
Morphological, Physiological, and Molecular Stomatal Responses in Local Watermelon Landraces as Drought Tolerance Mechanisms
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Plant Materials
2.2. Planting and Design of Experiment
2.3. Soil Water Content
2.4. Measurement of Physiological Parameters
2.5. Expression Analysis of Stomatal Aperture Genes and Transcription Factors (TFs)
2.6. Statistical Analysis
3. Results
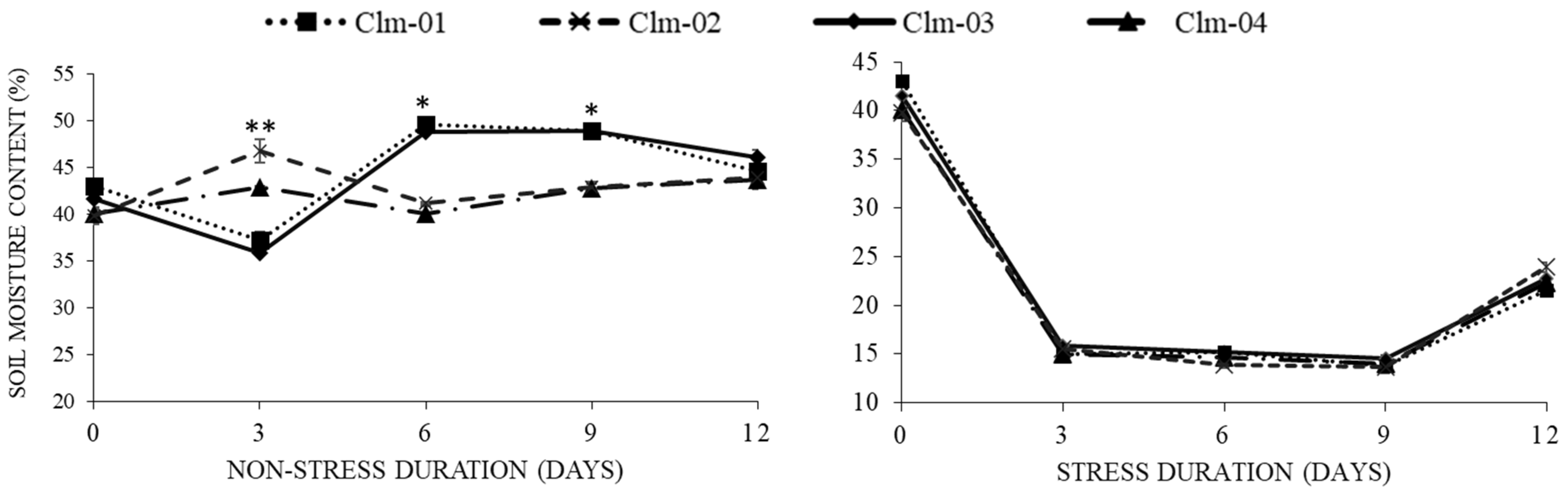
3.1. Soil Water Content
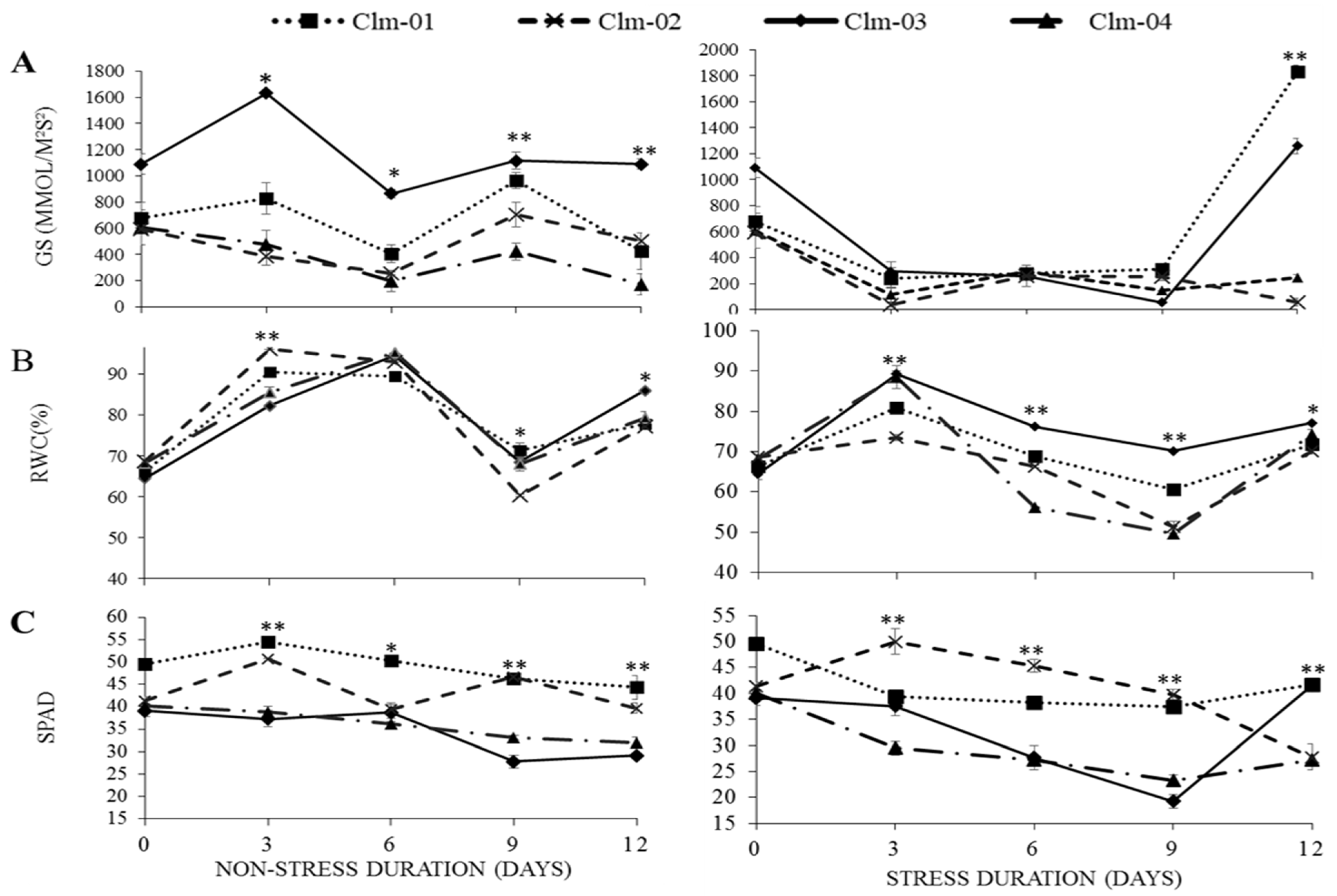
3.2. Physiological Parameters
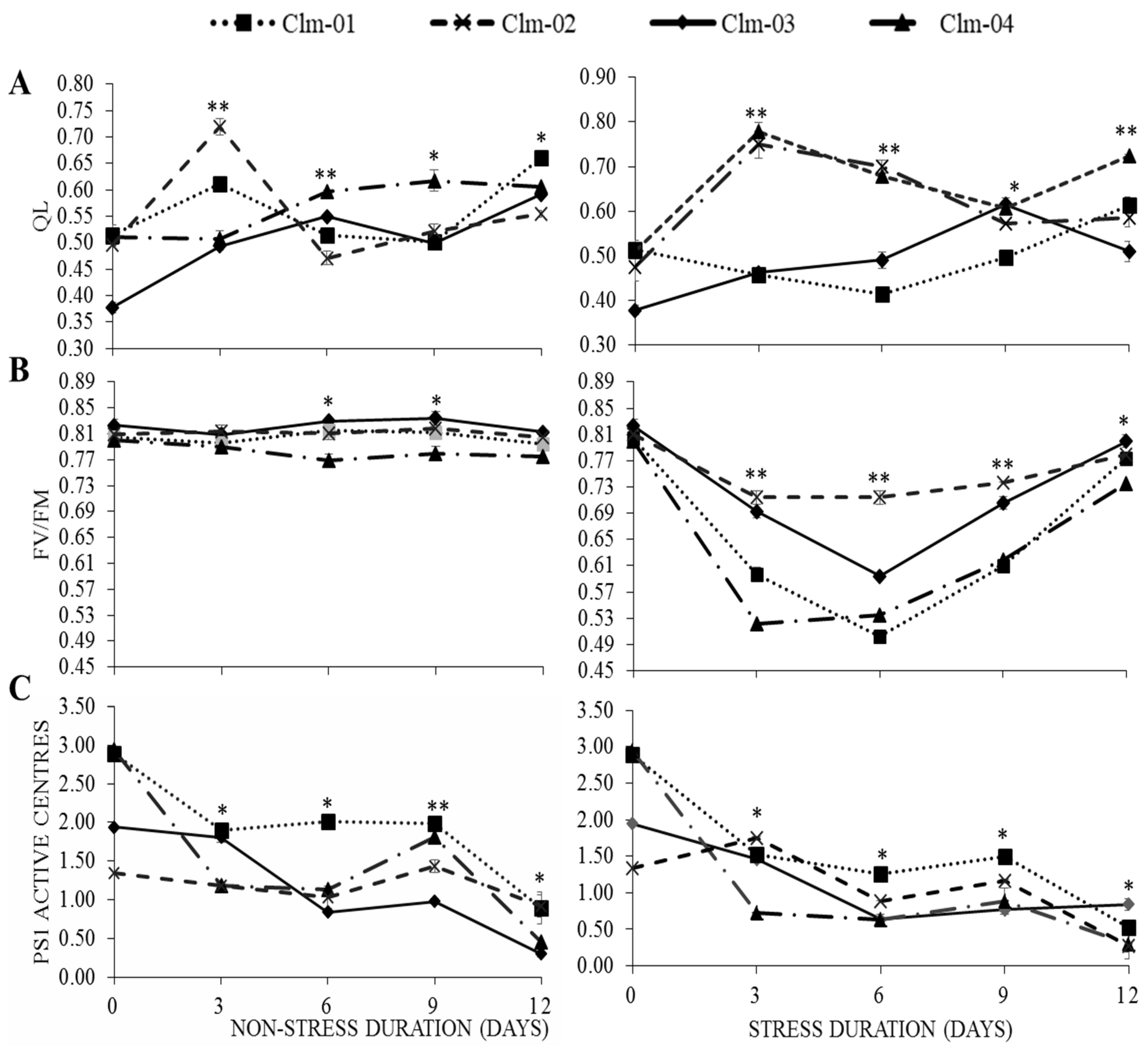
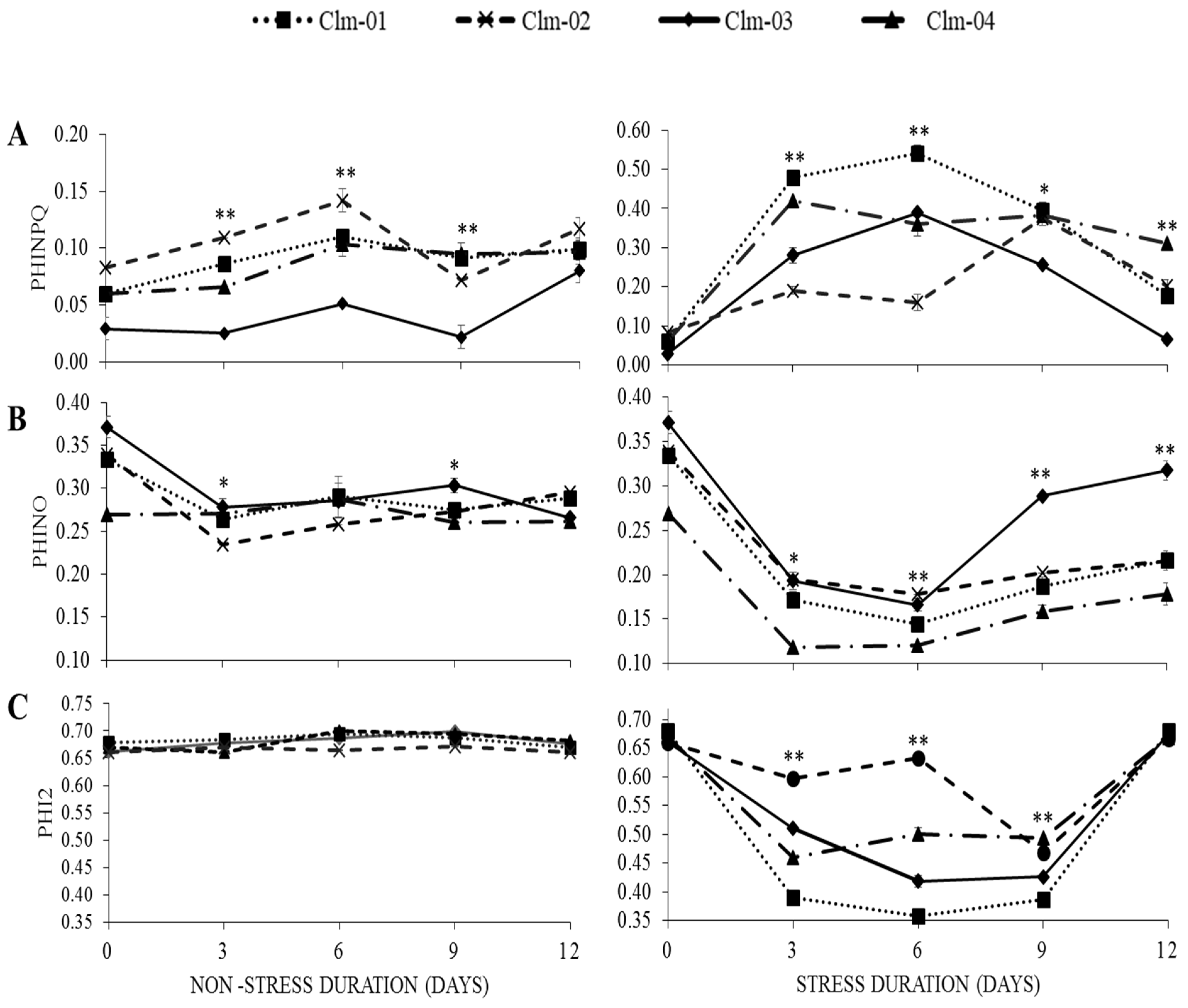
3.3. Chlorophyll Fluorescence (CF) Parameters
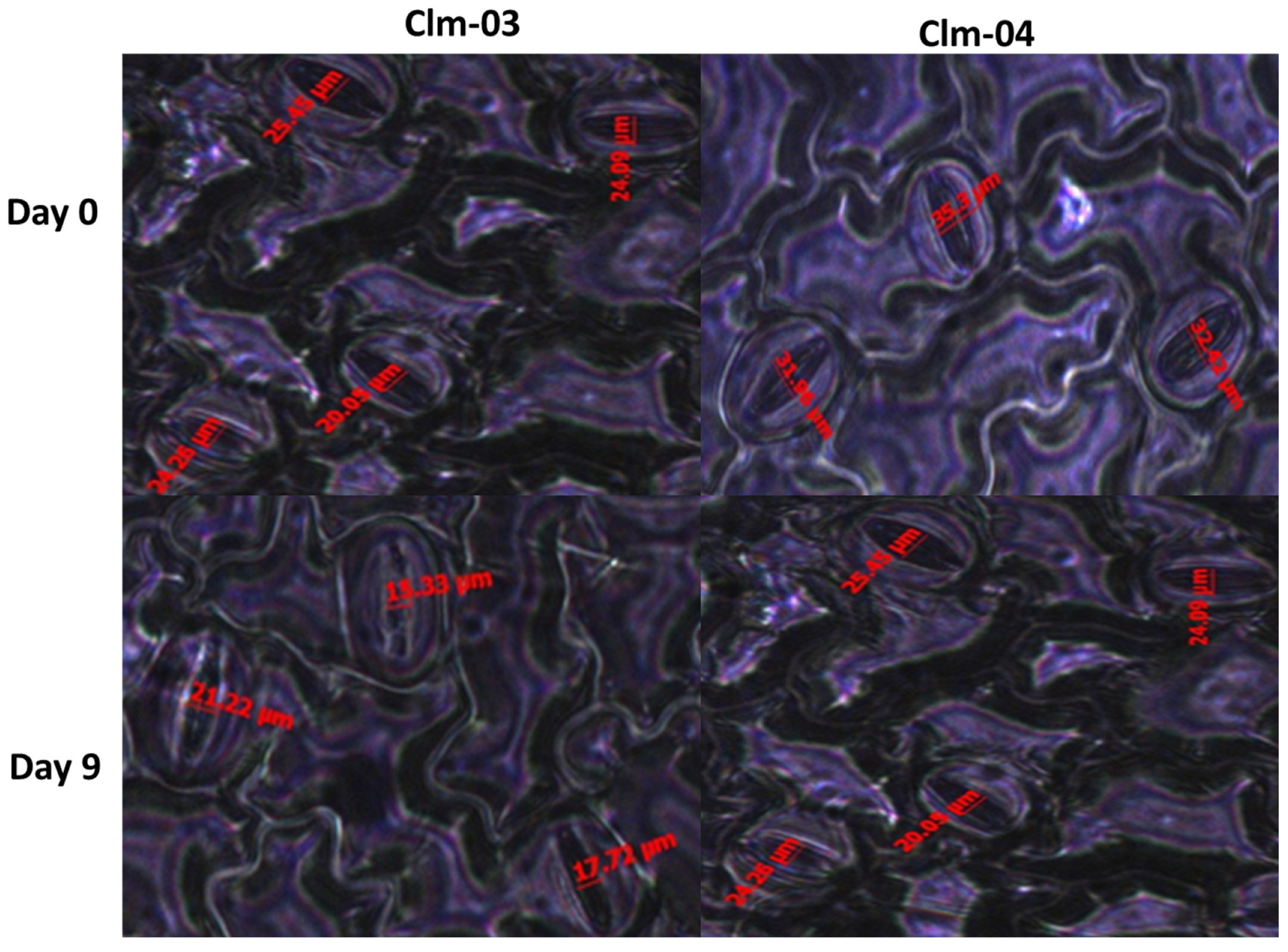
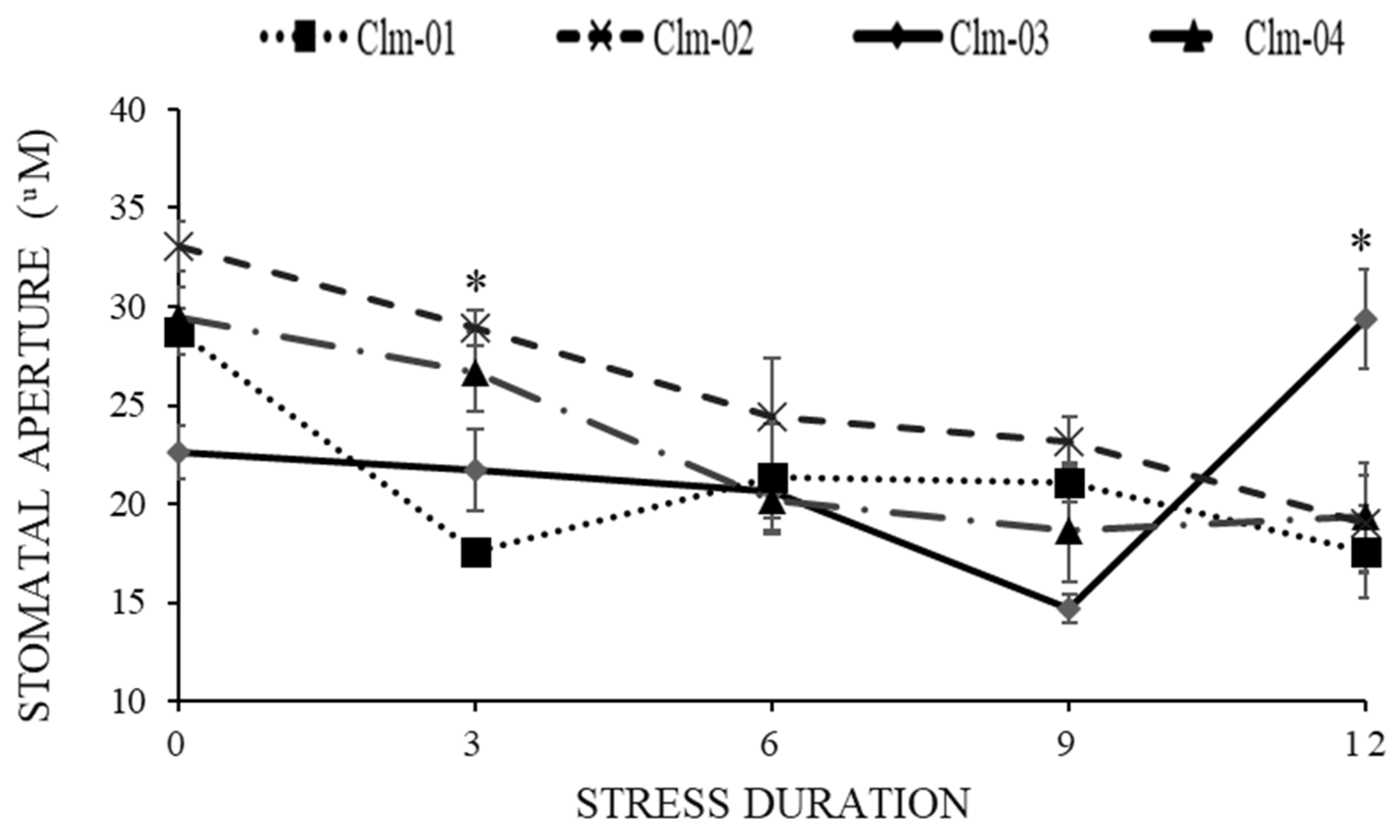
3.4. Stomatal Morphology (Stomatal Aperture Measurements)
3.5. Relative Expression of Plant Transcriptional Regulators with a Function in Stomatal Movement
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Serdeczny, O.; Adams, S. Climate change impacts in Sub-Saharan Africa: From physical changes to their social repercussions. Reg. Environ. Chang. 2017, 17, 1585–1600. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M. Impacts of 1.5 °C global warming on natural and human systems. Glob. Warm. 2018, 175–311. [Google Scholar]
- Ramankutty, N.; Evan, A.T.; Monfreda, C. Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Glob. Biogeochem. Cycles 2002, 16, 1135. [Google Scholar] [CrossRef]
- Sloat, L.L.; Nagy, J.D.; Jones, C.S. Allometric scaling of gas exchange in crops and the implications for carbon balance. Glob. Chang. Biol. 2020, 26, 2814–2828. [Google Scholar]
- Hadi, F.; Hassan, N.; Malik, A. Stomatal regulation for drought tolerance in plants. Int. J. Agric. Biol. 2016, 18, 282–289. [Google Scholar]
- Hajihashemi, S. Stomatal regulation as a drought tolerance mechanism. In Molecular Plant Abiotic Stress: Biology and Biotechnology, 1st ed.; Roychoudhury, A., Tripathi, D.K., Eds.; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2019. [Google Scholar]
- Solis, C.A.; Machado, M.A.; Kozarewa, I. Genetic variation in the wild relatives of cereals: Opportunities for crop improvement. Front. Plant Sci. 2020, 11, 94. [Google Scholar]
