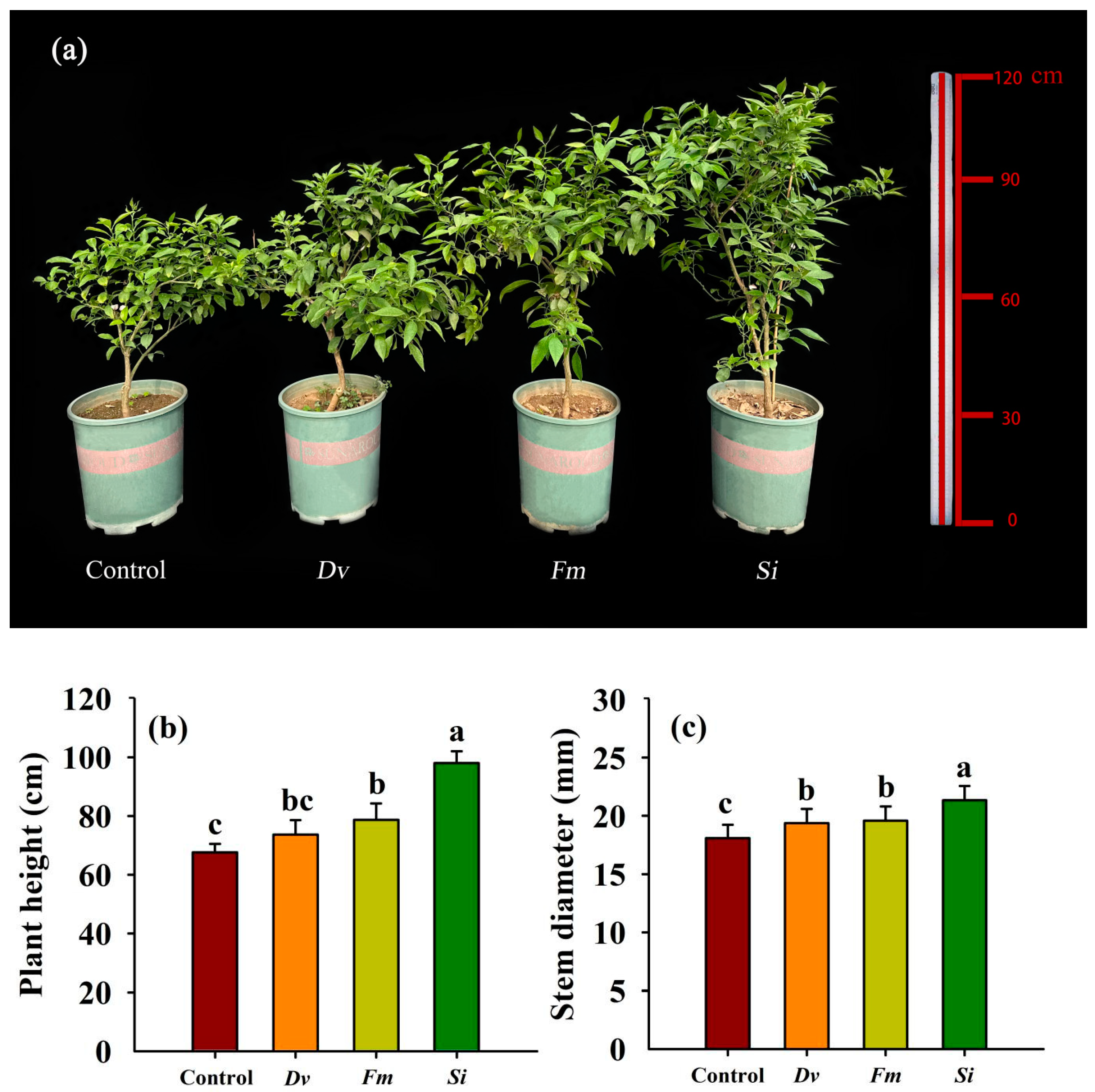
Inoculation with endophytic fungi resulted in significant improvements in tangor plants, depending on the fungal species, with Si having the greatest effect (Figure 2a). Compared with the uninoculated Control, inoculation with Fm and Si, but not Dv, significantly increased the plant height by 16.34% and 44.98%, respectively (Figure 2b). However, inoculation with Dv, Fm, and Si significantly increased the stem diameter by 7.13%, 8.35%, and 18.11%, compared with the uninoculated Control (Figure 2c).
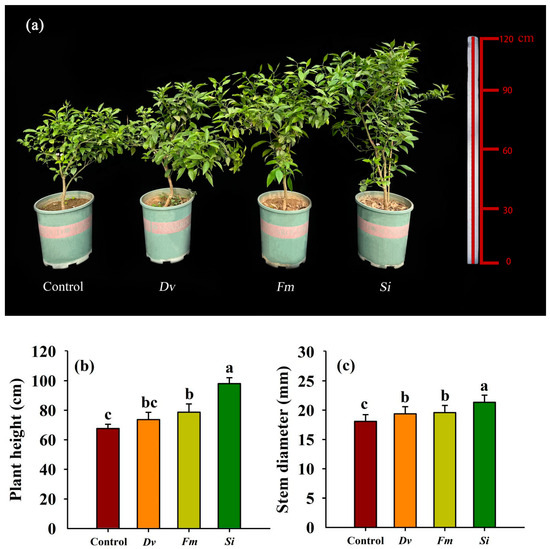
Figure 2.
Changes in plant growth performance (a), plant height (b), and stem diameter (c) in Beni-Madonna tangor (Citrus nanko × C. amakusa) plants inoculated with three endophytic fungi. Different letters on the bars (means ± SD, n = 4) indicate significant differences at the 0.05 level. Abbreviations: Control, inoculation without any tested fungus; Dv, Diversipora versiformis; Fm, Funneliformis mosseae; Si, Serendipita indica.
3.3. Changes in Leaf Chl and Nbi
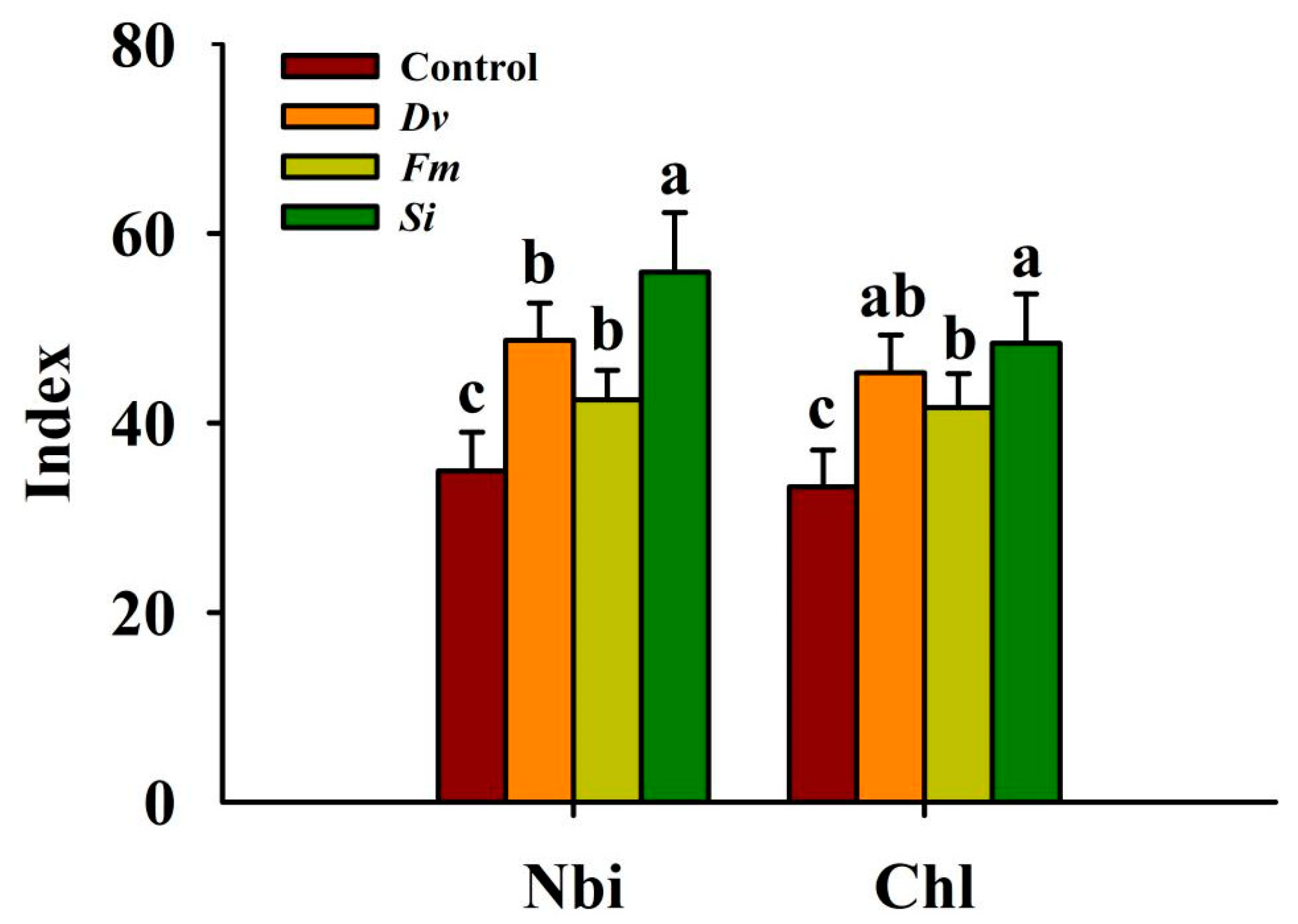
Compared with the uninoculated Control treatment, Si, Dv, and Fm inoculations significantly increased the leaf Nbi by 60.10%, 39.50%, and 21.52%, respectively; similarly, Si, Dv, and Fm inoculations also significantly increased Chl by 45.64%, 36.32%, and 25.20%, respectively, with Si responding more than the other two AMF species (Figure 3).
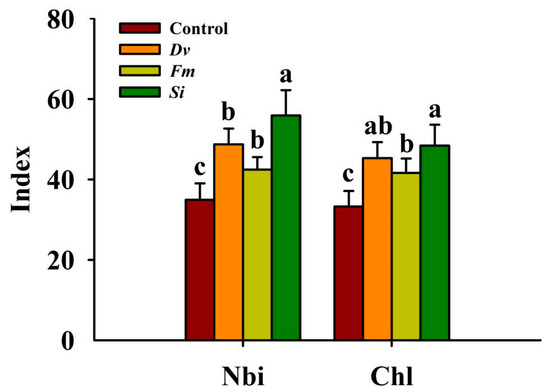
Figure 3.
Changes in the Nbi and Chl in leaves of Beni-Madonna tangor (Citrus nanko × C. amakusa) plants inoculated with three endophytic fungi. Different letters on the bars (means ± SD, n = 4) indicate significant differences at the 0.05 level. Abbreviations: Chl, chlorophyll index; Control, inoculation without any tested fungus; Dv, Diversipora versiformis; Fm, Funneliformis mosseae; Nbi, nitrogen-balance index; Si, Serendipita indica.
3.4. Changes in Leaf Gas-Exchange Parameters
All the three fungal species displayed a significantly varying response in the leaf gas-exchange parameters (Table 1). Dv and Fm treatments significantly reduced leaf Ci compared with the uninoculated Control treatment. Dv inoculation significantly increased leaf Pn, Gs, and Tr by 42.26%, 34.43%, and 23.26%, respectively, and Fm inoculation distinctly increased leaf Pn, Gs, and Tr by 23.23%, 16.39%, and 12.40%, respectively, compared with the Control treatment. Si inoculation, on the other hand, had no significant effect on the leaf Tr but did increase the leaf Pn and Gs by 18.52% and 18.03%, respectively, compared with the uninoculated Control treatment.

Table 1.
Changes in leaf gas-exchange parameters in Beni-Madonna tangor (Citrus nanko × C. amakusa) plants inoculated with three endophytic fungi.
3.5. Changes in Leaf Chlorophyll Fluorescence-Related Parameters
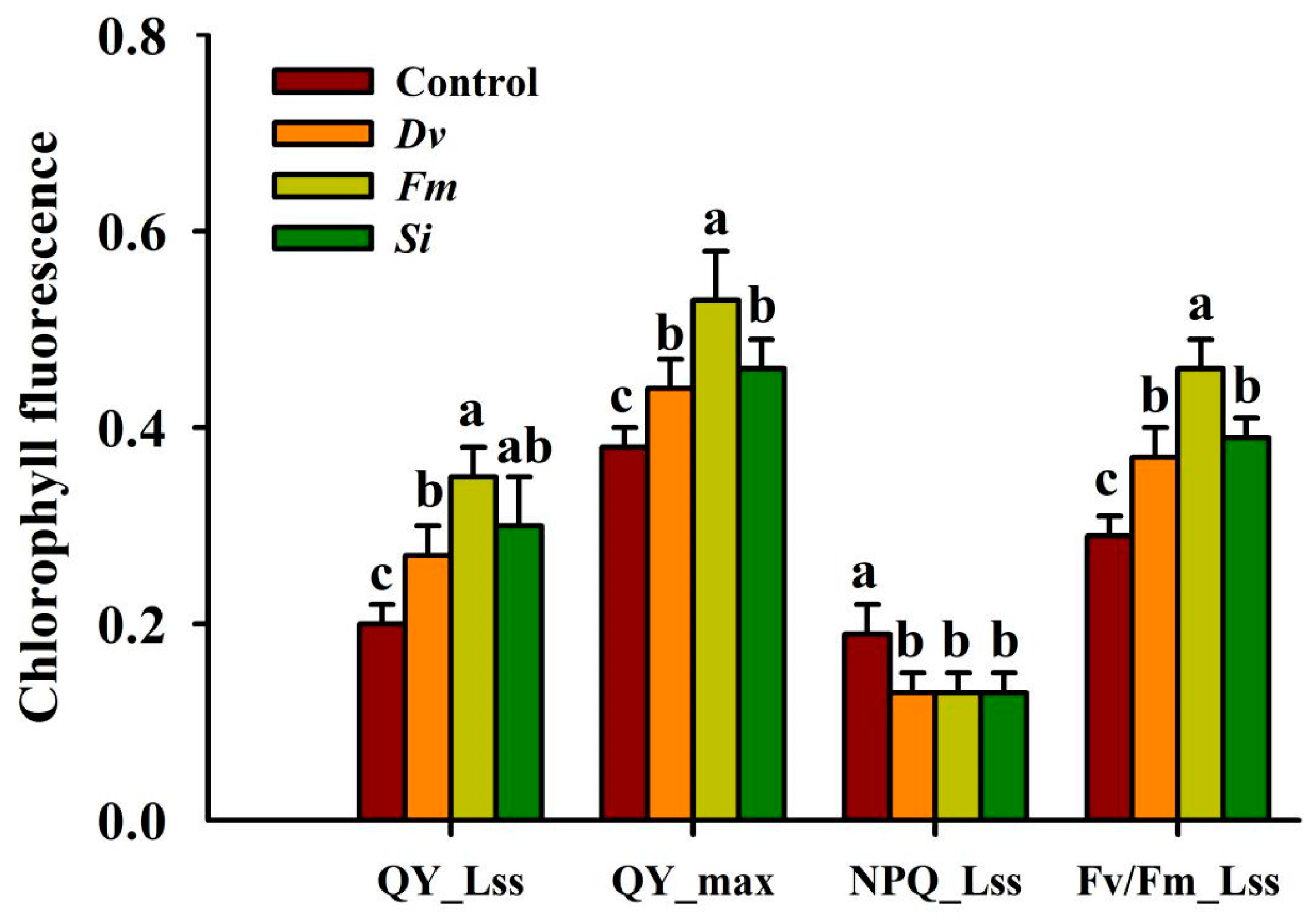
Compared with the uninoculated Control treatment, Fm inoculation significantly raised the QY_Lss, QY_max, and Fv/Fm_Lss by 75.00%, 39.47%, and 58.62%, while Si inoculation distinctly raised the QY_Lss, QY_max, and Fv/Fm_Lss by 50.00%, 21.05%, and 34.48%, and Dv inoculation profoundly increased the QY_Lss, QY_max, and Fv/Fm_Lss by 35.00%, 15.79%, and 27.59%, respectively (Figure 4). NPQ_Lss was significantly reduced following fungal inoculation. Interestingly, Fm inoculation produced a higher magnitude of response in leaf chlorophyll fluorescence parameters than other tested fungi.
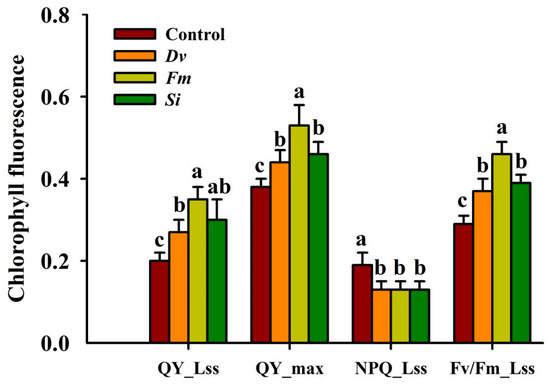
Figure 4.