- Guoxiong, C.; Yubao, L.; Shiping, W. Wild barley genetic diversity of drought tolerance. Plant Sci. 2002, 163, 837–843. [Google Scholar]
- Cai, S.; Chen, G.; Wang, Y.; Huang, Y.; Marchant, D.B.; Wang, Y.; Yang, Q.; Dai, F.; Hills, A.; Franks, P.J.; et al. Evolutionary Conservation of ABA Signaling for Stomatal Closure. Plant Physiol. 2017, 174, 732–747. [Google Scholar] [CrossRef]
- Chen, Z.H.; Chen, G.; Dai, F.; Wang, Y.Z.; Hills, A.; Ruan, Y.L.; Zhang, G.P.; Franks, P.J.; Nevo, E.; Blatt, M.R. Molecular Evolution of Grass Stomata. Trends Plant Sci. 2017, 22, 124–139. [Google Scholar] [CrossRef]
- Adhikari, K.N.; Buirchell, B.J.; Thomas, G.J.; Sweetingham, M.W.; Yang, H. Identification of anthracnose resistance in Lupinus albus L. and its transfer from landraces to modern cultivars. Crop Pasture Sci. 2009, 60, 472. [Google Scholar] [CrossRef]
- Gupta, C.; Salgotra, R.K.; Mahajan, G. Future Threats and Opportunities Facing Crop Wild Relatives and Landrace Diversity. In Rediscovery of Genetic and Genomic Resources for Future Food Security; Salgotra, R., Zargar, S., Eds.; Springer: Singapore, 2020. [Google Scholar]
- Mayaud, J.R.; Bailey, R.M.; Wiggs, G.F. Modelled responses of the Kalahari Desert to 21st century climate and land use change. Sci. Rep. 2017, 7, 3887. [Google Scholar] [CrossRef] [PubMed]
- Aninbon, C.; Jogloy, S.; Vorasoot, N.; Patanothai, A. Relationship between physiological and root traits of peanut genotypes under terminal drought stress. Int. J. Agric. Technol. 2021, 17, 795–808. [Google Scholar]
- Smart, R.E.; Bingham, G.E. Rapid estimates of relative water content. Plant Physiol. 1974, 53, 258–260. [Google Scholar] [CrossRef] [PubMed]
- Kuhlgert, S.; Austic, G.; Zegarac, R.; Osei-Bonsu, I.; Hoh, D.; Chilvers, M.; Roth, M.; Bi, K.; Teravest, D.; Weebadde, P.; et al. MultispeQ Beta: A tool for large-scale plant phenotyping connected to the open PhotosynQ network. R. Soc. Open Sci. 2016, 3, 160592. [Google Scholar] [CrossRef]
- Pathoumthong, R.; Thammachuan, P.; Seifert, S. Rapid non-destructive method to phenotype stomatal traits. Plant Methods 2023, 19, 36. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Yuan, J.; Gao, L.; Zhao, L.; Cheng, F.; Huang, Y.; Bie, Z. Evaluation of appropriate reference genes for gene expression normalization during watermelon fruit development. PLoS ONE 2015, 10, e0130865. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ajaz, A.; Taghvaeian, S.; Khand, K.; Gowda, P.; Moorhead, J. Development and Evaluation of an Agricultural Drought Index by Harnessing Soil Moisture and Weather Data. Water 2019, 11, 1375. [Google Scholar] [CrossRef]
- Reynolds-Henne, C.E.; Langenegger, A.; Mani, J.; Schenk, N.; Zumsteg, A.; Feller, U. Interactions between temperature, drought, and stomatal opening in legumes. Environ. Exp. Bot. 2010, 68, 37–43. [Google Scholar] [CrossRef]
- Liu, X.; Wei, Z.; Manevski, K.; Liu, J.; Ma, Y.; Andersen, M.N.; Liu, F. Partial root-zone drying irrigation increases water-use efficiency of tobacco plants amended with biochar. Ind. Crops Prod. 2021, 166, 113487. [Google Scholar] [CrossRef]
- Jamaux, I.; Steinmertz, A.; Belhassen, E. Looking for molecular and physiological markers of osmotic adjustment in sunflower. New Phytol. 1997, 137, 117–127. [Google Scholar] [CrossRef]
- Altinkut, A.; Kazan, K.; Ipekci, Z.; Gozukirmizi, N. Tolerance to paraquat is correlated with the traits associated with water stress tolerance in segregating F2 populations of barley and wheat. Euphytica 2001, 121, 81–86. [Google Scholar] [CrossRef]
- Kawasaki, S.M.; Chikahiro, K.; Takayuki, F.; Shinichiro, U.; Masato, Y.A. Responses of Wild Watermelon to Drought Stress: Accumulation of an ArgE Homologue and Citrulline in Leaves during Water Deficits. Plant Cell Physiol. 2000, 41, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Goodarzian, G.M.; Mansurifar, S.; Taghizadeh-Mehrjardi, R.; Saeidi, M.; Jamshidi, A.M.; Ghasemi, E. Effects of drought stress and rewatering on antioxidant systems and relative water content in different growth stages of maize (Zea mays L.) hybrids. Arch. Agron. Soil. Sci. 2015, 61, 493–506. [Google Scholar] [CrossRef]
- Mo, Y.; Wang, Y.; Yang, R.; Zheng, J.; Liu, C.; Li, H.; Ma, J.; Zhang, Y.; Wei, C.; Zhang, X. Regulation of Plant Growth, Photosynthesis, Antioxidation and Osmosis by an Arbuscular Mycorrhizal Fungus in Watermelon Seedlings under Well-Watered and Drought Conditions. Front. Plant Sci. 2016, 7, 644. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Liu, J.; Zhang, J.; Guo, J. Effects of Drought Stress on Photosynthetic and Physiological Parameters of Tomato. J. Amer. Soc. Hort. Sci. 2020, 145, 12–17. [Google Scholar] [CrossRef]