Changes in the Fv/Fm_Lss (maximum efficiency of PSII), QY_max (maximum mass quantum yield), QY_Lss (minimum quantum yield), and NPQ_Lss (nonphotochemical burst) in leaves of Beni-Madonna tangor (Citrus nanko × C. amakusa) plants inoculated with three endophytic fungi. Different letters on the bars (means ± SD, n = 4) indicate significant differences at the 0.05 levels. Abbreviations: Control, inoculation without any tested fungus; Dv, Diversipora versiformis; Fm, Funneliformis mosseae; Si, Serendipita indica.
3.6. Changes in Leaf and Root P Levels
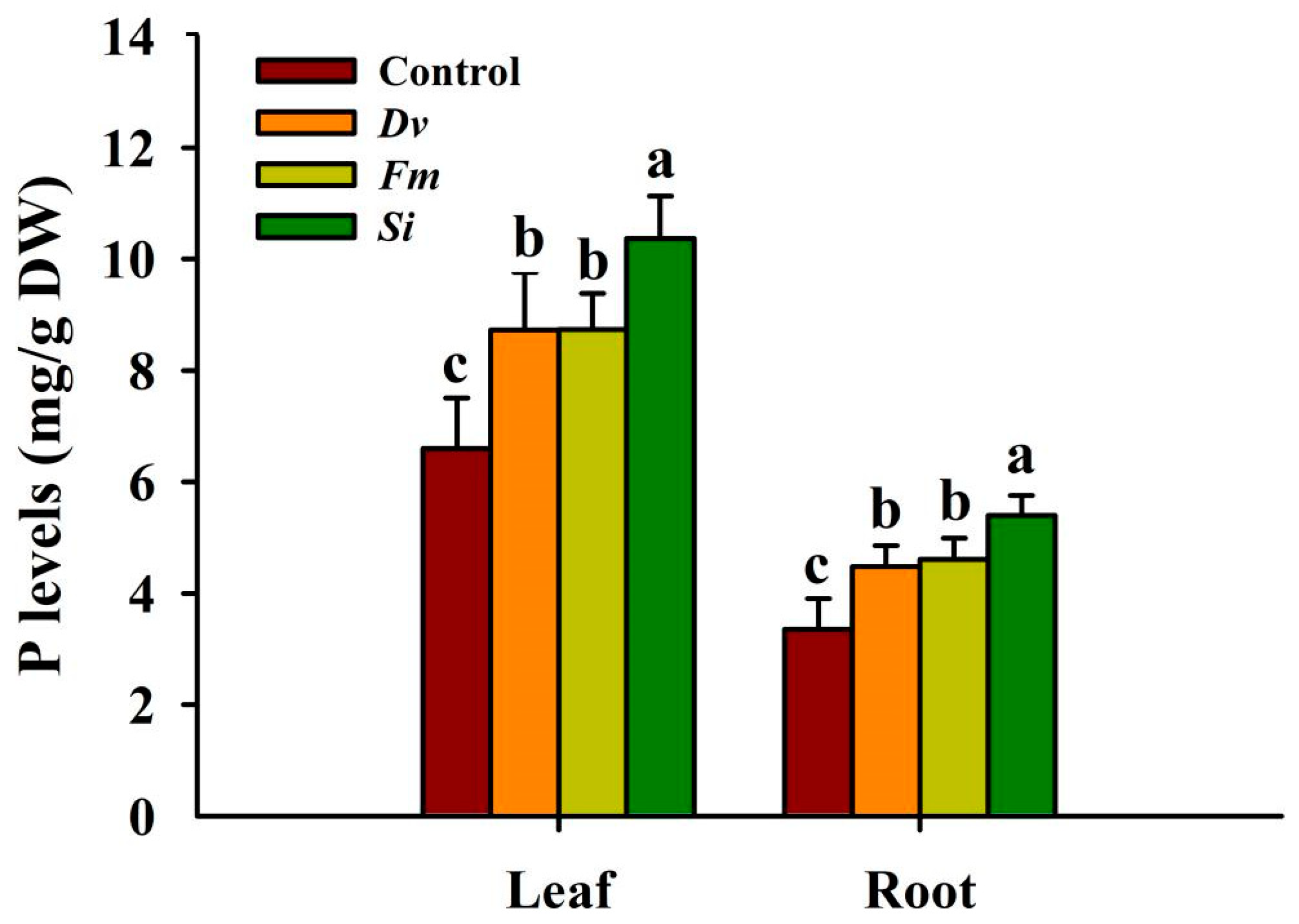
Si, Fm, and Dv inoculations significantly increased P concentrations by 57.05%, 32.43%, and 32.34% in leaves, and by 60.97%, 37.33%, and 33.54% in roots, respectively, compared with the uninoculated Control (Figure 5). However, there was no significant difference between the two AMF treatments with regard to either leaf P or root P concentration.
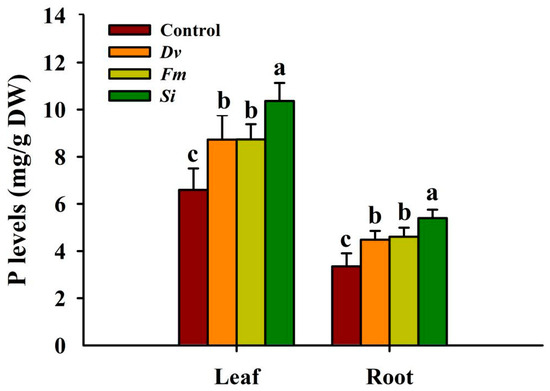
Figure 5.
Changes in leaf and root P levels in Beni-Madonna tangor (Citrus nanko × C. amakusa) plants inoculated with three endophytic fungi. Different letters on the bars (means ± SD, n = 4) indicate significant differences at the 0.05 level. Abbreviations: Control, inoculation without any tested fungus; Dv, Diversipora versiformis; Fm, Funneliformis mosseae; P, phosphorus; Si, Serendipita indica.
3.7. Changes in Root and Soil Acid Phosphatase Activities
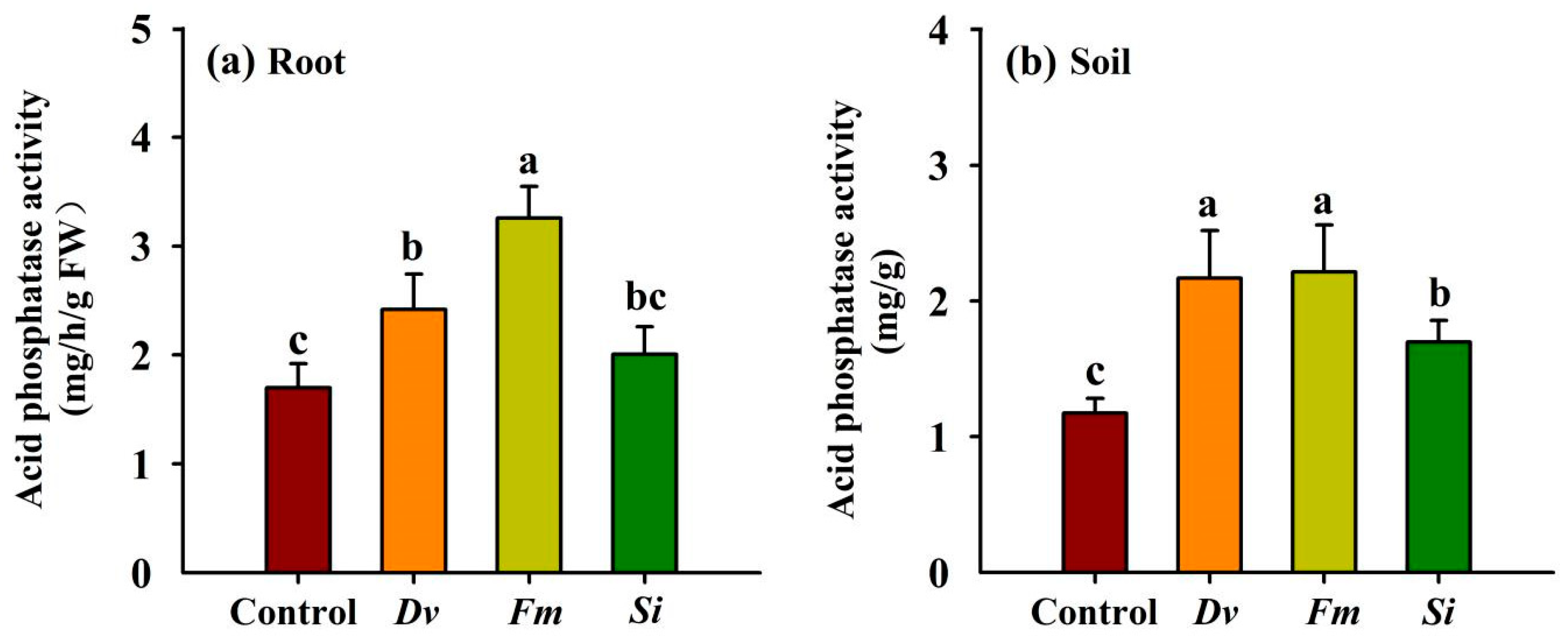
In comparison with the uninoculated Control, Dv and Fm inoculations significantly boosted root acid phosphatase activity by 42.46% and 91.81%, respectively (Figure 6a), without any significant response for Si inoculation. Soil acid phosphatase activity showed a significant increase of 84.75%, 88.59%, and 44.63% with Dv, Fm, and Si inoculations, respectively, compared with the uninoculated Control (Figure 6b).
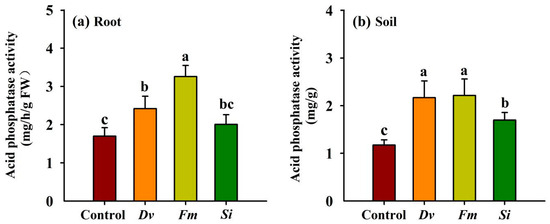
Figure 6.
Changes in root (a) and soil (b) acid phosphatase activities in Beni-Madonna tangor (Citrus nanko × C. amakusa) plants inoculated with three endophytic fungi. Different letters on the bars (means ± SD, n = 4) indicate significant differences at the 0.05 level. Abbreviations: Control, inoculation without any tested fungus; Dv, Diversipora versiformis; Fm, Funneliformis mosseae; Si, Serendipita indica.
3.8. Changes in the Soil Olsen-P Concentration
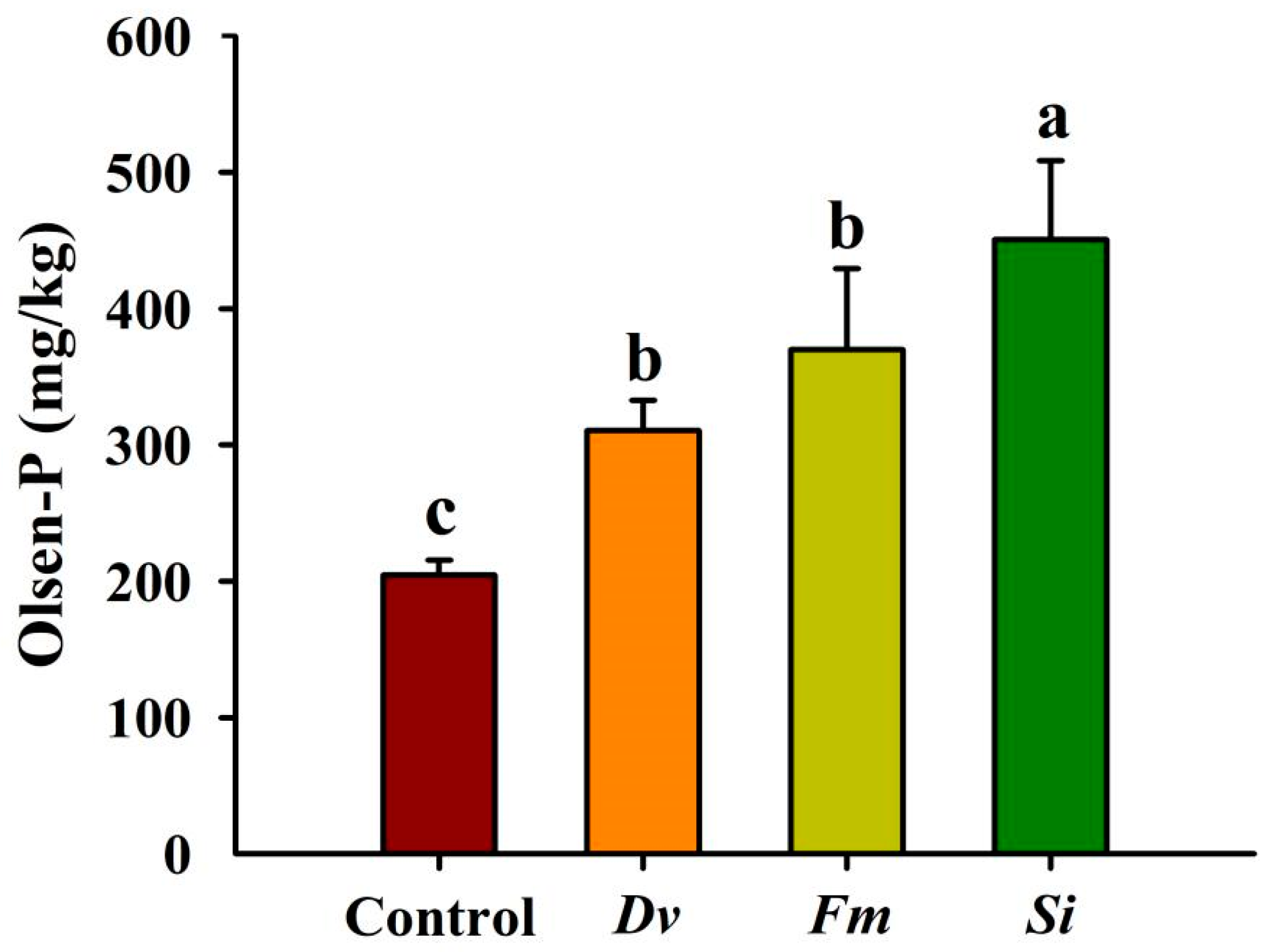
Si, Fm, and Dv inoculations significantly increased the soil Olsen-P concentration by 120.24%, 80.70%, and 51.81%, respectively, compared with the uninoculated Control (Figure 7). Additionally, the increased magnitude of Olsen-P induced by Si exceeded that by the other two AMF treatments.
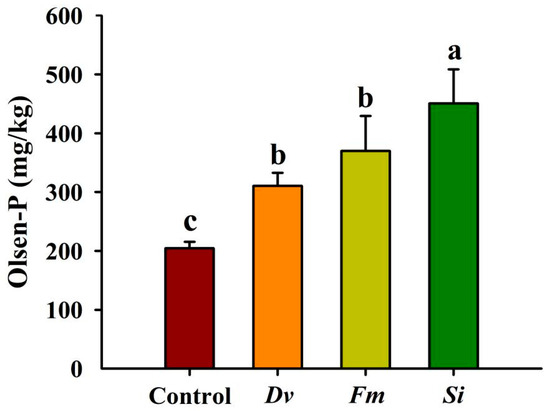
Figure 7.
Changes in soil Olsen-P levels in Beni-Madonna tangor (Citrus nanko × C. amakusa) plants inoculated with three endophytic fungi. Different letters on the bars (means ± SD, n = 4) indicate significant differences at the 0.05 level. Abbreviations: Control, inoculation without any tested fungus; Dv, Diversipora versiformis; Fm, Funneliformis mosseae; P, phosphorus; Si, Serendipita indica.