- Nankishore, A.; Farrell, A. The response of contrasting tomato genotypes to combined heat and drought stress. J. Plant Physiol. 2016, 202, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Haworth, M.; Marino, G.; Loreto, F.; Centritto, M. Integrating stomatal physiology and morphology: Evolution of stomatal control and development of future crops. Oecologia 2021, 197, 867–883. [Google Scholar] [CrossRef]
- Zhou, R.; Kjaer, K.; Rosenqvist, E.; Yu, X.; Wu, Z.; Ottosen, C.O. Physiological Response to Heat Stress during Seedling and Anthesis Stage in Tomato Genotypes Differing in Heat Tolerance. J. Agron. Crop Sci. 2017, 203, 68–80. [Google Scholar] [CrossRef]
- Ergo, V.V.; Lascano, R.; Vega, C.R.; Parola, R.; Carrera, C.S. Heat and water stressed field-grown soybean: A multivariate study on the relationship between physiological-biochemical traits and yield. Environ. Exp. Bot. 2017, 148, 1–11. [Google Scholar] [CrossRef]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.C.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. AJCS 2010, 4, 580–585. [Google Scholar]
- Herbinger, K.; Tausz, M.; Wonisch, A.; Soja, G.; Sorger, A.; Grill, D. Complex interactive effects of drought and ozone stress on the antioxidant defence systems of two wheat cultivars. Plant Physiol. Biochem. 2002, 40, 691–696. [Google Scholar] [CrossRef]
- Thomas, D.S.; Turner, D.W. Banana (Musa sp.) leaf gas exchange and chlorophyll fluorescence in response to soil drought, shading, and lamina folding. Sci. Hortic. 2001, 90, 93–108. [Google Scholar]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Li, P.; Wu, Y. Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce. Sci. Hortic. 2012, 135, 45–51. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Malambane, G.; Batlang, U.; Ramolekwa, K.; Tsujimoto, H.; Akashi, K. Growth chamber and field evaluation of physiological factors of two watermelon genotypes. Plant Stress. 2021, 2, 100017. [Google Scholar] [CrossRef]