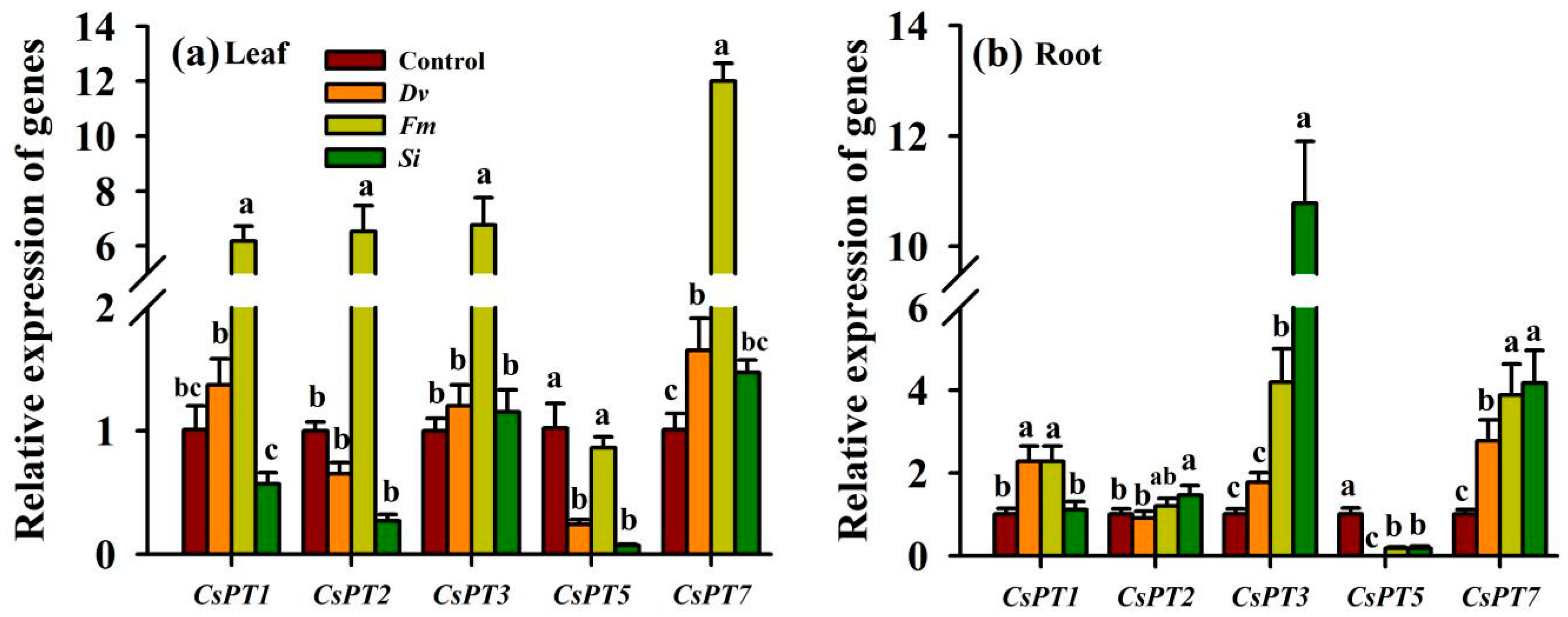
3.9. Changes in CsPT Gene Expression
Leaf and root CsPT gene expression changed significantly after being inoculated with the three endophytic fungi (Figure 8a,b). In leaves, the expression of CsPT1, CsPT2, and CsPT3 was up-regulated by 5.12-, 5.53-, and 5.76-fold, respectively, by Fm compared with the Control (Figure 8a). Nevertheless, the expression of the CsPT5 gene in leaves was reduced by 3.25- and 13.57-fold by Dv and Si inoculation, respectively. Dv and Fm inoculation, but not Si inoculation, significantly increased the expression of leaf CsPT7 by 0.63- and 10.87-fold, respectively, compared with the uninoculated Control.
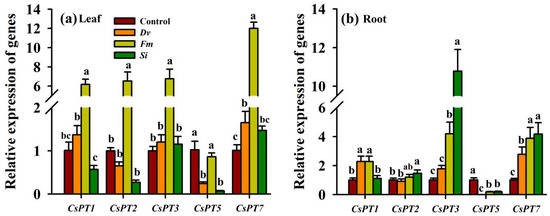
Figure 8.
Changes in CsPT gene expression in the leaf (a) and root (b) of Beni-Madonna tangor (Citrus nanko × C. amakusa) plants inoculated with three endophytic fungi. Different letters on the bars (means ± SD, n = 4) indicate significant differences at the 0.05 level. See Figure 1 for other abbreviations. Abbreviations: Control, inoculation without any tested fungus; Dv, Diversipora versiformis; Fm, Funneliformis mosseae; PT, phosphate transporter; Si, Serendipita indica.
Compared with the uninoculated Control, Dv inoculation significantly up-regulated CsPT1 and CsPT7 expression in the roots by 1.26- and 1.75-fold, while it down-regulated CsPT5 expression in the roots by 32.67-fold, with no significant change in CsPT2 and CsPT3 expression (Figure 8b). Similarly, compared with the uninoculated Control, Fm inoculation also up-regulated CsPT1, CsPT3, and CsPT7 expression in the roots by 1.32-, 3.15-, and 2.84-fold, while it down-regulated CsPT5 expression in the roots by 4.6-fold, with no significant change in CsPT2 expression. Si inoculation, however, significantly up-regulated the expression of CsPT2 (0.46 fold), CsPT3 (9.67 fold), and CsPT7 (3.61 fold) and down-regulated the expression of CsPT5 (4.32 fold) in the roots compared with the uninoculated Control, which was accompanied by no change in root CsPT1 expression.
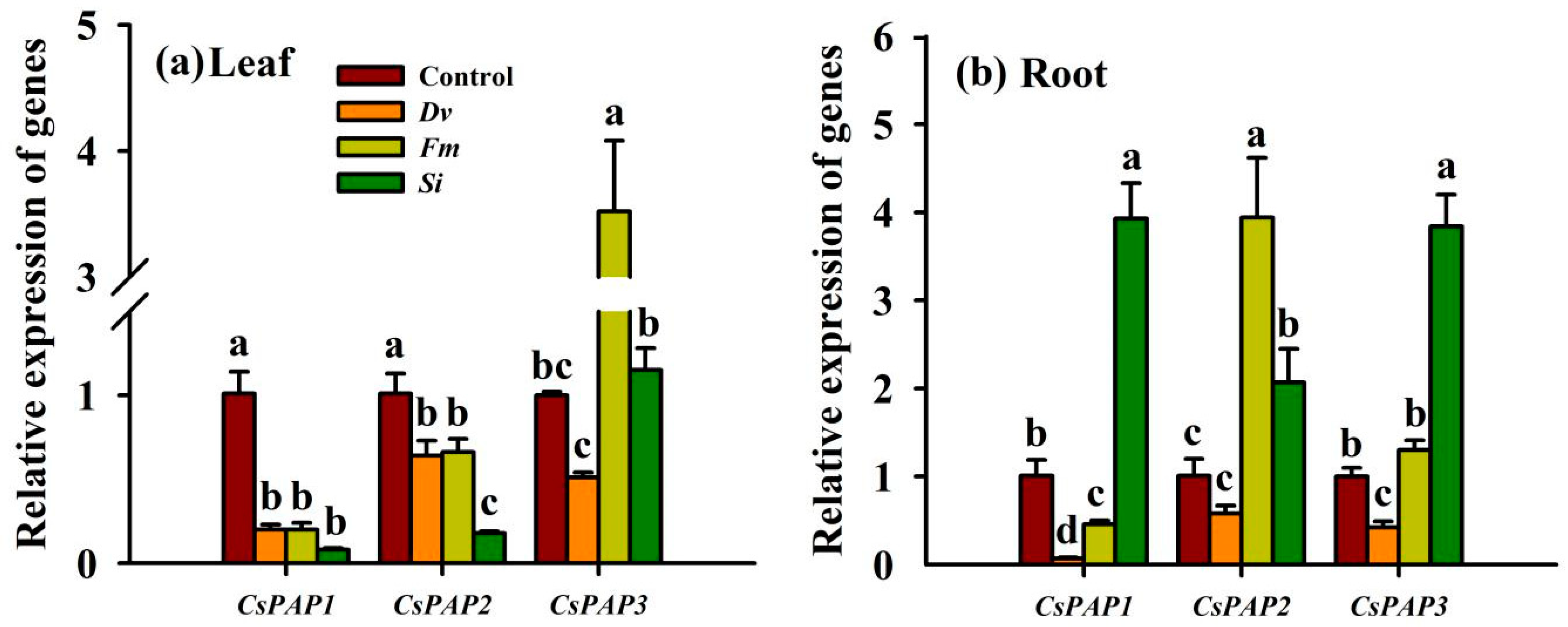
3.10. Changes in CsPAP Gene Expression
Expression of CsPAP was significantly affected to varying degrees in response to inoculation with the three endophytic fungi (Figure 9). In leaves, compared with the uninoculated Control treatment, endophytic fungal inoculations collectively decreased the expression of CsPAP and CsPAP2 by 4.05‒11.63 folds and 0.53‒4.61 folds, respectively (Figure 9a). Out of the three fungi, only Fm inoculation up-regulated the expression of CsPAP3 in the leaves, by 2.52-fold. In the roots, Dv inoculation produced a significant reduction in the expression of CsPAP1 of 13.43-fold and of CsPAP3 genes of 1.38-fold, with no significant response on the expression of CsPAP2, compared with the uninoculated Control treatment (Figure 9b). Fm inoculation significantly increased the expression of CsPAP2 (2.93-fold) in the roots, inhibited the expression of CsPAP1 (1.2-fold) in the roots, but had no effect on the expression of CsPAP3 in the roots. Si inoculation significantly increased the expression of CsPAP1, CsPAP2, and CsPAP3 in the roots by 2.89-, 1.05-, and 2.84-fold, respectively, compared with the uninoculated Control treatment.
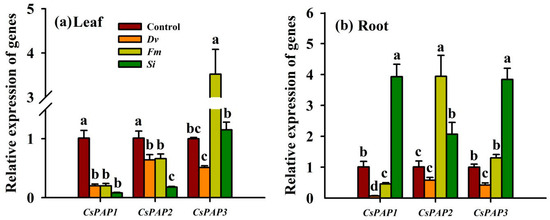
Figure 9.
Changes in CsPAP1‒3 gene expression in the leaf (a) and root (b) of Beni-Madonna tangor (Citrus nanko × C. amakusa) plants inoculated with three endophytic fungi. Different letters on the bars (means ± SD, n = 4) indicate significant differences at the 0.05 level. Abbreviations: Control, inoculation without any tested fungus; Dv, Diversipora versiformis; Fm, Funneliformis mosseae; PAP, purple acid phosphatase; Si, Serendipita indica.
3.11. Correlation Analysis
Correlation studies revealed that the root fungal colonization rate was significantly and positively correlated with soil Olsen-P, leaf P, root P, root CsPT2, CsPT3, CsPT7, CsPAP1, and CsPAP3 expression, while it was negatively correlated with root CsPT5 expression (Table 2). Root P was significantly and positively correlated with soil Olsen-P, root CsPT2, CsPT3, CsPT7, CsPAP1, and CsPAP3 expression but negatively correlated with root CsPT5 expression. On the other hand, leaf P showed a significantly positive correlation with soil Olsen-P and a negative correlation with leaf CsPT5, CsPAP1, and CsPAP2 expression.
Table 2.
The correlation coefficients between the root fungal colonization rate and the P-associated variables.
4. Discussion
Citrus spp. depend heavily on mycorrhizal symbiosis [17,25]. In this study, two AMF strains (Dv and Fm) and one endophytic fungus (Si) were found to colonize the roots of Beni-Madonna tangor citrus plants to varying proportions, with Si having the greatest root colonization rate (54.46%). Cheng et al. [30] also observed an increase in the root fungal colonization rate of Newhall navel orange plants following field inoculation with Dv, D. spurca and Si, with the Si colonization rate being significantly higher than that of AMF. The root colonization rate of Si in field Lane Late navel orange plants, however, was lower than that of the other three AMF strains [25]. Such a differential response among the endophytic fungi strengthens the dependence of citrus species on mycorrhizal species, while the intrinsic molecular mechanisms underlying the differences between AMF and Si with respect to citrus plant recognition and colonization remain to be urgently investigated.
The changes in the growth response of endophyte-inoculated plants are the result of mutualistically defined symbiotic benefits and the host-dependence on the fungus [31]. In our study, the inoculation response to Dv, Fm, and Si was associated with improvements in the plant height and stem diameter of tangor plants, and the growth improvement was consistent with their root colonization rates, strongly demonstrating the association of endophytic fungal colonization rate with the host plant growth response [32]. Among the three fungi used, inoculation with Si produced the best plant growth response. This improvement was correlated with their promotion of gas exchange and P acquisition. Endophytic fungi are also known as a source of bioactive secondary metabolites, such as auxins and gibberellins, which collectively promote plant growth [33,34]. Yang et al. [22] earlier reported a weaker growth-promoting response of Si than AMF in potted trifoliate orange, implying a difference in citrus growth response between AMF and Si.
The fungi are not able to photosynthesize, and thus they sustain their own growth and development by exchanging substances with the host plant, which regulates host gas exchange [35]. In this study, all endophytic fungal treatments elevated the Pn, Gs, and Tr of leaves, with the exception of Tr, which did not differ significantly between the Control treatment and Si treatment. This demonstrated that root-associated endophytic fungi, particularly AMF, enhance the photosynthetic capability of citrus plants by increasing P levels, which can accelerate leaf gas exchange [36]. In addition, endophytic fungi promote the host plant’s acquisition of several nutrients such as N, Mg, and Fe, which are favorable for chlorophyll synthesis [37]. It is, hence, concluded that endophytic fungi improve the host’s photosynthetic capacity through increased Chl and N levels. Li et al. [25] further reported that Si inoculation predominantly affected sucrose synthesis in citrus fruits by up-regulating the expression of sucrose phosphate synthase (SPS) genes, whereas AMF predominantly affected sucrose synthesis, catabolism, and transport in citrus fruits by regulating the expression levels of associated genes (SWEET, VINV, and NINV). This revealed significant differences between AMF and Si in the regulation of sugar metabolism.
Chlorophyll fluorescence reflects the whole process of absorption, utilization, transfer, and dissipation of light energy regulated through PSI and PSII [38]. In this study, all fungal inoculations increased the QY_Lss, QY_max, and Fv/Fm_Lss, which is similar to the findings of Cui et al. [39] and Khalvandi et al. [40]. Among the three tested fungal species, Fm displayed the greatest significant effect on the QY_Lss, QY_max, and Fv/Fm_Lss. Fv/Fm_Lss is an important indicator in determining photoinhibition [5]. In our study, all fungal inoculation treatments significantly increased the Fv/Fm_Lss, suggesting that symbiotic fungal-inoculated citrus plants suffered less photoinhibition. Endophytic fungi also significantly reduced the NPQ_Lss, resulting in less heat dissipation [41], facilitating an improved photochemical efficiency (QY_max). As a result, photosynthetic organs in inoculated plants can convert absorbed light energy into biochemical energy more efficiently, thereby maintaining the photosynthetic efficiency and quantum yield of PSII in inoculated plants [5]. Therefore, citrus plants inoculated with endophytic fungi show low photoinhibition and heat dissipation while exhibiting a high photosynthetic efficiency and quantum yield of PSII.
Soil phosphatase promotes the decomposition of soil organic Pi into inorganic Pi, thereby allowing roots to absorb soil P more efficiently [42]. In the present study, the soil acid phosphatase activity of plants inoculated with endophytic fungi was considerably raised, with AMF showing a greater increase than Si-inoculated plants. Similar results were reported in Newhall navel orange plants inoculated with AMF and Si in our earlier studies as well in [30]. Si colonization promoted P accumulation in trifoliate orange seedlings, although the promotion was lower than that by AMF [22]. On the other hand, the response with Dv and Fm, but not Si, also conferred improvements in the root acid phosphatase activity. Hassan [43] observed faster growth of Teucrium polium plants treated with Penicillium crustosum due to increased soil Pi solubilization. Previously, an endophytic fungus, Phialocephala fortinii, also decomposed soil insoluble P, thus promoting the growth of Asparagus officinalis plants [44]. Endophytic fungi like AMF and Si are known to increase the release of acid phosphatase from the host roots into the soil to dissolve organic or insoluble P, thus resulting in Pi accumulation and the subsequent absorption by the host roots [32]. This study also observed higher leaf and root P concentrations in inoculated versus uninoculated plants. Nevertheless, Si-inoculated tangor plants exhibited higher soil Olsen-P levels and lower soil and root phosphatase activities than AMF-inoculated plants. This may be due to the formation of polyphosphates into the AMF mycelia, thus affecting phosphatase activity. Another explanation could also be because the expression of CsPAP1 and CsPAP3 gene in roots was up-regulated to a relatively higher extent under Si versus AMF conditions. The root-associated endophytic fungi also generate a well-developed network of external hyphae on the surface of host roots, expanding the root–soil contact area, shortening the nutrient transport distance, and thus contributing toward the greater uptake of P by the host plant [45,46]. Such an increase in P acquisition on endophytic fungus-inoculated citrus plants suggests that the input of inorganic P fertilizer in citrus orchards can be reduced, which would provide a path for sustainable citrus development and green fruit production.
In the present study, both AMF and Si suppressed leaf CsPAP1‒3 expression. In roots, Si promoted CsPAP1‒3 expression, Fm only up-regulated CsPAP2 expression, and Dv has no effect on the CsPAP1‒3 expression. This suggested that Si was more effective than AMF at regulating the expression of CsPAPs in roots. Many plant-derived PAPs have acid phosphatase activity, capable of hydrolyzing organic Pi into inorganic Pi for plant uptake and utilization [10]. Koide and Kabir [47] demonstrated that mycorrhizal extraradical hyphae produced phosphatases both intracellularly and extracellularly. Based on these findings, it is suggested that Si aids in promoting acid phosphatase release primarily by up-regulating host PAP expression in roots. Nevertheless, AMF increased soil phosphatase activity primarily via its own phosphatase release, which remains to be further confirmed.