- Pinnola, A.; Dall’Osto, L.; Gerotto, C.; Morosinotto, T.; Bassi, R.; Alboresi, A. Zeaxanthin binds to light-harvesting complex stress-related protein to enhance nonphotochemical quenching in Physcomitrella patens. Plant Cell 2013, 25, 3519–3534. [Google Scholar] [CrossRef]
- Porcar-Castell, A.; Tyystj€arvi, E.; Atherton, J.; Van Der Tol, C.; Flexas, J.; Pfündel, E.E.; Moreno, J.; Frankenberg, C.; Berry, J.A. Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: Mechanisms and challenges. J. Exp. Bot. 2014, 65, 4065–4095. [Google Scholar] [CrossRef]
- Ismail, I.M.; Basahi, J.M.; Hassan, I.A. Gas exchange and chlorophyll fluorescence of pea (Pisum sativum L.) plants in response to ambient ozone at a rural site in Egypt. Sci. Total Environ. 2014, 497, 585–593. [Google Scholar] [CrossRef]
- Gitau, M.M.; Farkas, A.; Ördög, V.; Maróti., G. Evaluation of the biostimulant effects of two Chlorophyta microalgae on tomato (Solanum lycopersicum). J. Clean. Prod. 2022, 364, 132689. [Google Scholar] [CrossRef]
- Farid, R.; Mutale-Joan, C.; Redouane, B.; Mernissi Najib, E.L.; Abderahime, A.; Laila, S.; Arroussi Hicham, E.L. Effect of Microalgae Polysaccharides on Biochemical and Metabolomics Pathways Related to Plant Defense in Solanum lycopersicum. Appl. Biochem. Biotechnol. 2019, 188, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, W.; Yang, T.; Chen, Y.; Ni, W. Effects of cadmium stress on leaf chlorophyll fluorescence and photosynthesis of Elsholtzia argyi da cadmium accumulating plant. Int. J. Phytoremediation 2015, 17, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Jagtap, V.; Bhargava, S.; Streb, P.; Feierabend, J. Comparative effect of water, heat, and light stresses on photosynthetic reactions in Sorghum bicolor (L.) Moench. J. Exp. Bot. 1998, 49, 1715–1721. [Google Scholar]
- Jeon, M.W.; Ali, M.B.; Hahn, E.J.; Paek, K.Y. Photosynthetic pigments, morphology, and leaf gas exchange during ex vitro acclimatization of micropropagated CAM doritaenopsis plantlets under relative humidity and air temperature. Environ. Exp. Bot. 2006, 55, 183–194. [Google Scholar] [CrossRef]
- Mandizvo, T.; Odindo, A.O.; Mashilo, J.; Magwaza, L.S. Drought tolerance assessment of citron watermelon (Citrullus lanatus var. citroides (L.H. Bailey) Mansf. ex Greb.) accessions based on morphological and physiological traits. Plant Physiol. Biochem. 2022, 180, 106–123. [Google Scholar] [CrossRef]