In this study, Dv up-regulated the expression of CsPT1 and CsPT7 in roots, Fm up-regulated the expression of CsPT1, CsPT3, and CsPT7 in roots, and Si up-regulated the expression of CsPT2, CsPT3, and CsPT7 in roots. Fm inoculation significantly increased the expression of four of the five CsPTs (excluding CsPT5) in leaves. These findings demonstrated the effectiveness of Fm amongst the three investigated fungi. Enhanced P uptake and further transport by AMF was concentrated in the roots, since the mycorrhizal P-uptake pathway is located around the arbuscule membranes. The effects of different endophytic fungi on the expression of host PTs vary as per plant species and PT homologous genes. The expression of JrPTs by D. spurca on walnut leaves was affected by substrate P levels [48]. This study also observed that CsPT7 was up-regulated by nearly all three endophytic fungi in both leaves and roots, showing that CsPT7 may respond specifically to endophytic fungi, especially AMF. It is necessary to explore how CsPT7 participates in P transport in root cortical cells containing arbuscules.
5. Conclusions
Dv, Fm, and Si collectively colonized the roots of Beni-Madonna tangor plants and improved plant growth performance, along with increasing the photosynthetic efficiency, gas exchange, and chlorophyll levels. In addition, endophytic fungi also improved plant P levels, but the strategy differed between AMF and Si: Si up-regulated the expression of root CsPAP1‒3, CsPT2, CsPT3, and CsPT7; AMF increased the expression of root CsPT1 and CsPT7 as well as root phosphatase activity. Overall, Si had a prominent effect on plant growth and P acquisition, while Fm presented a significant effect in improving the photosynthetic efficiency and gas exchange. Since Si and AMF display different responses in tangor plants at the molecular level, the dual use of both kinds of fungi in a consortium may have additive effects, which needs to be confirmed by further studies. Additional observations on changes in the fruiting traits as well as fruit quality are needed. In addition, a long-term endophytic-fungal-treated change in citrus plants is required, which will allow for a comprehensive evaluation of ecological changes in the orchard as well as long-term sustainable citrus development. Given the property of Si to be propagated in vitro on a large scale, it is more advantageous for use in Beni-Madonna tangor plants for healthy growth.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae10020145/s1, Table S1: Primer sequences of genes screened by qRT-PCR.
Author Contributions
Conceptualization, Q.-S.W.; Methodology, Y.L., W.-J.W. and Z.-Y.R.; Formal analysis, W.-J.W.; Investigation, Y.L., M.-Y.Z., Z.-Y.R., T.-Z.Z., X.-P.T. and Y.H.; Resources, W.-J.W. and Q.-S.W.; Data curation, Y.L., M.-Y.Z. and Z.-Y.R.; Writing—original draft, Y.L.; Writing—review and editing, M.D.A., S.K.M., A.K.S. and Q.-S.W.; Visualization, Y.L.; Supervision, Q.-S.W.; Funding acquisition, Q.-S.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Hubei Agricultural Science and Technology Innovation Action Project (Hubei Nongfa [2018] No. 1) and the 2023 Undergraduate Innovation and Entrepreneurship Training Program of Yangtze University (Yz2023392). The authors extend their appreciation to the Princess Nourah bint Abdulrahman University Researchers Supporting Project number (PNURSP2024R355), Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
The authors extend their appreciation to the Princess Nourah bint Abdulrahman University Researchers Supporting Project number (PNURSP2024R355), Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ushijima, K.; Mizuta, K.; Otomo, S.; Ogaki, K.; Sanada, Y.; Hirata, Y.; Ihara, Y.; Urahashi, T.; Imai, Y.; Fujimura, A. Increased tacrolimus blood concentration by Beni-Madonna—A new hybrid citrus cultivar categorized as ‘Tangor’, in a liver transplant patient: Likely furanocoumarin-mediated inhibition of CYP3A4 or P-glycoprotein. Br. J. Clin. Pharmacol. 2018, 84, 2933–2935. [Google Scholar] [CrossRef]
- Huang, Z.D.; Wang, P.; Xu, J.G.; Lu, L.M.; Chen, G.Q.; Wen, M.X.; Lin, M. Relationship between fruit quality and nutrients in soil and leaves of ‘Hongmeiren’ citrus hybrid cultivated in eastern Zhejiang province. J. Fruit Sci. 2020, 37, 88–97. [Google Scholar] [CrossRef]
- Zhang, F.F.; Zhang, J.J.; Xia, H.L.; Zhu, H.Z. Simultaneous determination of 12 kinds of flavonoids in different parts of ‘Hongmeiren’ citrus by high performance liquid chromatography. J. Food Saf. Qual. 2022, 13, 2267–2273. [Google Scholar] [CrossRef]
- Lambers, H. Phosphorus acquisition and utilization in plants. Ann. Rev. Plant Biol. 2022, 73, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Wang, F.; Wang, Y.; Lin, R.; Wang, Z.; Mao, C. Molecular mechanisms and genetic improvement of low-phosphorus tolerance in rice. Plant Cell Environ. 2023, 46, 1104–1119. [Google Scholar] [CrossRef]
- Shao, X.; Zhang, J.; Guo, L. Assessing the feasibility of Olsen P and P sorption parameters in acidic soils. Commun. Soil Sci. Plant Anal. 2017, 48, 2049–2060. [Google Scholar] [CrossRef]
- Raiesi, T.; Moradi, B.; Mousavi, S.M. Alterations of P fractions and some biochemical features in rhizosphere soil induced by the root activities of citrus rootstocks with different P acquisition efficiency. Eurasian Soil Sci. 2022, 55, 212–220. [Google Scholar] [CrossRef]
- Wang, X.X.; Hoffland, E.; Feng, G.; Kuyper, T.W. Arbuscular mycorrhizal symbiosis increases phosphorus uptake and productivity of mixtures of maize varieties compared to monocultures. J. Appl. Ecol. 2020, 57, 2203–2211. [Google Scholar] [CrossRef]
- Hurley, B.A.; Tran, H.T.; Marty, N.J.; Park, J.; Snedden, W.A.; Mullen, R.T.; Plaxton, W.C. The dual-targeted purple acid phosphatase isozyme AtPAP26 is essential for efficient acclimation of Arabidopsis to nutritional phosphate deprivation. Plant Physiol. 2010, 153, 1112–1122. [Google Scholar] [CrossRef]
- Olczak, M.; Morawiecka, B.; Watorek, W. Plant purple acid phosphatases-genes, structures and biological function. Acta Biochim. Pol. 2003, 50, 1245–1256. [Google Scholar] [CrossRef]
- Nagarajan, V.K.; Jain, A.; Poling, M.D.; Lewis, A.J.; Raghothama, K.G.; Smith, A.P. Arabidopsis Pht1;5 mobilizes phosphate between source and sink organs and influences the interaction between phosphate homeostasis and ethylene signaling. Plant Physiol. 2011, 156, 1149–1163. [Google Scholar] [CrossRef]
- Akiyama, K.; Matsuzaki, K.I.; Hayashi, H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 2005, 435, 824–827. [Google Scholar] [CrossRef]
- Bennett, A.E.; Groten, K. The costs and benefits of plant–arbuscular mycorrhizal fungal interactions. Ann. Rev. Plant Biol. 2022, 73, 649–672. [Google Scholar] [CrossRef]
- Bourazza, M.; Chetto, O.; Talha, A.; Douira, A.; Benyahia, H. Diversity of arbuscular mycorrhizal fungi in the rhizosphere of four citrus genotypes. In International Conference on Advanced Intelligent Systems for Sustainable Development, AI2SD 2022; Kacprzyk, J., Ezziyyani, M., Balas, V.E., Eds.; Lecture Notes in Networks and Systems; Springer: Cham, Switzerland, 2023; Volume 713, pp. 623–636. [Google Scholar] [CrossRef]
- Manresa-Grao, M.; Pastor-Fernández, J.; Sanchez-Bel, P.; Jaques, J.A.; Pastor, V.; Flors, V. Mycorrhizal symbiosis triggers local resistance in citrus plants against spider mites. Front. Plant Sci. 2022, 13, 867778. [Google Scholar] [CrossRef] [PubMed]
- Ortas, I.; Demirbas, A.; Akpinar, C. Time period and nutrient contents alter the mycorrhizal responsiveness of citrus seedlings. Eur. J. Hortic. Sci. 2018, 83, 72–80. [Google Scholar] [CrossRef]
- Zou, Y.N.; Zhang, F.; Srivastava, A.K.; Wu, Q.S.; Kuča, K. Arbuscular mycorrhizal fungi regulate polyamine homeostasis in roots of trifoliate orange for improved adaptation to soil moisture deficit stress. Front. Plant Sci. 2021, 11, 600792. [Google Scholar] [CrossRef]
- Liu, R.C.; Yang, L.; Zou, Y.N.; Wu, Q.S. Root-associated endophytic fungi modulate endogenous auxin and cytokinin levels to improve plant biomass and root morphology of trifoliate orange. Hortic. Plant J. 2023, 9, 463–472. [Google Scholar] [CrossRef]
- Cozzolino, V.; Di Meo, V.; Piccolo, A. Impact of arbuscular mycorrhizal fungi applications on maize production and soil phosphorus availability. J. Geochem. Explor. 2013, 129, 40–44. [Google Scholar] [CrossRef]
- Mohandas, S. Arbuscular mycorrhizal fungi benefit mango (Mangifera indica L.) plant growth in the field. Sci. Hortic. 2012, 143, 43–48. [Google Scholar] [CrossRef]
- Ren, A.T.; Lu, W.H.; Yang, J.J.; Liu, H.L.; Ma, C.H. Effects of arbuscular mycorrhizal fungi on growth and phosphorus utilization of alfalfa under different phosphorus levels. Chin. J. Grassl. 2014, 36, 72–78. [Google Scholar]
- Yang, L.; Zou, Y.N.; Tian, Z.H.; Wu, Q.S.; Kuča, K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci. Hortic. 2021, 277, 109815. [Google Scholar] [CrossRef]
- Ngwene, B.; Boukail, S.; Söllner, L.; Franken, P.; Andrade-Linares, D.R. Phosphate utilization by the fungal root endophyte Piriformospora indica. Plant Soil 2016, 405, 231–241. [Google Scholar] [CrossRef]
- Kushwaha, A.S.; Kumar, M. An effective in-gel assay protocol for the assessment of acid phosphatase (ACPase) isoform expression in the fungus Serendipita indica. 3 Biotech 2022, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.S.; Srivastava, A.K.; Zou, Y.N.; Wu, Q.S. Field inoculation responses of arbuscular mycorrhizal fungi versus endophytic fungi on sugar metabolism associated changes in fruit quality of Lane late navel orange. Sci. Hortic. 2023, 308, 111587. [Google Scholar] [CrossRef]
- Li, P.; Li, M.M.; Wang, H.; Jin, M.S.; Yu, G.H.; Xing, G.F. Determination of phosphorus content in different foxtail millet tissues by improved molybdenum antimony resistance colorimetry. Acta Laser Biol. Sin. 2020, 29, 237–244. [Google Scholar]
- Zhao, L.P.; Jiang, Y. Determination of soil phosphatase activity. Chin. J. Soil Sci. 1986, 17, 138–141. [Google Scholar]
- Tang, H.L.; Li, X.Q.; Zu, C.; Zhang, F.S.; Shen, J.B. Spatial distribution and expression of intracellular and extracellular acid phosphatases of cluster roots at different developmental stages in white lupin. J. Plant Physiol. 2013, 170, 1243–1250. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Cheng, X.F.; Xie, M.M.; Li, Y.; Liu, B.Y.; Liu, C.Y.; Wu, Q.S.; Kuča, K. Effects of field inoculation with arbuscular mycorrhizal fungi and endophytic fungi on fruit quality and soil properties of Newhall navel orange. Appl. Soil Ecol. 2022, 170, 104308. [Google Scholar] [CrossRef]
- Mayerhofer, M.S.; Kernaghan, G.; Harper, K.A. The effects of fungal root endophytes on plant growth: A meta-analysis. Mycorrhiza 2013, 23, 119–128. [Google Scholar] [CrossRef]
- Poveda, J.; Eugui, D.; Abril-Urías, P.; Velasco, P. Endophytic fungi as direct plant growth promoters for sustainable agricultural production. Symbiosis 2021, 85, 1–19. [Google Scholar] [CrossRef]
- Waqas, M.; Khan, A.L.; Kamran, M.; Hamayun, M.; Kang, S.M.; Kim, Y.H.; Lee, I.J. Endophytic fungi produce gibberellins and indoleacetic acid and promotes host-plant growth during stress. Molecules 2012, 17, 10754–10773. [Google Scholar] [CrossRef] [PubMed]
- Baron, N.C.; Rigobelo, E.C. Endophytic fungi: A tool for plant growth promotion and sustainable agriculture. Mycology 2022, 13, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.L.; Li, C.Y.; Wang, C.R.; Wang, Y.; Liu, J.; Chang, S.; Miao, Y.; Dang, T.H. Mechanism of arbuscular mycorrhizal fungi promoting plant uptake of soil phosphorus. J. Soil Water Conserv. 2019, 33, 10–20. [Google Scholar] [CrossRef]
- Pang, J.Y.; Zhao, H.X.; Bansal, R.; Bohuon, E.; Lambers, H.; Ryan, M.H.; Siddique, K.H.M. Leaf transpiration plays a role in phosphorus acquisition among a large set of chickpea genotypes. Plant Cell Environ. 2018, 41, 2069–2079. [Google Scholar] [CrossRef]
- Bondada, B.R.; Syvertsen, J.P. Leaf chlorophyll, net gas exchange and chloroplast ultrastructure in citrus leaves of different nitrogen status. Tree Physiol. 2003, 23, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.W.; Pan, Y.H.; Wang, M.; Liu, T.H.; Meng, X.S.; Guo, S.W. The effects of different nitrogen forms on chlorophyll fluorescence and photosystem II in Lonicera japonica. J. Plant Growth Regul. 2023, 42, 4106–4117. [Google Scholar] [CrossRef]
- Cui, L.; Guo, F.; Tang, Z.H.; Zhang, J.L.; Yang, S.; Wang, J.G.; Meng, J.J.; Geng, Y.; Li, X.G.; Wan, S.B. The effect of Funneliformis mosseae on the fluorescence parameters and physiological indexes of continuously cropped peanut leaves. Chin. J. Oil Crops Sci. 2020, 42, 851. [Google Scholar] [CrossRef]
- Khalvandi, M.; Amerian, M.; Pirdashti, H.; Keramati, S. Does co-inoculation of mycorrhiza and Piriformospora indica fungi enhance the efficiency of chlorophyll fluorescence and essential oil composition in peppermint under irrigation with saline water from the Caspian Sea? PLoS ONE 2021, 16, e0254076. [Google Scholar] [CrossRef]
- Shin, Y.K.; Jo, J.S.; Cho, M.C.; Yang, E.Y.; Ahn, Y.K.; Lee, J.G. Application of chlorophyll fluorescence parameters to diagnose salinity tolerance in the seedling of tomato genetic resources. J. Bio-Environ. Control 2021, 30, 165–173. [Google Scholar] [CrossRef]
- Margalef, O.; Sardans, J.; Maspons, J.; Molowny-Horas, R.; Fernández-Martínez, M.; Janssens, I.A.; Richter, A.; Ciais, P.; Obersteiner, M.; Peñuelas, J. The effect of global change on soil phosphatase activity. Glob. Chang. Biol. 2021, 27, 5989–6003. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.E.D. Plant growth-promoting activities for bacterial and fungal endophytes isolated from medicinal plant of Teucrium polium L. J. Adv. Res. 2017, 8, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Surono; Narisawa, K. The dark septate endophytic fungus Phialocephala fortinii is a potential decomposer of soil organic compounds and a promoter of Asparagus officinalis growth. Fungal Ecol. 2017, 28, 1–10. [Google Scholar] [CrossRef]
- Etesami, H. Enhanced phosphorus fertilizer use efficiency with microorganisms. In Nutrient Dynamics for Sustainable Crop Production; Meena, R., Ed.; Springer Nature: Singapore, 2020; pp. 215–245. [Google Scholar] [CrossRef]
- Hoysted, G.A.; Field, K.J.; Sinanaj, B.; Bell, C.A.; Bidartondo, M.I.; Pressel, S. Direct nitrogen, phosphorus and carbon exchanges between Mucoromycotina ‘fine root endophyte’ fungi and a flowering plant in novel monoxenic cultures. New Phytol. 2023, 238, 70–79. [Google Scholar] [CrossRef]
- Koide, R.T.; Kabir, Z. Extraradical hyphae of the mycorrhizal fungus Glomus intraradices can hydrolyse organic phosphate. New Phytol. 2000, 148, 511–517. [Google Scholar] [CrossRef]
- Zou, Y.N.; Xu, Y.J.; Liu, R.C.; Huang, G.M.; Kuča, K.; Srivastava, A.K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Two different strategies of Diversispora spurca-inoculated walnut seedlings to improve leaf P acquisition at low and moderate P levels. Front. Plant Sci. 2023, 14, 1140467. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).