- Bashir, N.; Athar, H.; Hazem, M.; Kalaji, J.W.; Mahmood, S.; Zafar, Z.U.; Ashraf, M. Is Photoprotection of PSII One of the Key Mechanisms for Drought Tolerance in Maize? Int. J. Mol. Sci. 2021, 22, 13490. [Google Scholar] [CrossRef]
- Ruban, A.V.; Wilson, S. The Mechanism of Non-Photochemical Quenching in Plants: Localization and Driving Forces. Plant Cell Physiol. 2021, 62, 1063–1072. [Google Scholar] [CrossRef]
- Bosabalidis, A.M.; Kofidis, G. Comparative effects of drought stress on leaf anatomy of two olive cultivars. Plant Sci. 2002, 163, 375–379. [Google Scholar] [CrossRef]
- Xu, Z.Z.; Zhou, G.S. Effects of water stress and high nocturnal temperature on photosynthesis and nitrogen level of a perennial grass Leymus chinensis. Plant Soil. 2005, 269, 131–139. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Malambane, G.; Madumane, K.; Sewelo, L.T.; Batlang, U. Drought stress tolerance mechanisms and their potential common indicators to salinity, insights from the wild watermelon (Citrullus lanatus): A review. Front. Plant Sci. 2023, 13, 1074395. [Google Scholar] [CrossRef] [PubMed]
- Bertolino, L.T.; Caine, R.S.; Gray, J.E. Impact of stomatal density and morphology on water use efficiency in a changing world. Front. Plant Sci. 2019, 10, 225. [Google Scholar] [CrossRef] [PubMed]
- Century, K.; Lynne, R.; Oliver, J.R. Regulating the Regulators: The Future Prospects for Transcription-Factor-Based Agricultural Biotechnology Products. Plant Physiol. 2008, 147, 20–29. [Google Scholar] [CrossRef]
- Finkelstein, R.; Gampala, S.; Rock, C. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002, 14, 15–45. [Google Scholar] [CrossRef]
- Leung, J.; Giraudat, J. Abscisic acid signal transduction. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 199–222. [Google Scholar] [CrossRef]
- Nilson, S.E.; Assman, S.M. The Control of transpiration. Insights from Arabidopsis. Plant Physiol. 2007, 143, 19–27. [Google Scholar] [CrossRef]
- Schroeder, J.I.; Kwak, J.M.; Allen, G.J. Guard cell abscisic acid signaling and engineering drought hardiness in plants. Nature 2001, 410, 327–330. [Google Scholar] [CrossRef]
- Liu, H.; Shen, J.; Yuan, C.; Lu, D.; Acharya, B.; Wang, M.; Chen, D.; Zhang, W. The Cyclophilin ROC3 Regulates ABA-Induced Stomatal Closure and the Drought Stress Response of Arabidopsis thaliana. Front. Plant Sci. 2021, 12, 668792. [Google Scholar] [CrossRef]
- Nanasato, Y.; Miyake, C.; Takahara, K.; Kohzuma, K.; Munekage, Y.N.; Yokota, A.; Akashi, K. Chapter 23 Mechanisms of Drought and High Light Stress Tolerance Studied in a Xerophyte, Citrullus lanatus (Wild Watermelon). In Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2010; pp. 363–378. [Google Scholar] [CrossRef]
- Eleonora, C.; Massimo, G.; Chiara, T. Transcription: Transcription factors controlling stomatal movements and drought tolerance. Transcription 2010, 1, 41–45. [Google Scholar] [CrossRef]
- Matthews, J.S.A.; Vialet-Chabrand, S.; Lawson, T. Role of blue and red light in stomatal dynamic behavior. J. Exp. Bot. 2020, 71, 2253–2269. [Google Scholar] [CrossRef] [PubMed]
- Daszkowska-Golec, A.; Szarejko, I. Open or close the gate—Stomata action under the control of phytohormones in drought stress conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar] [CrossRef]
- Simeoni, F.; Skirycz, A.; Simoni, L.; Castorina, G.; de Souza, L.P.; Fernie, A.R.; Alseekh, S.; Giavalisco, P.; Conti, L.; Tonelli, C.; et al. The AtMYB60 transcription factor regulates stomatal opening by modulating oxylipin synthesis in guard cells. Sci. Rep. 2022, 12, 533. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qin, L.; Han, L.; Xiang, Y.; Zhao, D. Overexpression of maize SDD1 (ZmSDD1) improves drought resistance in Zea mays L. by reducing stomatal density. Plant Cell Tissue Organ Cult. 2015, 122, 147–159. [Google Scholar] [CrossRef]
- Hughes, J.; Hepworth, C.; Dutton, C.; Dunn, J.A.; Hunt, L.; Stephens, J.; Waugh, R.; Cameron, D.D.; Gray, J.E. Reducing stomatal density in barley improves drought tolerance without impacting on yield. Plant Physiol. 2017, 174, 776–787. [Google Scholar] [CrossRef]
- Caine, R.S.; Yin, X.; Sloan, J.; Harrison, E.L.; Mohammed, U.; Fulton, T.; Biswal, A.K.; Dionora, J.; Chater, C.C.; Coe, R.A.; et al. Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. New Phytol. 2019, 221, 371–384. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Accession Type | Cultivation | Source | Type of Use | Flesh Color |
|---|---|---|---|---|---|
| Clm-01 | Landrace | Domesticated | BNPGRC | Fresh eating | Red |
| Clm-02 | Landrace | Domesticated | BNPGRC | Cooking | Yellow |
| Clm-03 | Landrace | Wild | BNPGRC | Livestock feed | White |
| Clm-04 | Hybrid | Domesticated | Commercial shop | Fresh eating | Red |
| Name | Pathways | Role | Target TFs of Stomatal Aperture and Reference Genes in Wild Watermelon |
|---|---|---|---|
| Cla017012 | dark/light, ABA | opening | F_CCAATACTGGGTTGCTTAGATGTAG R_GTTCTTTGTGGAAGGTATGAAGCTA |
| Cla017389 | ABA | opening | F_TCTCTACACTGTTCCTGAAAATTCC R_TGTTCTGACCAAAACCTAATCTCTC |
| Cla016849 | ABA | closure | F_ACATCTTCATGCACTAAACAGAGTG R:GATCCATTAGAGATGCTTGTGATCT |
| Cla004380 | ABA | closure | F_GTGGAAGAAGCTCTACAAAGTCAAC R_AAGAGTGTCTTCTTCCTGGTTGTAA |
| Glyceraldehyde 3-phosphate dehydrogenase(GAPDH) | F_CTGGCAGTACTTTGCCAACA R_AGGATTGGAGAGGAGGTCGT | ||
| Tubulin | F_CAGCACTCCTAGCTTTGGTGA R_CGGGGAAATGGGATTAGATT | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madumane, K.; Sewelo, L.T.; Nkane, M.N.; Batlang, U.; Malambane, G. Morphological, Physiological, and Molecular Stomatal Responses in Local Watermelon Landraces as Drought Tolerance Mechanisms. Horticulturae 2024, 10, 123. https://doi.org/10.3390/horticulturae10020123
Madumane K, Sewelo LT, Nkane MN, Batlang U, Malambane G. Morphological, Physiological, and Molecular Stomatal Responses in Local Watermelon Landraces as Drought Tolerance Mechanisms. Horticulturae. 2024; 10(2):123. https://doi.org/10.3390/horticulturae10020123
Chicago/Turabian StyleMadumane, Kelebogile, Lesego T. Sewelo, Metseyabeng N. Nkane, Utlwang Batlang, and Goitseone Malambane. 2024. "Morphological, Physiological, and Molecular Stomatal Responses in Local Watermelon Landraces as Drought Tolerance Mechanisms" Horticulturae 10, no. 2: 123. https://doi.org/10.3390/horticulturae10020123
APA StyleMadumane, K., Sewelo, L. T., Nkane, M. N., Batlang, U., & Malambane, G. (2024). Morphological, Physiological, and Molecular Stomatal Responses in Local Watermelon Landraces as Drought Tolerance Mechanisms. Horticulturae, 10(2), 123. https://doi.org/10.3390/horticulturae10020123